氧化对羊肉肌原纤维蛋白分子与理化特性的影响
2020-12-31张海璐安凤平陆健康
张海璐,黄 翔,杨 燃,安凤平,陆健康*,黄 群,*
(1.贵州医科大学食品科学学院,贵州 贵阳 550025;2.福建农林大学食品科学学院,福建 福州 350001;3.塔里木大学生命科学学院,新疆 阿拉尔 843300)
红肉、家禽和鱼类肌肉的主要蛋白质是肌原纤维蛋白,约占总蛋白质的50%~55%[1]。肉制品在加工及贮藏过程中不可避免地受到环境影响,使肌原纤维蛋白和基质蛋白都受到氧化的影响,引起肌原纤维蛋白分子结构的改变,导致肉制品的理化特性如持水性、氨基酸的改变[1-2]。与脂肪氧化反应类似,蛋白氧化也是由自由基链式反应所引起,主要由起始、传递、终止3 个阶段组成[3-4]。加工中各类不规范操作引入的金属离子、与带辐射物质接触或加入氧化酶等因素都可能促进蛋白氧化的发生[5-7]。目前已有大量关于蛋白氧化的研究报道,将羊肉进行羟自由基氧化后研究其糜流变与凝胶特性,羊肉出现持水性下降、凝胶特性降低等现象,说明蛋白氧化使蛋白结构发生变化[8]。Xiong Youling L.等[9]研究了3 个不相同的氧化体系对猪肉肌原纤维蛋白分子结构和化学键的影响。Sante-Lhoutellier等[10]报道了氨基酸氧化、蛋白质聚集以及蛋白质氧化与蛋白水解之间的相互关系,发现蛋白质的氧化会诱导各种氨基酸的修饰并将其转化为羰基衍生物。本研究主要针对羊肉中肌原纤维蛋白,采用体外模拟不同程度的氧化,探究不同氧化时间对羊肉肌原纤维蛋白分子与理化特性的影响,以期为羊肉及其制品的加工和贮藏提供理论依据。
1 材料与方法
1.1 材料与试剂
新鲜羊后腿肉(8 月龄白山羊)购自福州市山野农贸市场。
溴酚蓝、三羟甲基氨基甲烷(Tris)、三氯化铁、抗坏血酸、过氧化氢 国药集团化学试剂有限公司;5,5’-二硫代双(2-硝基苯甲酸)(5,5’-dithiobis(2-nitrobenzoic acid),DTNB)、2,4-二硝基苯肼(2,4-dinitrophenylhydrazine,DNPH) 北京索莱宝科技有限公司;N,N,N’,N’-四甲基乙二胺(N,N,N’,N’-tetramethylethylenediamine,TEMED) 上海生工生物工程股份有限公司。
1.2 仪器与设备
UV-1780紫外分光光度计、傅里叶变换红外光谱(Fourier transform infrared spectroscopy,FTIR)仪岛津仪器(苏州)有限公司;F-4600荧光分光光度计、LA8080氨基酸自动分析仪 日本日立公司;Omni多角度粒度与高灵敏度Zeta电位分析仪 美国布鲁克海文仪器公司;3000粒度仪 英国马尔文仪器有限公司;JY600E电泳仪 北京君意东方电泳设备有限公司。
1.3 方法
1.3.1 原料预处理
