山羊FGF18基因变异及其对羊绒性状的影响
2021-01-08张瑞国李少斌王继卿罗玉柱
张瑞国,李少斌,王继卿,刘 秀,罗玉柱
(甘肃农业大学 动物科学技术学院,甘肃省草食动物生物技术重点实验室,甘肃 兰州 730070)
成纤维细胞生长因子(Fibroblast growth factors, FGFs)是20世纪在大脑及垂体的组织提取物中发现的一种能促进成纤维细胞生长的活性物质[1],具有多种功能的信号蛋白,在生长发育、组织修复、脂肪代谢和器官形成中起着重要作用[2]。FGF18基因是FGF基因家族的一员,其编码的蛋白质属于分泌蛋白,在山羊上被定位于20号染色体,全长35 978 bp,含有5个外显子,编码207个氨基酸。FGF18通过与FGFs受体结合来进行信号传导,其功能的发挥依赖于FGFs受体的时空表达[3]。
FGF18在骨的形成中起着关键作用[4],在这方面的研究已经成为热点。进一步研究表明,FGF18在毛囊的发育和生长周期也起着重要的调控作用。吴瑜瑜[5]研究发现,在甘肃超细毛羊胎龄第87~108天,FGF18相对表达量逐渐升高,推测其在胎儿皮肤形成和发育过程中发挥了重要作用。在成年绵羊毛囊的生长旺盛期和缓慢期之间、颈部和腹部之间的FGF18基因表达量存在差异[6]。FGF18基因在小鼠角蛋白5-阳性角蛋白细胞中被有条件地敲除时,毛囊的静止期会变得非常短,从而使毛囊生长周期明显缩短,表明FGF18是调节毛囊静止期的关键信号[7]。曹校瑞等[8]研究表明,FGF18信号可以促进小鼠毛囊从静止期进入再生长期。
目前,有关FGF18基因多态性研究报道较少,并且集中在人类上。已有的研究表明,FGF18基因的遗传多态性与中国汉族膝骨关节炎[9]和人类的非综合征性唇腭裂[10]显著相关,但其多态性对毛发影响方面的研究尚未见相关报道。毛发是毛囊生长发育的产物,凡影响毛囊生长发育的信号通路或基因都能间接地影响毛发的结构和生长[11]。鉴于FGF18基因对毛囊生长发育有重要影响,并且在山羊上尚未有FGF18基因多态性的研究报道,探讨其变异及其对羊绒性状的影响具有重要意义。研究结果将为丰富山羊基因组研究及其遗传改良提供基础资料。
