深圳东涌红树林生态位和空间结构二元分布研究
2021-01-05丘建煌许恒涛王章芬张红洁韩维栋
丘建煌 许恒涛 王章芬 张红洁 韩维栋









摘要: 以深圳东涌红树林群落为研究对象,运用重要值、生态位宽度及生态位指数对深圳东涌红树林不同群落中主要种群的生态位特征进行分析,同时采用空间结构指数交互分析的方法探讨了各种群空间结构的二元分布特征。结果表明:(1)各种群的生态位宽度排序为桐花>秋茄>海漆>黄瑾,说明各种群对环境各有各自的适应能力。(2)总体上各种群间的生态位重叠值并不大,最大的桐花和秋茄生态位重叠值也值为0.04~0.05。两者处于动态平衡的关系,说明东涌红树林各种群间有对资源的竞争,但竞争力度较小,有比较明显的分布区域和界限。(3)从空间结构一元分布来看,该地红树林群落总体上处于中度混交状态,且随机分布,大多林木处于中庸状态。(4)从空间二元分布来看,群落Ⅰ、Ⅱ、Ⅲ总体上处于中度混交且随机分布,大多林木处于中庸状态;群落Ⅳ则总体上处于零度混交且劣势,大多处于不均匀分布。
关键词: 深圳东涌; 红树林; 生态位; 空间结构; 二元分布
中图分类号: S 718. 54 + 2 文献标识码: A 文章编号:1001 - 9499(2021)04 - 0043 - 06
紅树林是低纬度热带、亚热带地区特有的植物群落[ 1 ],在沿海海岸中发挥着防风减灾,加固海岸,净化海洋、维护海洋生态平衡等重要作用[ 2 ]。深圳市也是我国红树林主要分布区域,主要呈带状分布于沿海地带。近年来,随着国家对红树林保护的重视以及深圳致力于打造园林城市,在红树林的保护和恢复方面开始重视起来,使得红树林的面积得到了一定的恢复。国内对红树林物种多样性的研究较多,大多对中国红树林生态系统的植物种类、植物多洋性、功能及其保护的研究[ 3 ],但是对红树林的空间结构的研究还比较少。因此,本文采用广泛应用的生态位原理以及林木空间结构参数,分析了解东涌红树林区种群的组成结构、内部关系以及与其他种之间的关系,揭示各个种的生态适应机理,以期为后续的红树林保育提供可行性指导。
1 场地概况
深圳市大鹏新区东涌红树林湿地地处深圳东南大鹏半岛的最南端,位于大鹏半岛自然保护区试验区范围内,同时也在深圳市基本生态控制线范围内,沿海岸风景优美。这是我国面积最大的红树林自然保护区,区内物种繁多,是鸟类的天堂,有着丰富的浅海生物资源。该保护区内红树、半红树树种丰富,主要包括秋茄(Kandelia obvata),桐花树(Aegiceras corniculatum)、白骨壤(Avicennia marina)、木榄(Bruguiera gymnorrhiza)、海漆(Excoecaria agallocha )等十余种主要红树树种。
2 调查与研究方法
2. 1 调查方法
在深圳东涌红树林湿地中选择海漆+秋茄+桐花树(群落Ⅰ)、桐花树+海漆+秋茄+黄瑾(群落Ⅱ)、桐花树+秋茄(群落Ⅲ)、海漆+桐花树(群落Ⅳ)4种不同的群落类型分别取20 m2的样方,根据地块的实际情况,群落Ⅰ、Ⅱ、Ⅲ分别设置4块面积为5 m×5 m的样地,群落Ⅳ设置2块10 m×10 m的样地,各样地之间除树种组成不同外,其他立地条件基本一致。调查每个样方内乔木树种的种类、数量、相对位置(x,y)以及胸径、树高、冠幅等生长因子。