新鲜羊后腿肉于0~4 ℃预冷1 h,剔除多余脂肪和结缔组织,将羊肉分割成大小、厚薄均等的肉块(1 cm×1 cm×1 cm),用流水进行清洗,纸巾吸干表面水分后置于4 ℃冰箱中临时保存。将处理后羊肉打浆6 次(每次30 s)后,打浆后肉糜密封并于4 ℃冷藏备用。
1.3.2 肌原纤维蛋白提取
肌原纤维蛋白的提取参照孙金辉等[11]的方法并稍作修改。称取适量羊肉糜,按肉糜质量加入4 倍体积的0.05 mol/L磷酸盐缓冲液(pH 7.0),高速匀浆1 min后,冷冻离心(3 000×g、15 min,后同),取残渣再重复提取两次。将残渣与4 倍体积的0.1 mol/L NaCl溶液混合,高速匀浆30 s后冷冻离心;所得残渣重复前述处理两次,将所获残渣与4 倍体积的0.1 mol/L NaCl溶液混合,用蒸煮过的4 层纱布过滤;调节pH值至6.2,冷冻离心所得肌原纤维蛋白于4 ℃冷藏,一周内用完。
1.3.3 肌原纤维蛋白氧化处理
肌原纤维蛋白氧化处理参考Li Shugang等[12]的方法并适当修改。将提取的肌原纤维蛋白均分成6 份,置于Fenton氧化体系(1.0 mmol/L FeCl3、0.1 mmol/L抗坏血酸和10 mmol/L H2O2)中,使其最终质量浓度为10 mg/mL,密封避光于4 ℃恒温水浴中分别氧化1、2、3、4、5 h,获得不同氧化时间的氧化蛋白质,以未经氧化的肌原纤维蛋白为对照。将肌原纤维蛋白样品一部分置于-80 ℃后冻干处理,用于十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)和氨基酸含量的测定;另一部分置于4 ℃冷藏,用于测定其他指标。
1.3.4 羰基、总巯基、二聚酪氨酸含量及表面疏水性测定
羰基含量参照李德海等[13]的2,4-二硝基苯肼反应方法测定。
总巯基含量测定参照Wang Yu等[14]的方法,计算时摩尔消光系数为13 600 L/(mol·cm)。
二聚酪氨酸含量测定参照李玲等[15]的方法并稍作修改。肌原纤维蛋白用磷酸盐缓冲液(含0.6 mol/L KCl,pH 6.0)稀释至1 mg/mL,5 000×g冷冻离心3 min,利用荧光分光光度计测定上清液的荧光强度(激发波长325 nm、发射波长420 nm),并以荧光强度表征二聚络氨酸含量。
表面疏水性测定参照Chelh等[16]的方法,肌原纤维蛋白用磷酸盐缓冲液(含0.6 mol/L NaCl,pH 6.25)稀释至5 mg/mL,取1 mg/mL溴酚蓝溶液0.2 mL添加到1 mL 5 mg/mL蛋白溶液中,混匀后,离心15 min(4 ℃、5 000×g),取上清液稀释10 倍,利用紫外-可见分光光度计测定595 nm波长处的吸光度,对照组为无蛋白溶液添加的磷酸盐缓冲液。表面疏水性使用溴酚蓝结合量表示,具体按下式计算。

1.3.5 紫外吸收光谱测定
将样品用磷酸盐缓冲液(pH 7.0)稀释至1 mg/mL后,置于UV-1780紫外分光光度计中测定紫外吸收光谱,设置扫描波长范围为240~340 nm,扫描速率为2 nm/min。
1.3.6 内源荧光光谱测定