1 材料和方法
1.1 血样的采集
采集453只山羊血样,其中陇东绒山羊(甘肃环县)293只,柴达木绒山羊(青海漠河清牧场)110只,中卫山羊(甘肃景泰)50只。颈静脉采血,肝素钠(Heparin sodium)抗凝、-70 ℃保存。
1.2 羊绒性状数据的测定
收集12月龄陇东绒山羊羊绒样品,并收集记录耳号、性别、胎次和父系等信息。抓绒后对羊绒进行称质量得到产绒量;在山羊的体侧部测量绒层高度;采用投影显微镜法,用CU纤维细度仪测定绒纤维直径,测定工作在内蒙古农业大学动物科学学院生产与管理学实验室完成。
1.3 DNA的提取
采用北京全式金的EasyPure®Blood Genomic DNA Kit试剂盒,按照其说明书提取山羊血液全基因组DNA。
1.4 引物的设计
参照GenBank中山羊FGF18基因组的序列(序列号NC_030827),应用Primer 5软件设计3对引物分别扩增FGF18基因第3,4,5外显子(表1),引物由华大基因公司合成。
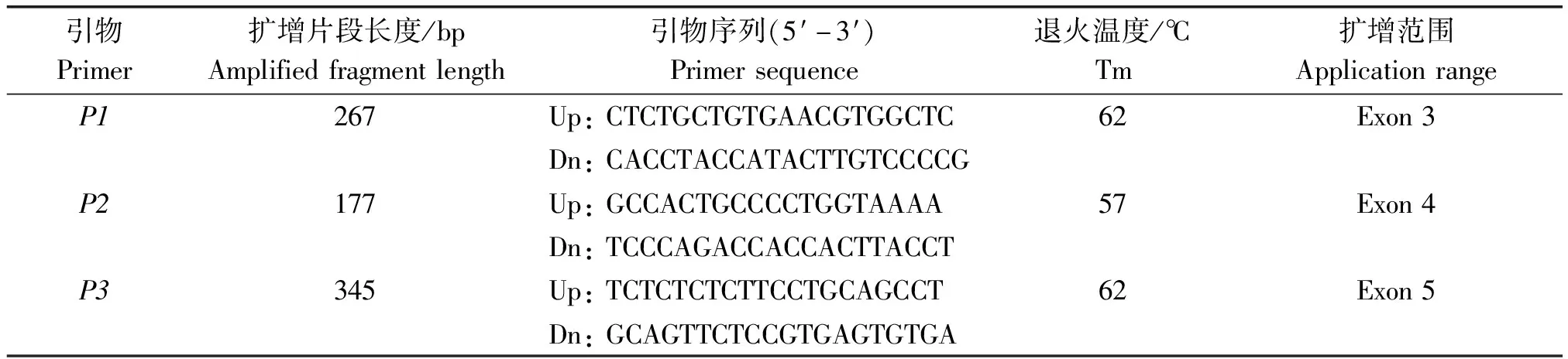
表1 FGF18基因各个外显子的引物序列
1.5 PCR扩增条件的建立
PCR反应总体积20 μL,上、下游引物(0.25 μmol/L)各0.8 μL,TaqDNA预混酶(南京诺唯赞生物技术有限公司)10 μL,dNTP 0.8 μL,ddH2O 7.6 μL。
PCR扩增程序:94 ℃预变性5 min;94 ℃变性30 s,按各引物退火温度退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸10 min,4 ℃保存。PCR扩增产物用1%琼脂糖凝胶电泳检测。
1.6 SSCP检测与测序
