基于iTRAQ 的玉米V12 时期响应干旱胁迫的 蛋白质组学分析
2021-01-08刘松涛董安忆杨亚桐段会军
刘 过,刘松涛,董安忆,杨亚桐,段会军
(河北农业大学 农学院/华北作物种质资源教育部重点实验室,河北 保定 071000)
玉米是重要的粮食作物之一,在农业生产占据重要地位。干旱是制约玉米高品质丰产的主要原因,在不同程度上限制了植物的生长发育,导致植物的分子、细胞、器官和个体水平发生不利变化,如水平衡在植物中受到破坏,导致老叶枯黄,新叶萎蔫生长缓慢,活性氧清除系统紊乱,激素分泌紊乱 等[1]。同时,植物对干旱协迫做出有利的应激反应,首先通过传递干旱胁迫信号来调控抗旱基因的表达,然后合成一系列调控植物生长的蛋白质。这些胁迫调节反应反映在形态变化、气孔调节、渗透调节、活性氧清除等方面,但干旱胁迫的响应机制往往需要多种机制的协同作用[2]。玉米在喇叭口期生长发育最旺盛,对肥水条件最敏感,这一阶段水分的缺乏会导致雄穗发育不良,雌穗分化受阻,导致每穗粒数严重减少,最终影响后期产量[3]。因此,研究V12(玉米12 叶龄)时期玉米响应干旱胁迫差异表达蛋白,对揭示玉米V12 时期抵御干旱的调控机理有重要意义。
2001 年,真正意义上有关玉米的蛋白质组学研究正式开始[4],包括玉米不同器官/组织的蛋白质组学分析,玉米不同器官间相互关系的蛋白质组学分析,玉米细胞器蛋白质组学分析,玉米胁迫相关蛋白质组学分析。近20 年来,国内外玉米蛋白质组学进行了大量研究,主要集中在玉米不同组织及生物或非生物胁迫。本研究采用iTRAQ 技术,以耐旱性不同的玉米杂交种为材料,比较干旱胁迫下V12 时期玉米叶片蛋白质水平上的差异,分析与响应干旱胁迫相关的差异蛋白的特性及生物学功能,初步揭示玉米在V12 时期抵御干旱胁迫的分子机理,为玉米抗旱育种及品种改良进一步研究提供参考。
1 材料与方法
1.1 试验材料与干旱处理
试验在河北农业大学清苑实验基地抗旱棚内进行,以玉米杂交种‘农单476’和‘众信978’为试验材料,于2018 年6 月15 日播种于河北保定清苑抗旱棚。杂交种‘农单476’是由河北农业大学玉米改良中心育成的国审品种,‘众信978’是由河北众信种子科技有限公司选育省审品种。由河北农林科学院旱作所鉴定‘农单476’为抗旱品种,‘众信978’为敏感品种。小区面积25 m2,行间距0.6 m, 株距0.3 m,双粒穴播,水旱区1 m 防水墙间隔。各小区底肥均为复合肥512 kg/hm2。