内源荧光光谱的测定参照Cao Yungang等[17]的方法并略作修改。用磷酸盐缓冲液(含0.6 mol/L NaCl,pH 6.7)将肌原纤维蛋白稀释至0.1 mg/mL后,置于F-4600荧光分光光度计中测定溶液的内源荧光光谱,设置激发波长280 nm,发射波长300~400 nm,电压为600 V。
1.3.7 FTIR光谱测定
取适量冻干样品与溴化钾按质量比1∶100充分混均,用玛瑙研钵碾磨,制成溴化钾压片。置于FTIR仪中,设置波数范围为400~4 000 cm-1,分辨率为4.0 cm-1,扫描次数为32,并使用PeakFit 4.12软件对图谱酰胺I带1 600~1 700 cm-1范围内进行曲线拟合,计算各二级结构的相对含量。
1.3.8 Zeta电位测定
Zeta电位测定参照Cai Luyun等[18]的方法并略作修改。肌原纤维蛋白溶于20 mmol/L磷酸盐缓冲液(含0.6 mol/L NaCl,pH 6.7)中得到质量浓度为0.2 g/100 mL的溶液,用0.22 μm的醋酸膜(水系)进行过滤,采用Omni多角度粒度与高灵敏度Zeta电位分析仪测定样品的Zeta电位,使用35 mW固态激光器(λ=660 nm)在25 ℃下测定,并将蒸馏水作为溶剂,每组样品平行测定5 次。
1.3.9 粒径测定
粒径测定参照Feng Meiqin等[19]的方法利用粒度仪进行测定。使用蒸馏水作为分散剂,所有测定设5 次平行,折射率为1.46,吸收系数为0.01,并且颗粒为非球形。D(v, 0.1)、D(v, 0.5)、D(v, 0.9)分别为10%、50%、90%的颗粒小于该尺寸的颗粒尺寸。
1.3.10 蛋白组成测定
采用SDS-PAGE进行蛋白组成分析,分离胶质量分数为12%,浓缩胶质量分数为4%,分别取肌原纤维蛋白10 μg,放入4×上样缓冲液,沸水浴5 min;先80 V恒压使溴酚蓝移动至浓缩胶与分离胶分界处,再换电压至120 V,恒压至溴酚蓝移动至胶底。电泳结束后进行考马斯亮蓝G-250染色、扫描,扫描模式为256灰阶透视扫描,分辨率为200 dpi。
1.3.11 氨基酸含量测定
取0.5 g样品于水解管中,加入20 mL体积分数50% HCl溶液,置于电热鼓风干燥箱110 ℃水解22~24 h。取出冷却,转移至25 mL比色管中定容。从比色管中取1 mL溶液,85 ℃水浴吹干,加蒸馏水1 mL,氮气吹干。加入10 mL 0.02 mol/L HCl溶液,摇匀,取500 μL加入250 μL 0.1 mol/L异硫氰酸苯酯乙腈、250 μL 1 mol/L三乙胺乙腈,衍生1 h后,加入2 mL正己烷,振荡、静置分层。取下层液过0.45 μm有机膜后用氨基酸自动分析仪测定氨基酸含量。
1.4 数据统计与分析
所有实验数据均为3 次重复实验结果的平均值。通过DPS 7.55软件对数据进行单因素方差分析,Duncan新复极差法进行差异显著性分析,P<0.05表示差异显著。采用Origin Pro 8.5软件作图。