取2 μL的PCR产物,加入8 μL变性上样缓冲液(98%去离子甲酰胺,10 mmol/L EDTA,0.025%溴酚蓝和0.025%二甲苯氰),105 ℃变性5 min后,立即置于冰水混合物中,上样于14%的非变性聚丙烯酰胺凝胶(Acr∶Bis=37.5∶1),在230 V,15 ℃,0.5×TBE的条件下电泳18 h。电泳结束后,将非变性聚丙烯酰胺凝胶进行银染显色[12],然后置于A4纸上判型。将初步判定为纯合子样品的PCR 扩增产物进行直接测序,序列测定由上海生工生物有限公司完成。
1.7 数据的统计与分析
用MEGA 5进行序列比对,利用 Popgen 32.0软件计算有效等位基因数(Effective number of alleles, Ne)、遗传杂合度(Heterozygosity, He)、遗传纯合度(Homozygosity, Ho)和等位基因频率。采用 PIC 0.6 软件计算群体多态信息含量(Polymorphism information content, PIC)。应用SPSS 19.0一般线性混合模型分析基因型对陇东绒山羊羊绒性状的影响。研究发现,父系对所有羊绒性状均有影响(P<0.05),所以作为随机因素纳入模型。性别对羊绒性状有影响(P<0.05)或潜在影响(P<0.20),所以作为固定因素纳入模型。分析模型如下:
Yijk=μ+Gi+Fj+Sk+eijk
式中,Yijk为个体性状表型值,μ为群体均值,Gi为基因型效应,Fj为父系随机效应,Sk为性别效应,eijk为随机误差,下标i,j和k为个体序号。
2 结果与分析
2.1 PCR扩增产物的验证
山羊FGF18基因各扩增区的琼脂糖凝胶电泳检测结果见图1。引物P1、P2和P3的目的片段分别为267,177,345 bp,扩增产物与目的片段大小一致,且特异性较好,可做后续分析。
2.2 SSCP检测与测序
山羊FGF18基因不同外显子扩增区的SSCP检测结果显示,只在Exon 5区检测到3种带型,而在其Exon 3和Exon 4区只检测到1种带型(图2),表明Exon 3和Exon 4区无多态性。在山羊FGF18的Exon 5区检测到的3种带型,对应3种基因型EE、FF和EF,由E和F 2个等位基因构成。