当75%的玉米植株长到8 叶时(7 月18 日)开始干旱处理,采用TZS-1 土壤水分测定仪(浙江拓普科技有限公司)对地下1 m 土壤相对含水量进行监测,每20 cm 测量1 次。对照组控制田间土壤含水量20%~30%,处理组控制田间土壤含水量5%~10%。到12 叶时(7月30 日)取长势相近的玉米的旗叶叶尖部分五厘米左右液氮速冻,3 个生物学重复,然后储存在-80 ℃ 超低温冰箱用于进行进一步分析。
1.2 总蛋白提取
利用冷丙酮法[5-6]从两个玉米杂交种的试验组和对照组的叶片组织中提取总蛋白,每个样品设置3 次生物学重复。
1.3 蛋白酶解与iTRAQ 标记
分别取蛋白样品100 μg,加入蛋白溶液至100 μL, 加入2 μL 1μg/μL 胰酶和500 μL 100 mmol/L TEAB 缓冲液,混匀,37 ℃下酶切过夜。加入相同体积的1%甲酸,混匀,室温12 000 r/min 离心5 min。取上清,缓慢通过C18 脱盐塔,用1 mL 清洗液(0.1%甲酸,4%乙腈)连续3 次。加入0.4 mL 洗脱液(0.1%甲酸,45%乙腈)连续2 次。洗脱液样品混合后,冷冻干燥。加入20 μL 0.5 mmol/L TEAB 缓冲液再次溶解,加入足够的iTRAQ 标记试剂溶于异丙醇中,室温下倒置搅拌1 h。加入100 μL 50 mmol/L Tris-HCl(pH=8)停止反应,等体积标记后取样并混合、脱盐和冷冻干燥。多个标记组时取所有样品进行等质量蛋白混合作为标记组间的共同参照样品。
1.4 SCX 分级与LC-MS/MS
采用L-3000 高效液相色谱系统进行强阳离子交换(SCX)色谱分离iTRAQ 标记的肽混合物。详细步骤参考本实验室已发表论文[7]。采用Q 型精密TM HF-X 质谱仪,易构建TM 离子源,设置离子喷射电压为2.3 kV,离子传输管温度320 ℃,质谱仪采用数据相关采集方式,质谱扫描范围为m/Z 350 ~1 500,一级质谱仪分辨率为60 000(200 m/z), C-trap 的最大容量为3×106,C-trap 的最大注入时间为20 ms,用高能碰撞劈裂HCD 法选择离子强度在全扫描范围内前40 位的母离子进行裂解,并用二次质谱法进行检测。二次质谱的分辨率设定为15 000 (200 m/z),C-trap 的最大容量为1×105,C-trap 的最大注入时间为45 ms,肽片段的碰撞能量设定为32%,阈值强度设定为8.3×103,排除的动态范围设定为60 s,产生质谱检测原始数据。