2 结果与分析
2.1 氧化时间对羊肉肌原纤维蛋白羰基含量的影响
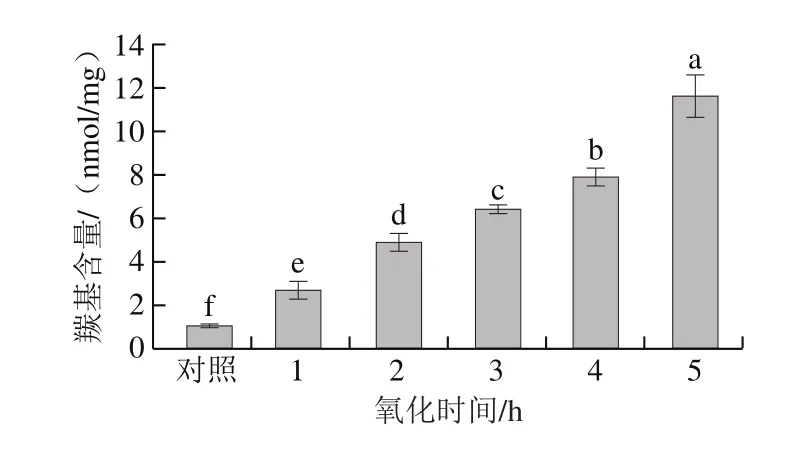
图1 氧化时间对肌原纤维蛋白羰基含量的影响Fig.1 Effect of oxidation time on carbonyl group content
羰基含量是广为使用的衡量蛋白质氧化程度的指标,蛋白质骨架侧链带有的部分氨基酸在氧化时易被转化成羰基基团[20]。由图1可知,随着氧化时间的延长,羰基含量显著增加(P<0.05),氧化5 h的肌原纤维蛋白羰基含量比对照组增加了10.03 倍。这与李学鹏等[21]研究的经羟自由基不同氧化时间处理的六线鱼肌原纤维蛋白羰基含量的结果一致。
2.2 氧化时间对羊肉肌原纤维蛋白总巯基含量的影响
巯基在蛋白质中有两种存在形式:一种存在于蛋白质表面;另一种是包埋在蛋白质内部,总巯基包括以上两种形式[22]。由图2可知,随着氧化时间的延长,总巯基含量显著降低(P<0.05),并在5 h时达到5.09 μmol/g,说明羊肉的肌原纤维蛋白的氧化程度随着氧化时间的延长而呈上升趋势。有报道认为总巯基含量的减少代表二硫键的生成[23]。

图2 氧化时间对肌原纤维蛋白总巯基含量的影响Fig.2 Effect of oxidation time on total sulfhydryl group content
2.3 氧化时间对羊肉肌原纤维蛋白二聚酪氨酸含量的影响
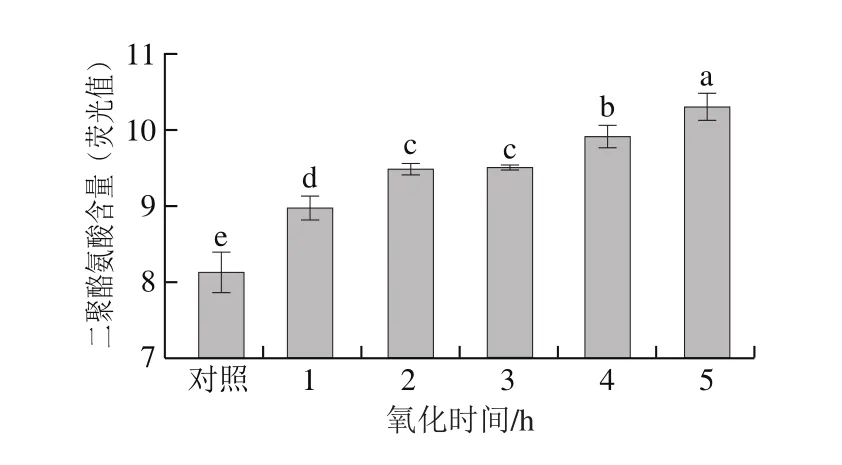
图3 氧化时间对肌原纤维蛋白二聚酪氨酸含量的影响Fig.3 Effect of oxidation time on tyrosine dimer content
二聚酪氨酸含量也是衡量蛋白质是否发生氧化的指标,主要在于氧化的发生使酪氨酸残基发生不同程度的聚合,从而形成二聚酪氨酸[15]。由图3可知,随着氧化时间的延长,二聚酪氨酸含量呈上升趋势;与对照组相比,氧化样品的二聚酪氨酸含量显著增加(P<0.05),而2、3 h处理组之间聚酪氨酸含量差异不显著(P>0.05);4 h后继续上升。蛋白氧化后二聚酪氨酸含量的增加也可能蛋白氧化后通过部分共价键和非共价键的作用发生了聚集[24]。
2.4 氧化时间对羊肉肌原纤维蛋白表面疏水性的影响
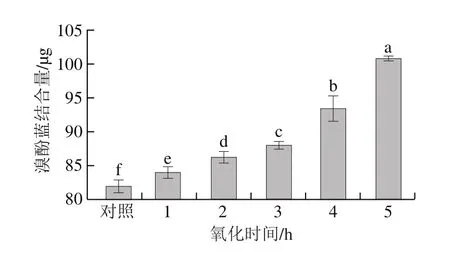
图4 氧化时间对肌原纤维蛋白表面疏水性的影响Fig.4 Effect of oxidation time on protein surface hydrophobicity
蛋白质表面的疏水性结构对其功能性质存在较大作用,一般通过蛋白与溴酚蓝的结合量判断其在氧化过程中的变性程度[21,25]。由图4可知,对照组溴酚蓝的结合量为82.00 μg,氧化时间为1、2、3、4 h和5 h时,结合量分别为84.08、86.38、88.14、93.58 μg和101.22 μg;随着氧化时间的延长,溴酚蓝的结合量呈显著增加趋势(P<0.05),说明氧化使肌原纤维蛋白的表面疏水性显著增强。这可能是由于氧化改变部分疏水性氨基酸结构,使更多疏水性基团暴露于蛋白质表面,从而促进蛋白质与溴酚蓝的结合,使得表面疏水性呈增加趋势[25-26]。