2. 2 研究方法
2. 2. 1 生态位计算
(1)乔木层重要值=(相对密度+相对频度+相对显著度)/3×100%(1)
式中:相对密度 R=ni/N×100%;ni为某种植物的个体数,N为全部植物个体数;相对频 度 R= Fi/F×100%,Fi=ni/N×100%;ni为某一物种出现的样方数,N为样方总数。Fi为某一物种的频度,F为所有物种的频度和。相对显著度R=Gi/G×100%;Gi为某个树种的胸高断面积,G为全部树种的总胸高断面积[ 5 - 6 ]。
(2)生态位宽度计算方法
Levins 生态位宽度计算式、Shannon-Wiener生态位宽度指数进行测度:
式中:B(sw)i为物种i的Shannon-Wiener指数计算的生态位宽度;B(L)i为物种i的Levins指数计算的生态位宽度;Pij是物种i在第j资源位上的重要值之比,r为样方数。其中nij为物种i在第j样方的重要值[ 4 - 5 ]。
(3)Levins生态位重叠指数计测:
式中:Lih为物种i重叠物种h的生态位重叠指数;Lhi为物种h重叠物种i的生态位重叠指数;B(L)i为物种i的Levins生态位宽度指数[ 4 - 5 ]。
2. 2. 2 空间结构计算方法
(1)混交度(林种空间隔离指数)[ 6 ]
混交度(Mi)是用来说明混交林中树种空间隔离的程度。它被定义为参照树i的n株最近相邻木中与参照树不属同种的个体所占的比例,用公式表示为[ 8 - 9 ]:
式中:Vij是一个离散型的变量,其值定义为:当参照树i与第j株相邻木非同种时Vij=1,反之,Vij=0。根据惠刚盈的研究[ 7 - 8 ],n取为4时,Mi较能准确反映林木不同树木在林地空间的分布,本研究取n=4。其中M为0.00(零度混交)、0.25(弱度混交)、0.50(中度混交)、0.75(强度混交)、1.00(极强度混交)。
(2)大小比数(林木竞争指数)[ 9 ]
树木成林郁闭后,林木之间形成竞争,产生林木大小的分化。大小比数(Ui)被定义为大于参照树的相邻木所考察的全部最近相邻木的比例,用公式表示为[ 8 - 9 ]:
式中:若参照树比相邻木小,Kij=1,否则Kij=0[ 8 - 9 ]。其中U为1.00(绝对劣势)、0.75(劣势)、0.50(中庸)、0.25(亚优势)、0.00(优势)。
(3)角尺度(林木空间分布格局指数)[ 10 ]
角尺度是反映树木分布状况的一种空间指标,是描述林木个体分布格局的结构参数,它反映了相邻木围绕参照树的均匀性。其计算公式为[ 7 - 8 ]:
式中:n 为相邻木株数;Zij为角度尺取值变量,它是离散型的,当 ai小于标准角a0=72时, Zij=1,否则 Zij=0[ 7 - 8 ]。其中W为0.00(绝对均匀)、0.25(均匀)、0.50(随机)、0.75(不均匀)、1.00(集聚)。
在红树林将角尺度、混合度以及大小比数两两联合(分别是角尺度和混交度、混交度和大小比数、 角尺度和大小比数三种),分别计算出一个参数在另一个参数上的对应取值(共 25 种组合),将其除以样地中林木的数量即可获得2个参数的组合频率值,从而形成二进分布[ 11 - 13 ]。