1.5 数据质量控制
在蛋白质数据库中搜索质谱数据。数据库的选择是生物信息后续分析的关键步骤。这次使用的数据 库 是:P101SC18122835-01-Zea_mays-ensemblrelea.fasta:131585 个序列。质谱仪的数据格式为*raw,存储了质谱仪数据的完整扫描信息。原始文件将直接导入Proteome Discoverer2.2 软件,用于数据库检索、肽和蛋白质定量。
1.6 蛋白质功能注释
GO 注释是利用interproscan 软件对鉴定到的蛋白质进行注释分析,涉及6 个数据库的搜索。KEGG与COG 注释首先将鉴定到的蛋白质进行BLAST 比对(blastp,E-value ≤1e-4),然后对于每一条序列的BLAST 结果选取score 得分最高的进行注释。
1.7 差异蛋白筛选
确定需要比较的样品对:2 个品种、2 个处理确定4 个比较分组,分别为ZXD_ZXC、ZND_ZNC、ZND_ZXD、ZNC_ZXC。取比较样品对中各蛋白质生物重复性所有定量值的平均值之比作为差异倍数FC。采用t检验对各蛋白的相对定量值进行检验,以p值作为判断差异是否显著的指标。当FC ≥1.2 且P-vaule ≤0.05 表示蛋白质表达量上调,FC ≤0.83 且P-vaule ≤0.05 表示蛋白质表达量下调[8]。
1.8 差异表达蛋白富集分析
GO 功能显著性富集分析首先把所有差异蛋白质向Gene Ontology 数据库(http://www.geneontology.org/)的各个term 映射,计算每个term 的蛋白质数目,然后应用超几何检验,找出与所有蛋白质背景相比,在差异蛋白质中显著富集的GO term。计算得到P-value 值,以P-value ≤0.05 为阈值,满足此条件的GO term 定义为在差异蛋白质中显著富集的GO term。KEGG Pathway 显著性富集分析方法同 GO 功能富集分析,是以 KEGG Pathway 为单位,应用超几何检验,以P-value ≤0.05 为阈值,找出与所有鉴定到蛋白背景相比,在差异蛋白中显著性富集的 Pathway。
2 结果与分析
2.1 iTRAQ 数据鉴定及质控分析结果