2.5 氧化对羊肉肌原纤维蛋白紫外吸收光谱的影响
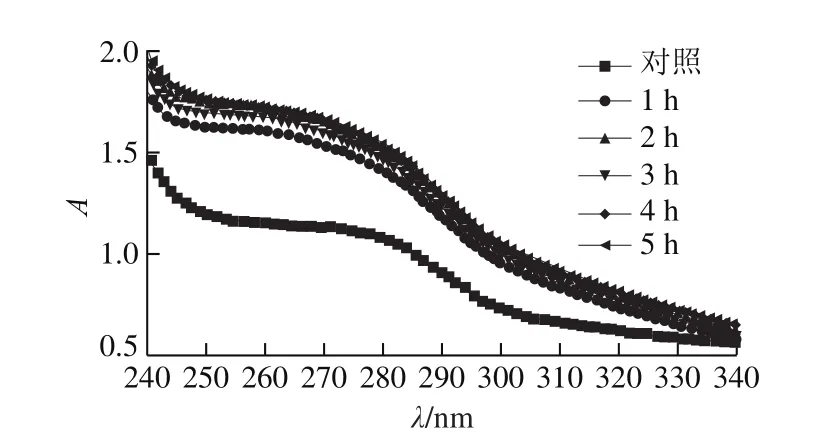
图5 氧化时间对肌原纤维蛋白紫外吸收光谱的影响Fig.5 Effect of oxidation time on ultraviolet and visible absorption spectra of myofibrillar protein
蛋白质中含有芳香环结构的氨基酸,如酪氨酸和色氨酸,能够引起紫外光的吸收,因此紫外吸收光谱常用于反映蛋白质在氧化过程中的结构[27]。由图5可知,经不同时间氧化后的肌原纤维蛋白的紫外吸收光谱相似,均在280 nm波长左右出现峰的拐点,说明氧化不会改变肌原纤维蛋白整体图谱趋势。但随着氧化时间的延长,峰位出现轻微蓝移,可以表明在氧化体系下,不同氧化时间使肌原纤维蛋白质中的部分氨基酸基团结构发生不同程度改变,并且从而使吸收的紫外光谱产生差异,并对蛋白质的功能性质产生影响。
2.6 氧化时间对羊肉肌原纤维蛋白内源荧光光谱的影响
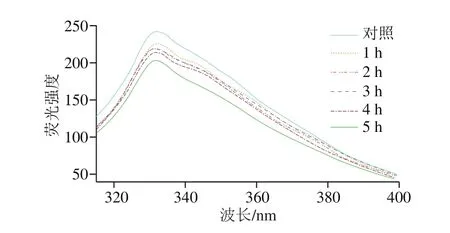
图6 氧化时间对肌原纤维蛋白内源荧光强度的影响Fig.6 Effect of oxidation time on endogenous fluorescence intensity of myofibrillar protein
内源荧光光谱常用来反映蛋白质构象。如图6所示,随着氧化时间的延长,蛋白质的荧光强度呈现降低趋势,且发生轻微蓝移;氧化5 h时荧光强度最低,氧化2、3 h时的荧光强度差异较小。这可能是氧化导致蛋白质分子内部发生能量的转移,部分结构逐渐发生改变;随着氧化时间的延长,部分氨基酸荧光被猝灭,从而导致肌原纤维蛋白的荧光强度呈下降趋势,这与Cao Yungang等[28]发现氧化处理后肌原纤维蛋白荧光强度下降的结论一致。
2.7 氧化时间对羊肉肌原纤维蛋白粒径的影响

表1 氧化时间对羊肉肌原纤维蛋白粒径的影响Table 1 Hffect of oxidation time on protein particle size of myofibrillar protein
粒径反映可溶性蛋白的粒度分布和氧化肌原纤维蛋白的聚集情况[29]。由表1可知,D(v, 0.1)、D(v, 0.5)随着氧化的前4 h呈上升趋势,但5 h时下降;随着氧化时间的延长,D(v, 0.9)呈显著增加趋势(P<0.05)。这可能是较高氧化程度可以促进不可溶性组分形成,导致蛋白分子聚集,使蛋白质粒径增大,而蛋白粒径增大会导致蛋白质持水性降低、蒸煮损失增加等功能特性劣变[30]。Bao Yulong等[31]在猪肉肌原纤维蛋氧化的研究中也发现蛋白聚集。