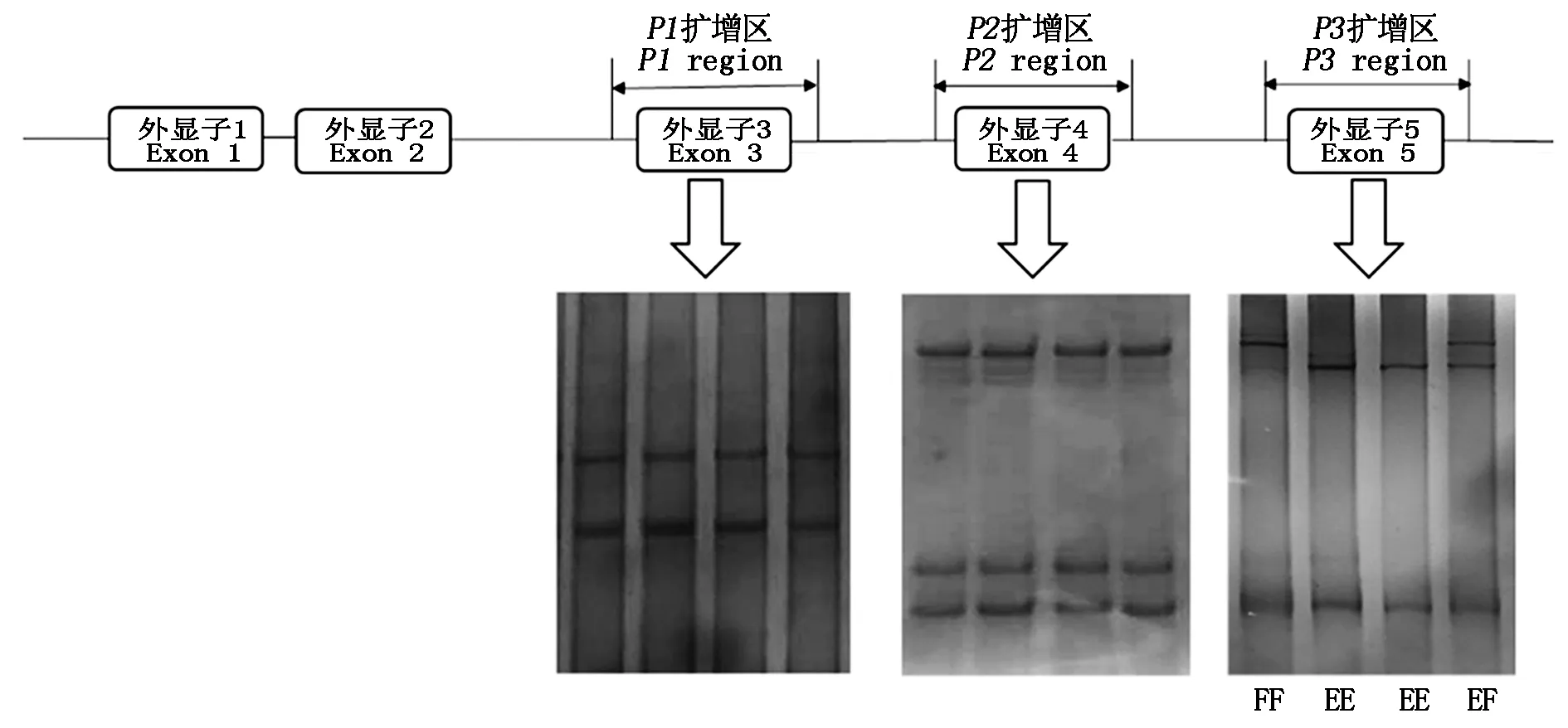
图2 山羊FGF18 SSCP检测
山羊FGF18基因的Exon 5区各基因型测序峰图和等位基因的序列比对结果见图3,4。结果显示,在山羊FGF18基因蛋白质编码区的498处碱基发生T→C的突变(c.498T>C)。氨基酸序列分析发现,该突变位点没有引起氨基酸种类的变化,属于同义突变。
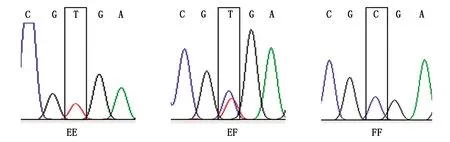
标有方框的区域表示核苷酸序列突变的区域。
2.3 遗传学分析
山羊FGF18基因 Exon 5的基因型频率和等位基因频率见表2。在3个山羊群体中,等位基因E均为优势等位基因。基因型在3个群体中的分布略有差异,陇东绒山羊、柴达木绒山羊和中卫山羊的优势基因型分别是EF(46.42%)、EE(54.50%)和EF(48.00%)。

标有下划线处表示引物序列。
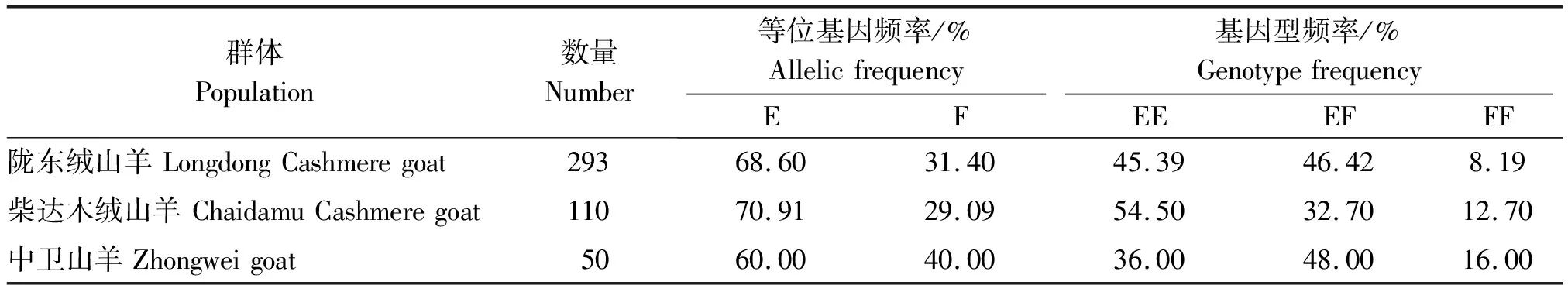
表2 FGF18基因Exon 5在3个山羊品种中的基因型频率和等位基因频率
陇东绒山羊、柴达木绒山羊和中卫山羊的FGF18基因的纯合度(Ho)、杂合度(He)、有效等位基因数(Ne)和多态信息含量(PIC)等遗传参数见表3。陇东绒山羊、柴达木绒山羊和中卫山羊的FGF18基因Exon 5区的PIC值都处于0.25~0.50,均表现为中度多态。经Hardy-Weinberg 平衡检验,FGF18基因Exon 5区在3个山羊群体中都处于Hardy-Weinberg 平衡(P> 0.05),说明FGF18基因Exon 5区在这3个山羊群体中都达到了遗传平衡。
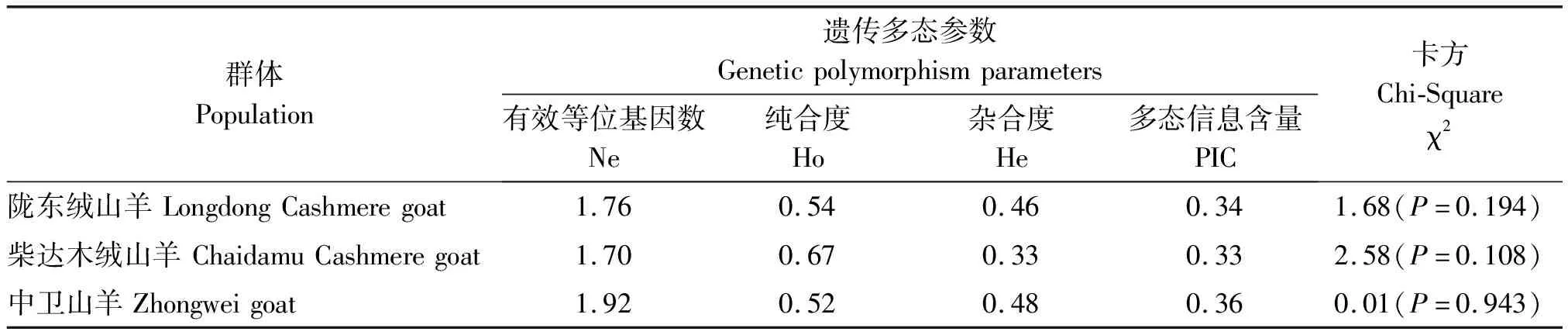
表3 FGF18基因在3个山羊品种中的遗传多态参数和Hardy-Weinberg 平衡检测
2.4 山羊FGF18不同基因型对陇东绒山羊羊绒性状的影响
从表4可知,FF型山羊的产绒量和绒层高度显著高于EE型和EF型山羊(P<0.05),EE型和EF型山羊的产绒量和绒层高度没有显著差异(P>0.05)。各基因型山羊的羊绒纤维直径没有显著差异(P>0.05)。