利用Proteome Discoverer2.2 软件对检索结果进一步过滤:可信度在95%以上的肽匹配图谱(Peptide Spectrum Matches,简称PSMs)为可信PSMs,至少包含一个unique 肽段(特有肽段,简称肽段)的蛋白为可信蛋白,只保留可信的肽匹配图谱和蛋白,并做FDR 验证,剔除FDR 大于5%的肽段和蛋白。下表是鉴定到的肽段数和蛋白数总体情况。只利用表中展示的过滤后的结果来做后续分析。
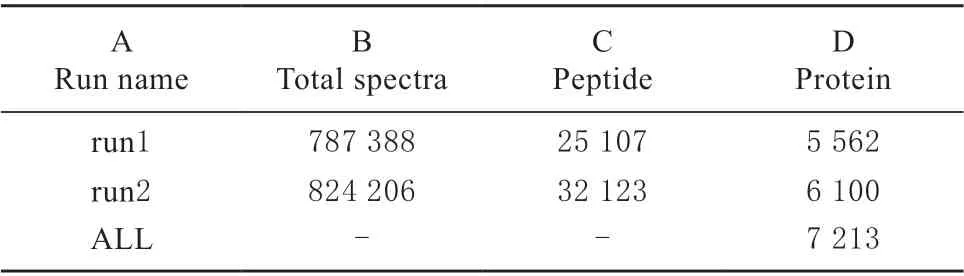
表1 蛋白质鉴定信息Table 1 Information of protein identification
质谱下机的数据,在搜库完成后,需要进行一系列质控,包括肽段长度分布(图1A)、蛋白分子量分布(图1B)、蛋白覆盖度分布(图1C)、肽段数分布(图1D)。每种质谱仪都有自身的测量范围,本次鉴定到肽段的长度分布如图1A 所示,肽段长度90%以上在6 ~22,峰值在10 ~12,肽段长度及分布合理。
2.2 蛋白功能注释
本研究将鉴定到的蛋白质分别注释到GO、KEGG、COG 和IPR 数据库,以了解不同蛋白质的功能特性,数据库注释结果统计见图2。这4个数据库共同注释到的蛋白质有3 468 个,GO 数据库没有单独注释到蛋白,COG 数据库单独注释到1 个蛋白,KEGG 数据库单独注释到490 个蛋白,IPR 数据库单独注释到6 个蛋白。
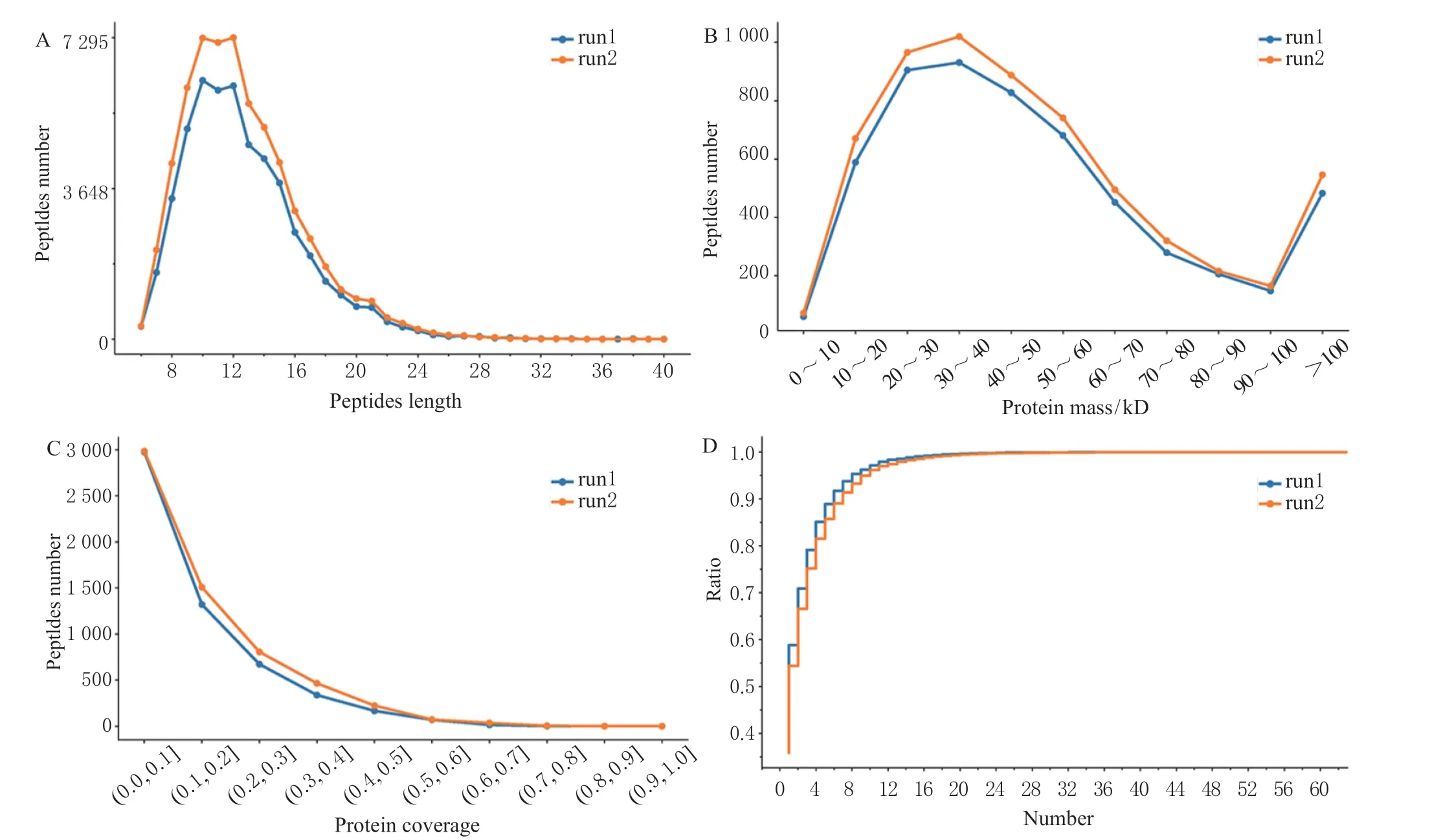
图1 蛋白数据质控Fig. 1 Protein data quality control
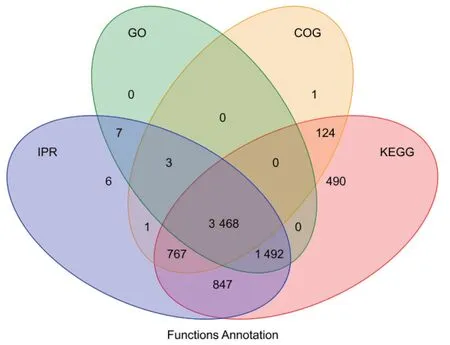
图2 功能注释结果Fig. 2 Function annotation result
2.3 玉米V12 时期响应干旱胁迫差异表达蛋白
通过iTRAQ 技术标记差异比较组样本蛋白,经过质谱鉴定和定量获得不同表达量蛋白,分析后得到差异蛋白。本研究以2 种具有不同耐旱性的玉米杂交种为材料,在干旱胁迫和正常浇水条件下,‘农单476’差异表达蛋白49 个,包括上调29 个,下调20 个(表2、表3),‘众信978’差异表达蛋白64 个,包括上调46 个,下调18 个(表2、 表4),表明干旱胁迫下耐旱性不同的玉米杂交种叶片蛋白表达量均发生不同程度变化以抵御干旱保持植株正常生长,耐旱型‘农单476’差异表达蛋白数目低于敏感型‘众信978’差异表达蛋白数目,表明耐旱型玉米品种在干旱胁迫下只需要较少蛋白互作变化就能抵御干旱,正常生长,适应性更强。