计算每棵树的角尺度、大小比数和混交度,使用 Excel 透视表计算群落的单向和双向分布频率,并使用 Excel 绘制 X-Y 双向分布图[ 14 ]。
3 结果与分析
3. 1 红树林种群中各种群的重要值
根据深圳市东涌红树林群落实地调查资料,得到各群落树种位置图并计算出主要树种的重要值(表1),群落Ⅰ中黄瑾重要值最大;群落Ⅱ中的桐花树重要值最大;群落Ⅲ中的秋茄重要值最大;群落Ⅳ中海漆重要值最大,说明它们在各资源位中具有一定的优势。各物种重要值因资源位不同而不同,如桐花树在群落Ⅰ为0.260 2,而在群落Ⅱ为0.402 5。
3. 2 红树林种群中各种群的生态位宽度分析
从表2和表3可见,虽然 Levins 和Shannon-Wiener 生态位宽度公式计算出的数值结果存在差异,但它们的规律是一样的[ 15 ]。不管是生态位宽度还是BSW、BL都显示种群的生态位宽度值为桐花>秋茄>海漆>黄瑾。从总体上看,桐花树、秋茄、海漆分布都较广,生态位宽度相差不大且各有各的分布特点。主要呈现由滩涂到陆地:桐花树←秋茄←海漆。桐花树主要分布于滩涂沿岸,且树高较低,主要是为了与秋茄竞争光照,秋茄则与桐花树处于相邻的位置,也有相互重叠,总体而言两者处于相互竞争的关系;海漆属于大型的乔木类型,在资源的利用上明显相对于秋茄和桐花树有很大的优势。这三种红树林植物在每个资源位中都有出现,表明其在林分内利用资源位较为充分,对资源的利用能力较强;黄瑾生态位宽度值相对于其他三种来说生态位宽度值较小,说明它们分布点较少,利用资源的能力较弱。
3. 3 红树林种群中各种群的生态位重叠分析
如表4所示,种群之间共有6对生态位重叠,各种群间的生态位重叠值Lih、Lhi分别为0~0.041 7、0~0.049 1。各种群间的生态位重叠值不大,均在0.01~0.05,说明它们利用生境的模式有相似之处。生态位重叠最大的是桐花—秋茄种群,重叠值为0.04~0.05,重叠程度适中,说明该种群比较稳定[ 18 ]。结合生态位宽度可以看出,生态位重叠较大的物種生态位宽度值也较大。从整体上看,红树林种群生态位重叠值较小,说明各种间对资源的竞争较小,各种群间有较为明显的分布区域和分界线。
3. 4 不同红树林群落空间结构的一元分布
3. 4. 1 桐花树+秋茄+海漆+黄瑾种群结构参数的一元分布
由图1可以看出,群落Ⅰ的混交度为0.53,总体上处于中度混交状态,但是各等级分布差距并不是很大,黄瑾树决定了整个林分的混交程度,树种主要呈现出单种树种的聚集。角尺度的各等级分布差距较为明显,呈先上升后下降再上升的趋势,平均角尺度为0.62,该群落近乎是随机分布。林分的大小比数规律比较明显,各等级的分布频度相差不大,平均大小比数为0.49,说明该群落多数林木处于中庸状态。
3. 4. 2 桐花树+秋茄+海漆种群结构参数的一元分布
由图2可以看出,群落Ⅱ的平均混交度为0.51,各等级间差异较为明显,总体处于中度混交状态。角尺度的规律较为明显,呈先上升后下降的趋势,平均角尺度为0.57,群落总体呈现出随机分布的情况。大小比数各等级的差异不大,各等级的分布频度约为20%,平均大小比数为0.47,说明该群落多数林木处于中庸状态。