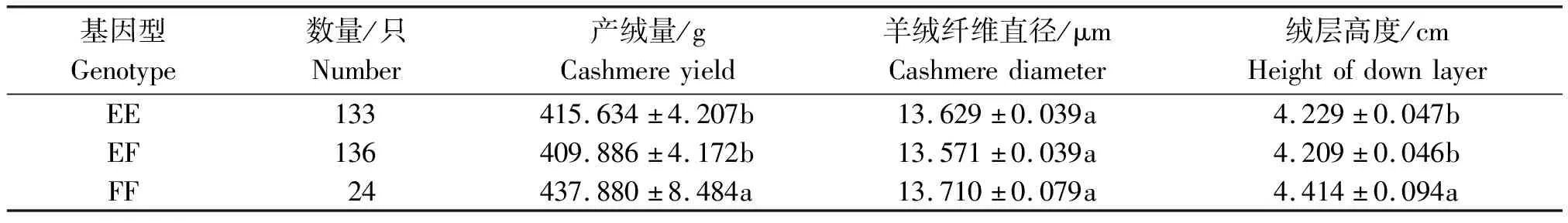
表4 山羊FGF18基因的不同基因型对陇东绒山羊羊绒性状的影响
3 讨论与结论
FGFs家族关于毛发方面的研究越来越被重视,已经有许多成员被发现与毛发的生长有关,如FGF1、FGF2、FGF5、FGF7、FGF18、FGF20、FGF22等FGFs家族成员均被发现在皮肤和毛囊细胞中表达,参与皮肤再生和毛发生长的过程[13]。研究该家族成员在山羊中的表达调控、遗传变异等分子生物学内容,具有重要的理论和实践意义。
本研究首次研究了山羊FGF18基因第3,4,5外显子的突变情况,结果显示,在山羊3个扩增区域仅检测到一个突变位点c.498T>C,表明该基因在这3个山羊品种中比较保守。有研究表明,FGF18的基因在进化过程中高度保守[14-15],核苷酸的变异程度较低,这与本试验结果相一致。FGF18基因的保守可能与其功能有关,它参与了一系列重要的生命活动过程,如脑的发育、腺垂体的分化和增殖、骨骼生长发育和胚胎发育等[16]。本研究结果表明,FGF18基因c.498T>C位点在陇东绒山羊、柴达木绒山羊和中卫山羊中都符合Hardy-Weinberg平衡,说明经过了选择和进化,该位点在3个群体中都达到了遗传平衡。c.498T>C位点对应的3种基因型在这3个山羊品种的分布存在差异,造成这种差异的原因有很多,比如其选育史、生活环境和品种特性等。本研究发现,FGF18基因的变异能同时对产绒量和绒层高度产生显著影响,这与绒层高度和产绒量之间表型相关[17]相一致,2个性状的表型相关系数是0.490(0.3<|r| ≤ 0.7),呈中度的正相关[17],推测其变异可能通过影响其中一个性状而对另一个性状产生间接影响。目前,关于FGF18基因变异对毛绒性状的研究未见报道,但有研究表明,FGF18基因能通过影响毛囊的周期来调节毛发的生长[8,13]。毛囊周期大致分为3个阶段,生长期、退化期和静止期[18],毛囊周期的变化会影响毛发的生长。