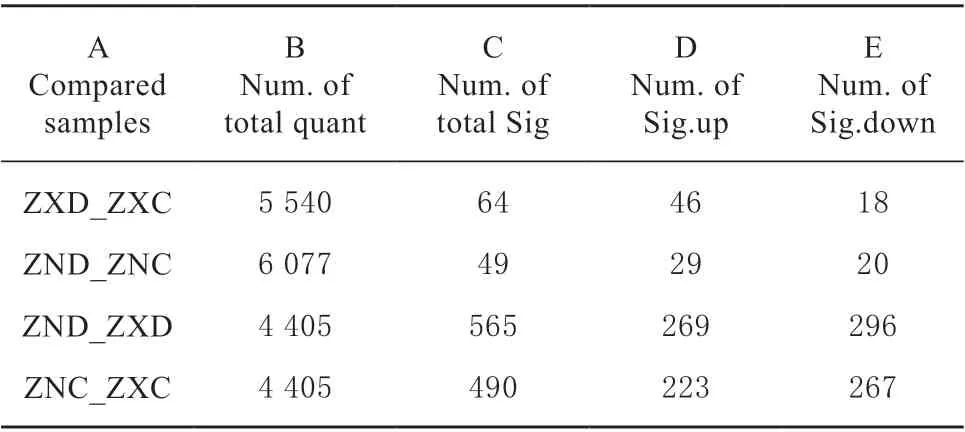
表2 蛋白差异结果统计Table 2 Statistical results for DAPs
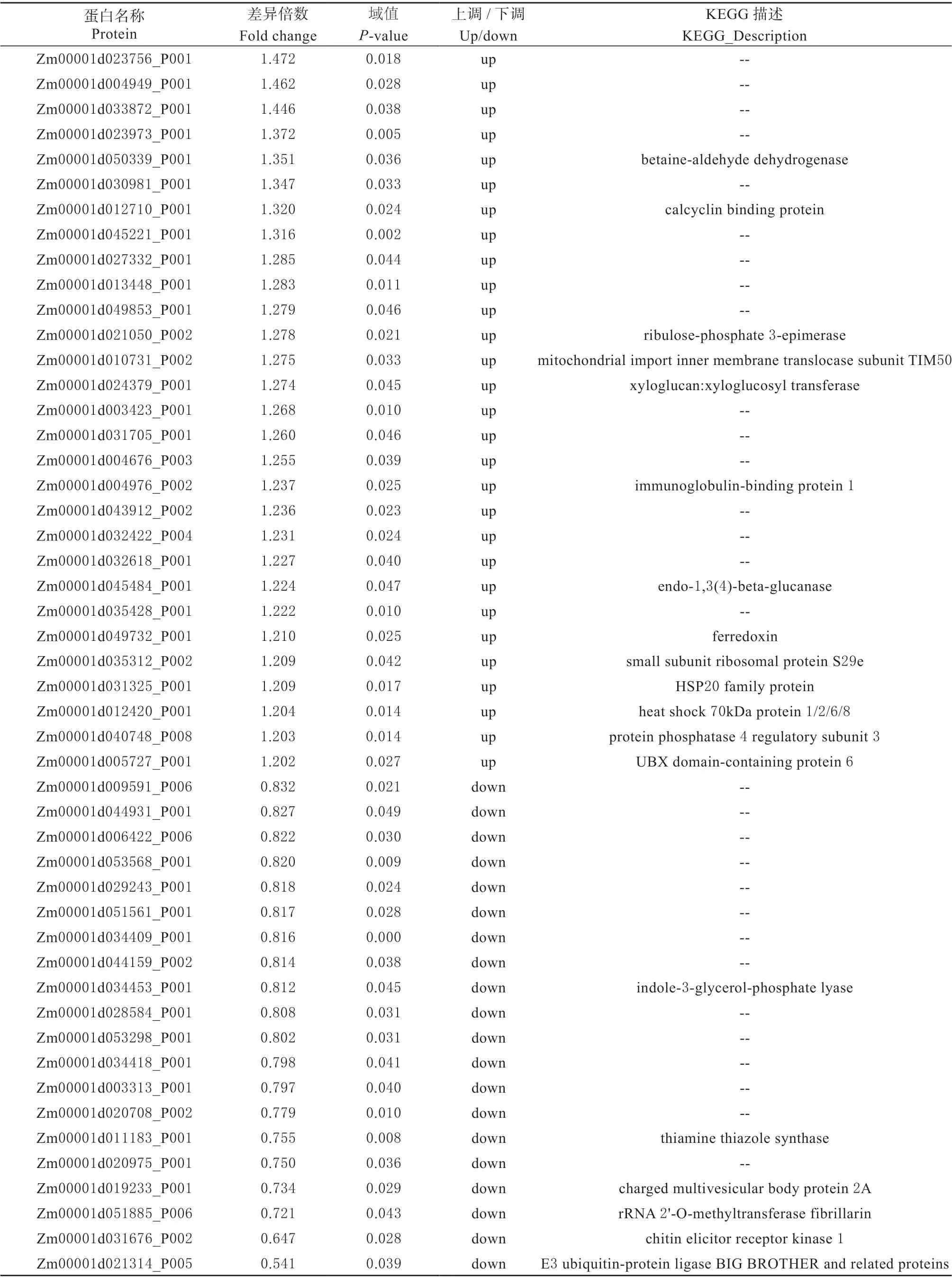
表3 ‘农单476’在干旱胁迫下的差异表达蛋白Table 3 The DAPs of‘ND476’under drought stress
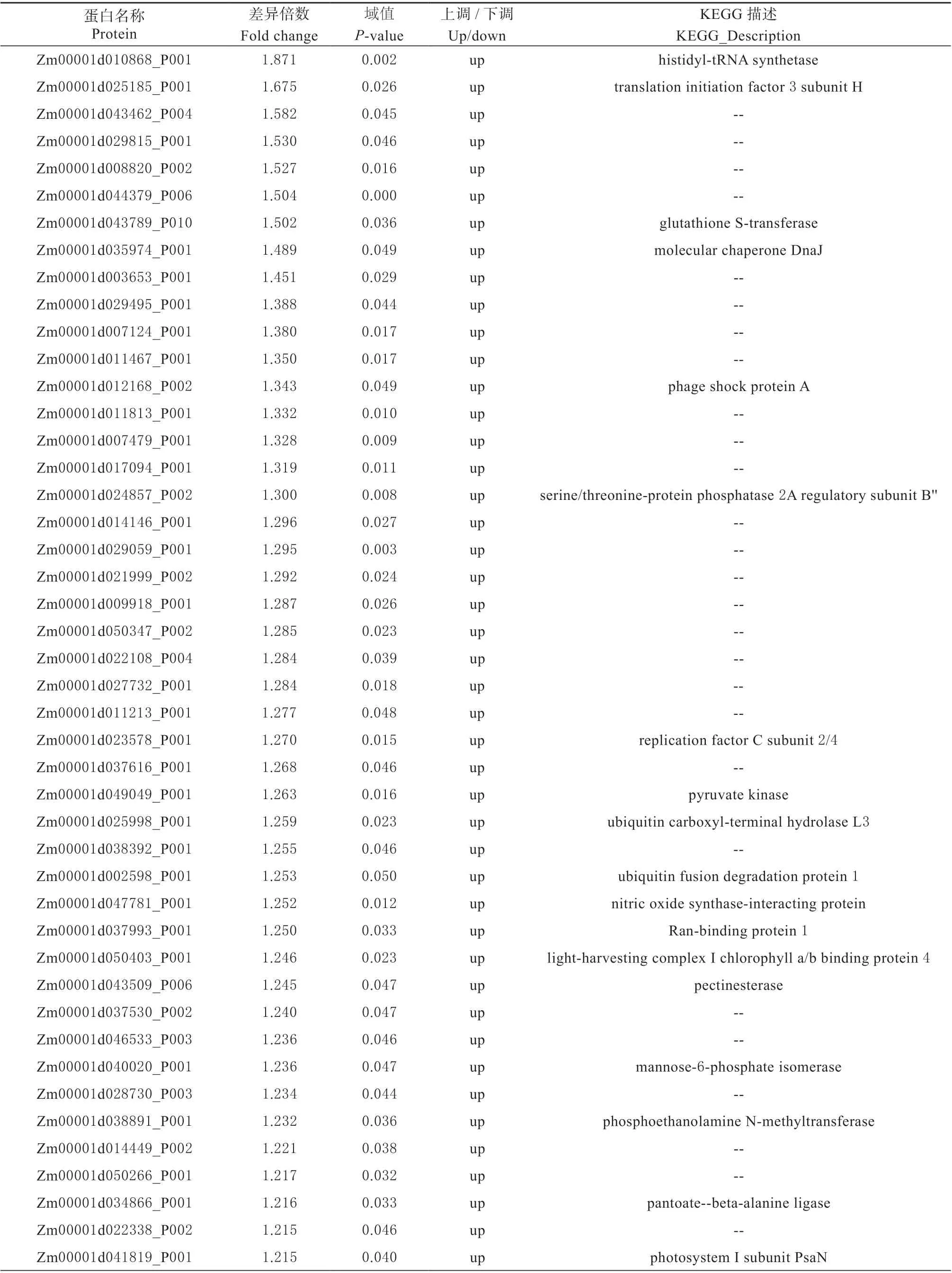
表4 ‘众信978’在干旱胁迫下的差异表达蛋白Table 4 The DAPs of‘ZX978’under drought stress
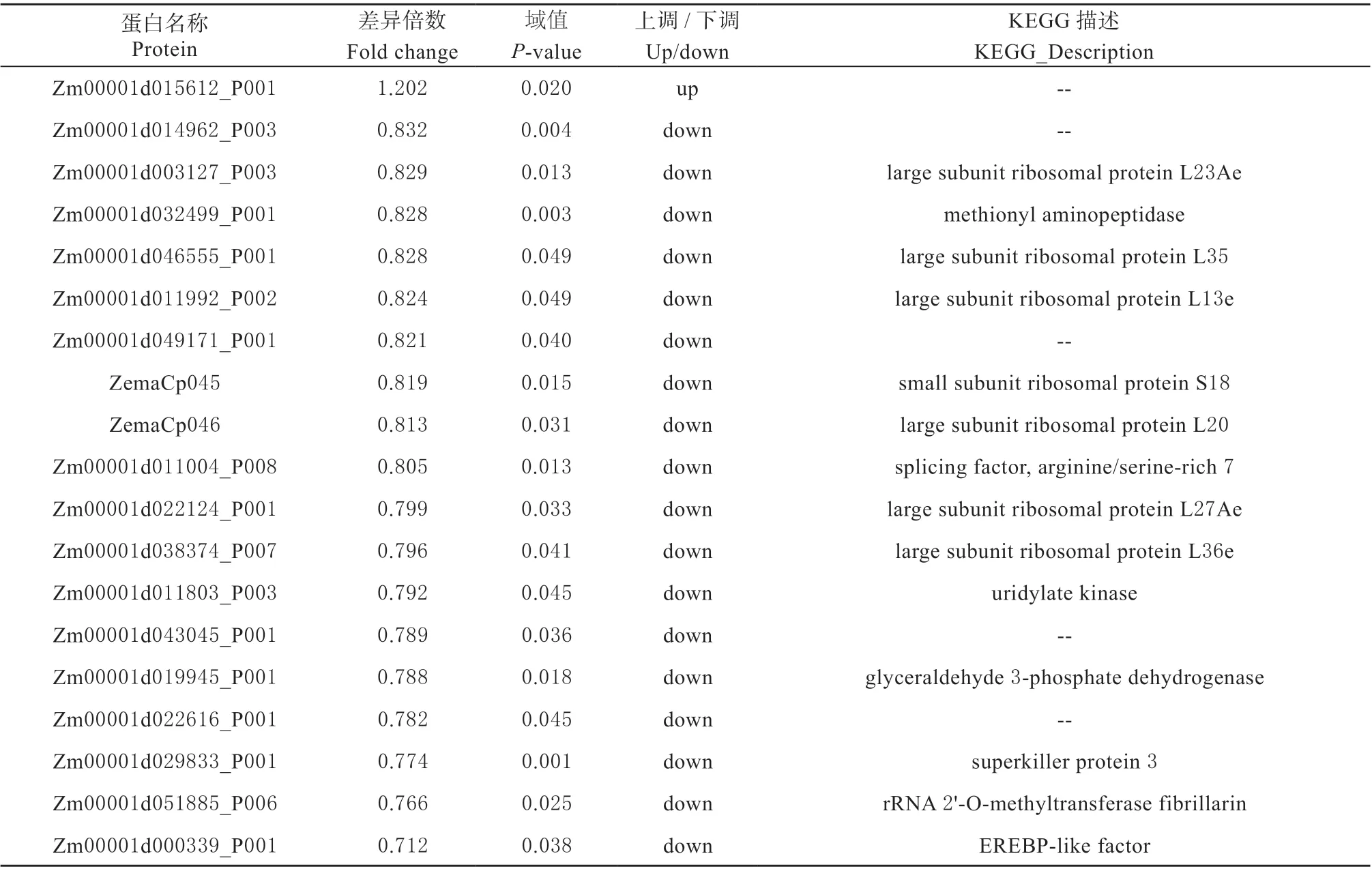
续表:
从图3A 中可以看出干旱胁迫下的‘农单476’的差异表达蛋白,上半部分(29 个DAPs)表达水平:ZND >ZNC,对于这部分差异蛋白来说,表达量逐渐升高,因此这部分差异蛋白表现为上调;下半部分(20 个DAPs)差异蛋白表现则相反。从图3B 中可以看出干旱胁迫下‘众信978’的差异表达蛋白,上半部分(18 个DAPs)表达水平:ZXD<ZXC,对于这部分差异蛋白来说,表达量逐渐降低,因此这部分差异蛋白表现为下调;下半部分(46个DAPs)差异蛋白表现则相反。
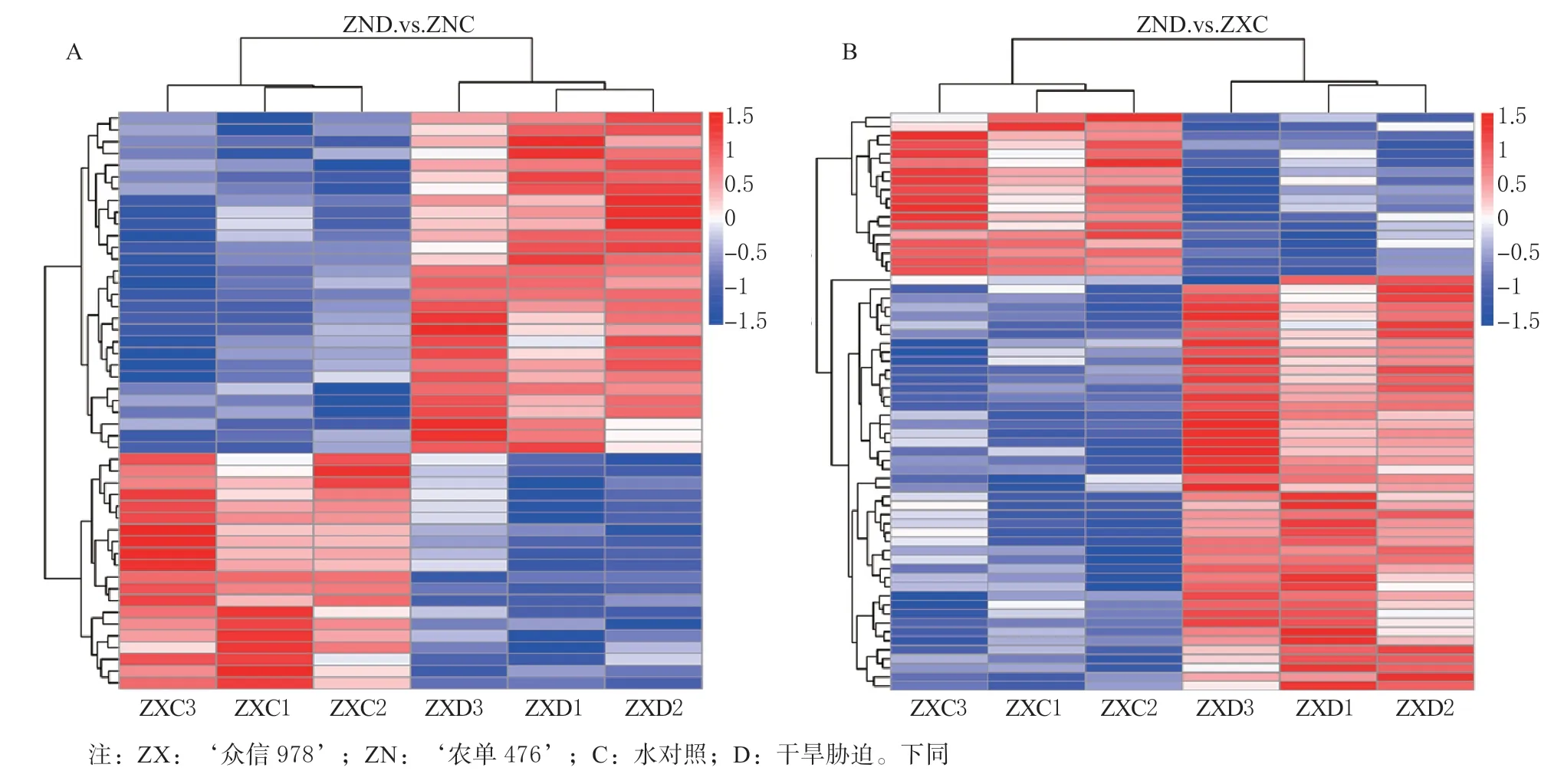
图3 干旱前后的DAPs 在所有个体中的聚类图Fig. 3 Cluster of DAPs before and after drought stress in all individuals
2.4 ZND_ZNC 比较组中差异蛋白功能富集分析
ZND_ZNC 比较组中得到差异蛋白49 个,ZND组与ZNC 组分别代表的是‘农单476’杂交种干旱处理后和水对照,通过这2 个组的比较,得到玉米耐旱品种中响应干旱的关键差异蛋白。对该比较组差异蛋白分别进行GO 富集和KEGG pathway 富集分析。GO富集分析发现差异蛋白主要富集到4 个分子功能的GO terms 见图4,分别为:过渡金属离子结合、半胱氨酸类肽酶活性、铁离子结合和电子载体活性。对差异表达蛋白的KEGG pathway 富集分析见图5,这些差异蛋白在内质网蛋白质加工和苯并恶唑嗪酮类化合物(BXs)的生物合成信号通路中最显著富集。