3. 4. 3 桐花树+秋茄种群结构参数的一元分布
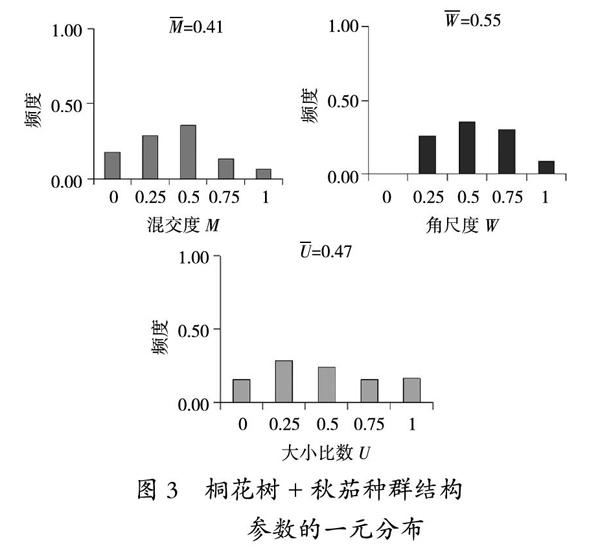
由图3可以看出,群落Ⅲ混交度的规律较为明显,大致呈现先上升后下降的趋势,平均混交度为0.41,说明该群落处于中度混交程度。在角尺度中,随机分布和不均匀分布比例较大,各占65%和26%,平均角尺度0.55,说明整个群落处于随机分布。平均大小比数0.47,表明该群落多数树木近乎处于中庸状态。
3. 4. 4 桐花树+海漆种群结构参数的一元分布
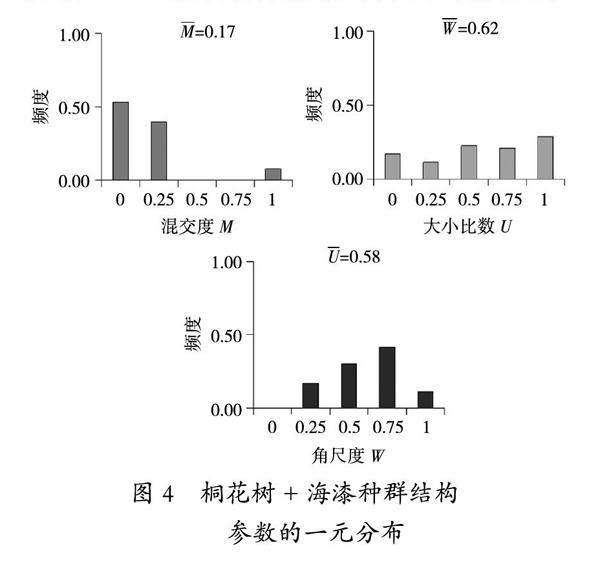
由图4可以看出,群落Ⅳ零度混交和弱度混交比例较大,两者总占92%。平均混交度为0.17,说明该群落处于弱度混交。林分优势树种海漆的混交程度决定了整个群落的混交程度,群落呈现单一树种的聚集。角尺度在各等级差异较为明显,其中随机分布和不均匀分布占比较大,分别为30%和42%,平均角尺度为0.62,表明整个群落处于随机分布的状态。平均大小比数为0.58,除了亚优势等级占比较小和绝对劣势占比较大,其余等级均在20%左右,表明该群落近乎处于中庸状态。
3. 5 不同红树林群落空间结构的二元分布
3. 5. 1 不同群落红树植物混交度-角尺度二元分布
从图5可以看出,在同一混交度下,除了群落Ⅳ外,相对频度大体呈现先增后减的趋势,且最大值均在W=0.5上,即群落Ⅰ、Ⅱ、Ⅲ在相同的混交条件下大多林木处于随机分布;群落Ⅳ的最大值在W= 0.75上,表明大多林木处于不随机分布。在同一角尺度条件下,群落Ⅰ、Ⅱ、Ⅲ相对频度同样呈现先增后减的趋势,且最大值M=0.5,即在相同的混交程度下,3个群落的大多树林木处于中度混交。群落Ⅳ的变化趋势呈逐渐下降趋势,最大值M=0,即在相同角尺度等级的条件下,同一分布格局中林分个体大多处于零度混交。
3. 5. 2 不同群落红树植物混交度-大小比数二元分布
从图6可以看出,在同一混交等级下,群落Ⅰ的相对频度随着大小比数的增加呈先增后减的趋势,最大值在U=0.75上,即相同混交条件下,林木个体大多处于劣势状态;群落Ⅱ、Ⅲ相对频度同样呈先增后减的趋势,但最大值主要集中在U=0.25上,即在相同混交条件下,林分中树木个体大多处于亚优势状态;群落Ⅳ的变化差异较大,最大值U=1,即在相同混交条件下,林分中树木个体大多处于绝对劣势状态。在同一大小比数等级下,群落Ⅰ、Ⅱ、Ⅲ,相对频度同样呈现先增后减的趋势,且最大值M=0.5,即在相同的混交程度下,3个群落的大多树林木处于中度混交。群落Ⅳ的变化趋势呈逐渐下降,最大值M=0,即在相同角尺度等级的条件下,同一分布格局中林分个体大多处于零度混交。
3. 5. 3 不同群落红树植物角尺度-大小比数二元分布