2.8 氧化时间对羊肉肌原纤维蛋白Zeta电位的影响
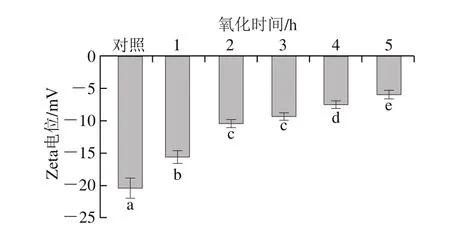
图7 氧化时间对肌原纤维蛋白Zeta电位的影响Fig.7 Effect of oxidation time on protein Zeta potential of myofibrillar protein
Zeta电位指胶态分散体中的电动势,绝对值高意味着肌原纤维蛋白具有更好的稳定性[32]。由图7可知,Zeta电位绝对值随着氧化时间的延长而呈显著降低趋势(P<0.05),表明静电相互作用随着氧化程度的增加而减弱;对照组Zeta电位绝对值为20.53,是氧化5 h样品的3.4 倍。随着氧化时间的延长,Zeta电位绝对值降低,蛋白溶液趋向凝聚状态,相互之间的吸引力大于排斥力,肌原纤维蛋白之间的分散状态被破坏而产生聚集,导致肌原纤维蛋白的稳定性降低[18]。
2.9 氧化时间对羊肉肌原纤维蛋白二级结构的影响
一般认为研究蛋白质的FTIR图是分析蛋白氧化过程中二级结构的变化规律的有效手段,羊肉肌原纤维蛋白在红外区有明显的特征光谱吸收带,在1 600~1 700 cm-1范围的酰胺I带主要以α-螺旋、β-折叠等空间结构相互作用为主,在不同外界因素影响下,蛋白质酰胺I带对肽链结构变化的反映十分准确、灵敏[33-34]。

图8 氧化对肌原纤维蛋白FTIR图谱(A)和二级结构(B)的影响Fig.8 Effect of oxidation time on protein FTIR (A) and secondary structure (B) of myofibrillar protein
由图8A可知,随着氧化时间的延长,羊肉肌原纤维蛋白FTIR图谱趋势相似。由图8B可知,随着氧化时间的延长,与对照组相比,蛋白α-螺旋相对含量呈先增加后降低的趋势,无规卷曲相对含量变化不大,β-折叠相对含量呈降低趋势,β-转角相对含量则先降低后增加。结果表明氧化过程中蛋白质的二级结构会发生不同程度的转变,氧化使得羊肉肌原纤维蛋白的螺旋、折叠结构之间相互转化,且是有序向无序的转变。Sun Weizheng等[30]采用FTIR研究氧化后猪肉肌原纤维蛋白的二级结构变化,发现随着氧化的发生,肌原纤维蛋白结构发生改变,与本实验结果一致。
2.10 肌原纤维蛋白SDS-PAGE分析

图9 不同氧化时间的肌原纤维蛋白SDS-PAGE图Fig.9 SDS-PAGE of myofibrillar protein at different oxidation times
由图9可知,肌原纤维蛋白氧化后结构上的聚集变化,通过氧化后的羊肉肌原纤维蛋白SDS-PAGE分析可知,肌原纤维蛋白的SDS-PAGE图中依次出现分子质量由大到小的肌球蛋白重链、α-肌动蛋白和肌动蛋白、肌球蛋白、肌钙蛋白。蛋白质分子发生降解主要表现为分子质量较高处的条带出现模糊、弱化和扩展,较低分子质量区域则呈现出新的条带或条带颜色加深[35-36]。