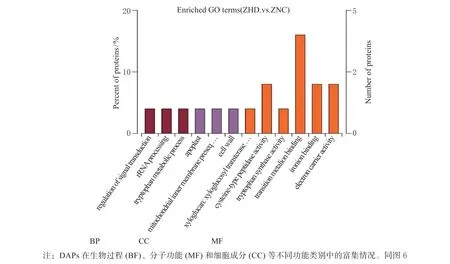
图4 ZND_ZNC 比较组差异蛋白的GO 富集分析Fig. 4 GO enrichment analysis of ZND_ZNC comparison DAPs
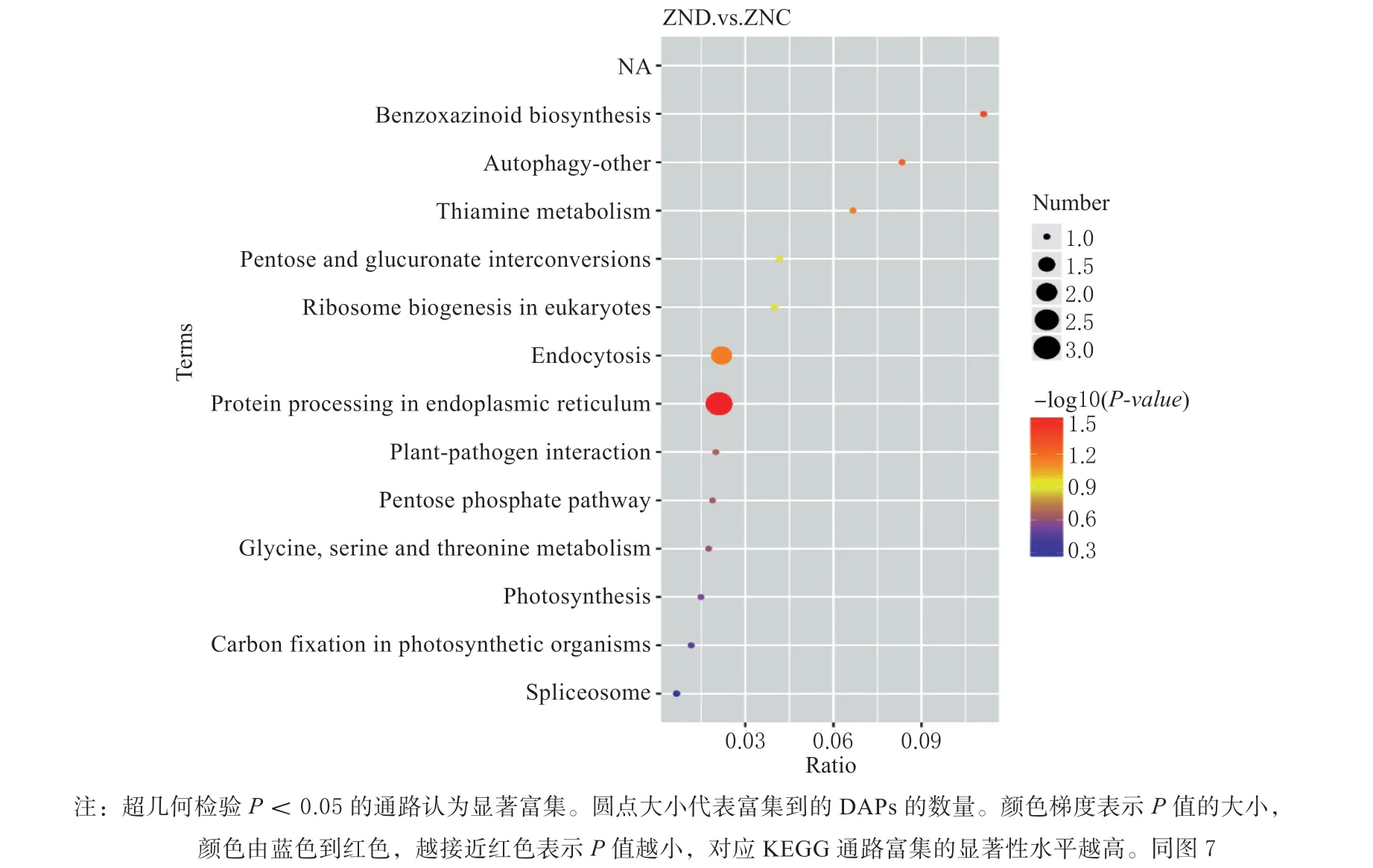
图5 ZND_ZNC 比较组差异蛋白的KEGG 途径富集分析Fig. 5 KEGG pathway enrichment analysis of ZND_ZNC comparison DAPs
2.5 ZXD_ZXC 比较组差异蛋白功能富集分析
ZXD_ZXC 比较组中有差异蛋白64 个,ZXD 组与ZXC 组分别代表的是‘众信978’杂交种干旱处理后和水对照,通过这2 个组的比较,得到玉米敏感型品种中响应干旱的关键差异蛋白。对该比较组差异蛋白分别进行GO 富集和KEGG pathway 富集分析(图6、图7)。发现差异蛋白主要富集到的GO terms,其中4 个显著富集的生物过程分别为细胞大分子代谢过程、细胞蛋白质代谢过程、基因表达过程、翻译过程;5 个显著富集的细胞组分类别分别为细胞部分、细胞、大分子复合物、细胞器、核糖体;2 个显著富集的分子功能分别为核糖体结构组成和RNA结合。ZXD_ZXC 比较组差异蛋白的KEGG pathway分析表明,差异蛋白在核糖体通路中显著富集。
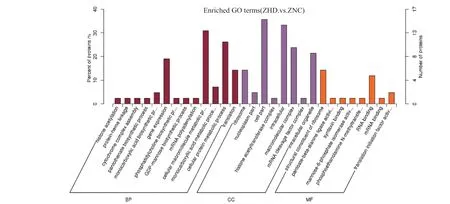
图6 ZXD_ZXC 比较组差异蛋白的GO 富集分析Fig.6 GO enrichment analysis of ZXD_ZXC comparison DAPs
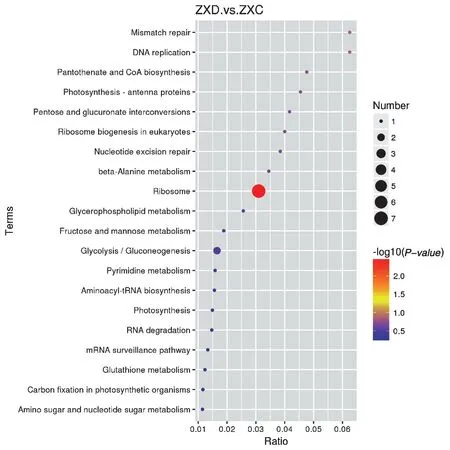
图7 ZXD_ZXC 比较组差异蛋白的KEGG 途径富集分析Fig. 7 KEGG pathway enrichment analysis of ZXD_ZXC comparison DAPs
3 讨论
面对全球气候变化,全球农业的主要目标是发展耐旱作物[9-10]。为此,充分了解与植物耐旱性有关的生理、生化和基因调控网络变得十分必要。在此,基于iTRAQ 技术对2 种耐旱性不同玉米杂交品种(耐旱型品种‘农单476’和敏感型品种‘众信978’)进行蛋白质组比较分析,分别对2 个品种在干旱胁迫条件下的差异表达蛋白进行了GO 和 KEGG 富集分析,报道了玉米耐旱的关键差异表达蛋白和调控途径。研究结果进一步加深了对玉米抗旱性调控机制的认识,为今后的定向克隆研究及耐旱关键基因的功能验证奠定了基础。