从图7可以看出,同一角尺度等级下,群落Ⅰ的相对频度随着大小比数的增大呈先增后减的趋势,最大值主要集中在U=0.75上,即相同混交条件下林木个体大多处于劣势状态;群落Ⅱ、Ⅲ的相对频度随着大小比数的增大均呈先增后减的趋势,且最大值均为U=0.25,分别占比43%和62%,表明相同角尺度条件下,同一分布格局中林分个体大多处于亚优势;群落Ⅳ的相对频度随着大小比数的增加大致呈先减少后递增的趋势,最大值主要在U=1上,占比28%,即在相同混交条件下,同一分布格局中树木个体大多处于绝对劣势的状态。在同一大小比数等级下,群落Ⅰ、Ⅱ、Ⅲ的相对频度随着角尺度的增加,呈先增后减的趋势,且最大值均在W=0.5,即在相同大小比数等级的条件下,同一分布格局中林分个体大多处于随机分布;群落Ⅳ的相对频度则为逐渐减少的趋势,最大值在W=0.75,表明在相同大小等级的条件下,同一分布格局中林分个体大多处于不均匀分布。
4 结 论
4. 1 通过计算重要值以及各种群的资源位得出各种群的生态位宽度和生态位重叠[ 16 ],桐花树、秋茄和海漆具有较大的生态宽度值,而黄瑾的生态位宽度值则比较小。有研究表明,生态宽度值越大,物种的特化程度就越小,说明该物种对生态环境的适应能力越强[ 17 ],其在群落里的分布范围就越广泛。从桐花树的生态宽度值来看,其在各种群内均有分布且分布规律较明显,这与实地调查的结果相符,说明桐花树的适应能力较强。
4. 2 从生态重叠值的情况来看,种群间的生态位重叠值并不大,最大的秋茄—桐花重叠值也值为0.04~0.05,这2个物种也只是出于动态平衡,即物种与物种之间有明显的分布区域和界限[ 18 ]。秋茄-桐花2个物种的生态宽度也是最大的,正好对应了生态宽度越大,生态位重叠就越大的研究结果[ 19 ]。
4. 3 从空间的一元分布结构结果来看,除群落Ⅳ为弱度混交外,群落Ⅰ、Ⅱ、Ⅲ均为中度混交,近乎随机分布,大多林木属于中庸状态。有研究表明,随着自然的不断演替更新,假如受到的外界干擾破坏比较少,群落的随机分布状态是群落分布格局的常态[ 20 - 21 ]。随着时间的不断推移变化,林分中的混交程度也会越来越大,在调整变更植物群落林分格局时,应该对此进行调整[ 22 ]。从东涌研究地情况来看,整体上处于中度混交,且随机分布,大多数林木处于中庸状态,表明该地受到的外界干扰较少。
4. 4 从空间的二元分布结构结果来看,群落Ⅰ总体上处于中度混交且劣势,大多林木属于随机分布,偏向聚集和高度混交的林木比较常见;群落Ⅱ总体上处于中度混交且偏优势,大多林木属于随机分布,偏向高度混交和优势林木较为常见;群落Ⅲ总体上处于中度混交劣势,大多林木属于随机分布;群落Ⅳ总体上处于零度混交且劣势,大多林木属于不均匀分布。在这些林分空间中,高度混交且随机分布的林木虽然占有一定比例,但与理想的林木空间结构相差甚远[ 23 ]。通过分析上述空间结构,不但可以改善现有红树的林分空间结构,也可以为东涌即将恢复的红树林生态结构提供可行性的依据。
参考文献
[1] 梁士楚. 广西英罗湾红树植物群落的研究[J]. 植物生态学报, 1996(4): 310 - 321.
[2] 单家林, 郑学勤. 海南岛红树林植物区系组成与特征[J]. 广东林业科技, 2005(2): 41-45.
[3] 林益明, 林鹏. 中国红树林生态系统的植物种类、 多样性、功能及其保护[J]. 海洋湖沼通报, 2001(3): 8 - 16.