Kawano等[13]把FGF18蛋白皮下注射到5只毛囊处于静止期的小鼠中,发现有4只小鼠的毛囊开始生长,说明FGF18能促进毛囊从静止期向再生长期的转变。Vandenabeele等[19]研究发现,FGF18蛋白的表达量在患有季节周期性侧面脱发的狗和正常狗中差异显著,表明FGF18基因与毛发的生长相关。以上研究都说明FGF18基因对毛发的生长起到重要的调控作用,提示其变异能对毛绒的性状产生一定影响。
虽然本研究在山羊FGF18外显子中发现的c.498T>C位点是同义突变,没有引起氨基酸的变化,但也可能对基因的功能产生影响。大量的研究结果表明,同义突变会从许多方面(基因剪接、mRNA稳定性、折叠和翻译、蛋白质折叠和基于miRNA的表达调控等)影响基因的功能,与各种疾病存在一定的联系[20]。FGF18对羊绒性状的影响也可能是与FGF10等附近基因连锁造成的。山羊的FGF10和FGF18基因都位于同一条染色体上,并且FGF10基因也能调控毛囊的周期[21]。
绒山羊产绒量、绒纤维直径、绒层高度是衡量生产性能的3个重要指标。育种中,在绒纤维直径保持不变的情况下,提高产绒量、增加绒层高度是十分必要的。产绒量是衡量山羊经济价值的重要指标,在绒山羊的鉴定和选种中起到指导作用。绒层高度也是山羊重要的经济性状,与产绒量联系紧密。在绒山羊生产中,产绒量和绒层高度提高是符合生产需求的。FF型山羊的产绒量和绒层高度均高于其他基因型山羊,提示若以这2个性状为选育目标,应该提高群体中FF型山羊的比例。在3个山羊群体中,FF型山羊的比例均是最低的,说明山羊FGF18基因c.498T>C变异位点受人工选育的程度较低。
本研究在陇东绒山羊、柴达木绒山羊和中卫山羊的第3,4,5外显子只检测到了一个突变位点c.498T>C,该位点在这3个山羊群体中均呈中度多态,并且都符合Hardy-Weinberg平衡。在陇东绒山羊中,c.498T>C位点对产绒量和绒层高度影响显著,提示若以这2个性状为选育目标,FGF18基因可以作为分子改良的候选基因。