3.1 ‘农单476’在干旱条件下的KEGG 通路响应
本研究中参与内质网蛋白质加工途径的3个 差 异 表 达 蛋 白HSP70(Zm00001d012420_P001)、sHSF(Zm00001d031325_P001) 和UBX(Zm00001d005727_P001)均为上调表达(表3)。内质网是蛋白质加工折叠的关键场所。滑面内质网主要与脂质的合成与运代谢及储存和调节 Ca2+代谢及储存和调节 Ca2+浓度相关;糙面内质网则与外输性蛋白质的合成、加工修饰及转运过程密切 相关[11]。刘缨等[12]从人骨骼肌cDNA 文库中,分离,鉴定视网膜感光受体外周蛋白的结合蛋白(PBP)-DNAJ(HSP40)类分子伴侣,并指出DNAJ 可与HSP70(DNAK)协同发挥重要功能外。分子伴侣能参与内质网蛋白的折叠,BiP 蛋白是内质网腔内最丰富的分子伴侣[13],BiP 蛋白属于HSP70 蛋白家族,能与ATP 结合,并与含有J 结构域的蛋白质(J 蛋白)互作[14]。J 蛋白与BiP 蛋白互作后能刺激细胞内ATP 水解,促进BiP 蛋白捕获进入内质网腔内的多肽,伴随着核苷酸翻译的改变,新合成的多肽片段被缓慢释放到内质网腔中[15]。前人研究已经证实了次级代谢途径对于玉米耐旱的机制与作用[16]。通过对差异蛋白的KEGG pathway 富集分析,差异蛋白在内质网蛋白质加工和苯并恶唑嗪酮类化合物(BXs)的生物合成信号通路中最显著富集。王萱等[7]对玉米耐旱自交系YE8112 特有差异蛋白进行分析发现其主要参与内质网蛋白加工和色氨酸代谢通路,与本研究结果一致。杨杰[17]的研究发现强耐旱系PHBA6 响应干旱的差异蛋白中参与内质网蛋白质加工的蛋白占总差异蛋白的8%。管培燕[11]的研究发现拟南芥内质网蛋白SES1 具有分子伴侣活性,能参与蛋白质的折叠过程,具有调控拟南芥盐胁迫抗性的重要作用。上述分析结果表明内质网蛋白质加工途径是‘农单476’响应干旱胁迫的重要通路。
3.2 ‘众信978’在干旱胁迫下的KEGG 通路响应