随着氧化时间的延长,肌球蛋白重联处条带有轻微扩散,这表明氧化可能形成蛋白聚集体,由此可假设,氧化使羊肉中肌原纤维蛋白结构改变,可能是蛋白分子的共价交联,如通过二硫键、二酪氨酸等形成交联,或分子间通过碳-碳共价交联等,导致氧化后肌原纤维蛋白产生了分子质量较大的聚集体。另外,在100 kDa处条带逐渐加深,35~40 kDa处条带呈现扩展现象,说明氧化对肌原纤维蛋白的不同部分有相同作用。Xia Minquan等[37]对不同氧化程度的猪肉纤维蛋白的SDS-PAGE分析表明,猪肌原纤维蛋白经不同处理后,低水平的氧化出现肌原纤维蛋白聚集体[37],这与本实验结果相类似。
2.11 氧化时间对羊肉肌原纤维蛋白氨基酸含量的影响

表2 氧化时间对肌原纤维蛋白氨基酸含量的影响Table 2 Effect of oxidation time on amino acids contents of myofibrillar protein
蛋白氧化造成侧链氨基酸发生不同程度氧化,从而使氨基酸的种类、比例改变。如表2所示,随着氧化时间的延长,胱氨酸含量显著增加(P<0.05),其他氨基酸含量呈下降趋势,氨基酸总量也逐渐下降,不同氨基酸对氧化时间的响应也不尽相同,这与李银等[38]的研究结果吻合。氨基酸总量减少是由于随着氧化时间的延长,蛋白质侧链的脂肪族氨基酸被氧化形成氢过氧化物、羰基衍生物,芳香族氨基酸残基的芳香环易被氧化,如酪氨酸易被氧化成二酪氨酸;含硫氨基酸如半胱氨酸和蛋氨酸被氧化为二硫化合物、蛋氨酸亚砜化合物。
与对照组相比,氧化5 h时天冬氨酸(Asp)、酪氨酸(Tyr)和蛋氨酸(Met)含量显著降低(P<0.05),谷氨酸(Glu)、苏氨酸(Thr)含量无显著差异(P>0.05),除胱氨酸外的其他氨基酸含量在氧化3 h后显著降低(P<0.05)。胱氨酸含量的增加可能是由于两分子半胱氨酸经氧化后生成胱氨酸。酪氨酸易被氧化成二聚酪氨酸而使酪氨酸含量下降、二聚酪氨酸含量上升,这与二聚酪氨酸含量结果相一致。
3 结 论
随着氧化时间的延长,羊肉肌原纤维蛋白的羰基含量显著增加、总巯基含量显著降低,表明蛋白质氧化程度逐渐增加。随着氧化时间的延长,羊肉肌原纤维蛋白的二聚酪氨酸含量、表面疏水性呈显著上升趋势,内源荧光强度降低并出现轻微蓝移。在氧化过程中,肌原纤维蛋白的粒径分布向粒径大的方向偏移,Zeta电位的绝对值呈现降低趋势。红外光谱显示蛋白质二级结构发生改变,α-螺旋相对含量先增加后降低,β-转角相对含量先降低后增加,β-折叠相对含量呈现降低趋势,无规卷曲相对含量无明显变化。SDS-PAGE结果表明,氧化引起蛋白质分子间的交联现象。总氨基酸含量呈下降趋势,胱氨酸含量随着氧化时间的延长而显著上升,其余氨基酸含量呈降低趋势,但不同氨基酸对氧化时间的响应不同。因此,氧化能诱导羊肉肌原纤维蛋白的分子与理化特性发生改变,进而对羊肉及其制品品质产生不良影响,研究结果可为调控羊肉品质提供理论基础。