[4] 陈俊, 艾训儒, 姚兰, 等. 木林子川陕鹅耳枥+青冈-水马桑群落优势种生态位与种间关联[J]. 湖北民族学院学报(自然科学版), 2017, 35(3): 255 - 260.
[5] 冯宜明, 陈学龙, 齐瑞, 等. 甘肃亚高山云杉人工林下植物种群生态位特征[J]. 草业科学, 2018, 35(4): 807 - 815.
[6] Hui G Y,Hu Y B. Measuring species spatial isolation inmixed forests[J]. For Res, 2001, 14 (1): 177 - 181.
[7] 胡艳波, 惠刚盈, 戚继忠, 等. 吉林蛟河天然红松阔叶林的空间结构分析[J]. 林业科学研究, 2003(5): 523 - 530.
[8] 张岗岗, 王得祥, 柴宗政, 等. 秦岭中段华北落叶松人工林空间结构的二元分布特征[J]. 西北农林科技大学学报(自然科学版), 2014, 42(9): 33 - 40.
[9] 张岗岗, 王得祥, 张丛珊, 等. 小陇山百花林场次生松栎混交林群落及优势种群空间结构特征[J].东北林业大学学报, 2015, 43(8): 40 - 45.
[10] Hui G Y, von Gadow K, Albert M. A new parameter forstand spatial structure neighborhood comparison[J]. For Res, 1999,12(1): 1 - 6.
[11] 梁文俊, 魏曦, 赵伟文, 等. 基于二元分布空间结构参数的林分结构研究[J]. 河南农业大学学报, 2018, 52(4): 540 - 544.
[12] 李远发. 林分空间结构参数二元分布的研究[D]. 北京:中国林业科学研究院, 2013.
[13] 彭辉, 周红敏, 惠刚盈. 萌生杉木林空间结构的二元分布特征[J]. 南京林业大学学报(自然科学版), 2017, 41(4): 136 - 140.
[14] Hui G Y, von Gadow K, Albert M . The neighborhoodpattem - a new structure parameter for describingdistribution of forest treeposition[J]. SciSilvsin, 1999, 35(1): 37 - 42.
[15] 万娟, 叶立新, 丁文勇, 等. 浙江省自然保护区常绿阔叶林优势种群的生态位特征[J]. 浙江大学学报(理学版), 2014, 41(6): 725 - 734.
[16] 胡喜生, 洪滔, 宋萍, 等. 万木林木荷群落优势种的生态位[J]. 亚热带农业研究, 2006(4): 285 - 289.
[17] 李德志, 石强, 臧润国, 等. 物种或种群生态位宽度与生态位重叠的计测模型[J]. 林业科学, 2006(7): 95 - 103.
[18] 马立宇, 黄剑坚. 特呈岛红树植物群落特征及生态位研究[J].防护林科技, 2019(7): 9 - 12.
[19] 赵永华, 雷瑞德, 何兴元, 等. 秦岭锐齿栎林种群生态位特征研究[J]. 应用生态学报, 2004(6): 913 - 918.
[20] 董灵波, 刘兆刚. 樟子松人工林空间结构优化及可视化模拟[J]. 林业科学, 2012, 48(10): 77 - 85.
[21] 周红敏, 何必庭, 彭辉, 等. 萌生杉木林空间结构特征研究[J]. 林业科学研究, 2015, 28(5): 686 - 690.
[22] 张家城, 陈力, 郭泉水, 等. 演替顶极阶段森林群落优势树种分布的变动趋势研究[J]. 植物生态学报, 1999(3): 3 - 5.
[23] 黄剑峰, 谭伟, 柴宗政, 等. 黔中马尾松近熟林空间结构特征及其调控[J]. 浙江农林大学学报, 2019, 36(4): 749 - 756.
第1作者简介: 丘建煌(1995-), 男, 在读研究生, 主要从事植物资源利用与开发研究。
通讯作者: 韩维栋(1963-), 男, 教授, 主要从事树木学教学与林学研究工作。
收稿日期: 2020 - 11 - 20
(責任编辑: 张亚楠)