核糖体是蛋白质合成的场所,是受非生物胁迫影响的基本生物学过程之一[18]。干旱胁迫条件下蛋白的表达情况与植物抗旱性有密切联系[19]。核糖体蛋白是组成核糖体的主要成分,在细胞内蛋白质生物合成中发挥重要作用,此外,还具有独立于蛋白质生物合成作用。核糖体具有参与DNA 修复、细胞发育调控和细胞分化等核糖体外功能[20-22]。核糖体蛋白L28 在马铃薯与青枯病菌互作过程中发挥作用[23];核糖体蛋白L14 受逆境调控,参与橡胶树对干旱胁迫的响应[24]。在本研究中‘众信978’在干旱条件下表达的差异蛋白显著富集在核糖体途径中,并且富集到核糖体途径的7 个蛋白(L23Ae、L27Ae、L35、L20、S18、L13e、L36e)均为下调表达(表4),造成该现象的原因为干旱胁迫对‘众信978’的影响。干旱胁迫条件导致‘众信978’的核糖体一些相关蛋白的表达下降,进而可能影响耐旱相关蛋白的合成。上述分析结果表明干旱胁迫可能通过影响核糖体蛋白的表达变化进而影响玉米杂交种‘众信978’的生长发育。
3.3 ‘农单476’转录与蛋白水平的干旱响应
之前的研究结果表明,干旱胁迫前后,杂交种‘农单476’在转录水平上筛选出70 个差异表达的基因[26],然而在蛋白水平上只有49 个显著差异的蛋白,其原因可能是部分差异基因并没有发挥实际的功能作用,也可能是检测和筛选手段造成的。但是有一点是确定的,即耐旱品种‘农单476’的差异基因和差异蛋白均少于敏感型品种‘众信978’。在转录结果中一个次级代谢产物生物合成相关的DEG 苯并恶嗪类1(Zm00001d048709) 差异表 达[25],与蛋白组研究结果一致,又互相佐证,表明次级代谢通路是‘农单476’响应干旱胁迫的一个 重要通路。
4 结论
对2 个抗旱性不同的玉米杂交种在V12 时期12 d 田间自然干旱处理后的玉米叶片进行了全面的iTRAQ 蛋白质组学比较分析。正常供水和干旱胁迫条件下总共有873 个DAPs 从4 个实验比较中被识别出来。‘农单476’的DAPs 主要富集在内质网蛋白质加工和次级代谢化合物的生物合成通路,而‘众信978’的DAPs 富集在核糖体的相关通路。本研究结果表明,‘农单476’玉米叶片具有较强的抵御干旱胁迫的能力,这是由于它们具有较高的sHSP 表达水平和较高的HSPs 表达水平;‘众信978’玉米叶片的抗旱相关蛋白均为下调表达。‘农单476’在转录及蛋白水平上均表现出更强的耐旱性。本研究加深了对V12 时期玉米叶片响应干旱的关键基因、蛋白和代谢途径的理解,为后续功能验证奠定了基础。
