鸡Prnp基因原核表达载体的构建及其在大肠埃希菌中的表达
2020-12-29李仙春毛耀芳杨海峰余海山马永华万学瑞
李仙春,芦 艳,毛耀芳,杨海峰,余海山,马永华,*,万学瑞,*
(1.甘肃农业大学 动物医学院,甘肃 兰州 730070; 2.西北师范大学 国有资产管理处,甘肃 兰州 730070)
朊病毒(prion virus)是一类无免疫性的疏水蛋白质,也被称作蛋白质侵染因子,能够在宿主细胞内复制并且感染动物。朊病毒没有RNA和DNA,不可以自我复制,它的复制方式是:生物体内正常的细胞型PrPC蛋白接触到朊病毒,分子构象发生了改变,从而形成了具有致病性的PrPSC蛋白[1]。在酵母和真菌中,朊病毒是由蛋白质组成的非染色体基因,通常是蛋白质的改变形式,其催化蛋白质的相同变化。因此,酵母朊病毒既可以垂直传播(作为蛋白质组成的基因),也可以水平传播(作为感染性蛋白或朊病毒)。淀粉样蛋白是大多数酵母和真菌朊病毒的基础,朊病毒蛋白是淀粉样蛋白多种构象形式的一种[2]。有学者认为,构象的转变可能是发生在局部变性的吞噬小胞内,由于较低的pH值诱导蛋白构象的转变,促进淀粉样蛋白的自身组装[3-5]。一般而言,pH、温度、离子强度、促溶剂、氧化应激、金属离子会严重影响蛋白的多肽构象[6]。朊病毒病首先在羊身上发现,被称为“羊瘙痒症”,后来又相继发现了牛海绵状脑病、传染性水貂脑软化病、马鹿和鹿的慢性消瘦病、猫的海绵状脑病、克雅病、库鲁病等[7]。朊病毒病传染动物,如牛的传染性海绵状脑病、羊的瘙痒病、麋鹿和鹿的慢性消耗性疾病(chronic wasting disease,CWD),它们被认为是通过消化道传染(bovine spongiform encephalopathy,BSE)或者是通过物种间的水平传播[8]。大量的研究发现,这些疾病的共同特征是人和动物产生意识和运动功能的严重衰退直至死亡,其临床和病理特征表现为脑组织的海绵体化、空泡化、形成星形胶质细胞和小胶质细胞[9]。
朊蛋白(prion protein,PrP)是由朊蛋白基因(prion protein gene,Prnp)编码的,具有高度保守性,不仅在人和动物细胞表面广泛表达,还在免疫系统中也有很高的表达,包括T、B淋巴细胞,树突状细胞,NK细胞等[10]。鸡是最早发现Prnp基因的禽类,最初鸡朊蛋白(ChPrPC)是作为乙酰胆碱受体从鸡神经组织中分离出来的,介导神经轴突的信号传递[11-12]。PrP通常以PrPC和PrPSC这两种形式存在,前者以α-螺旋为主,具有可溶性的细胞型正常朊蛋白,对蛋白酶K敏感;后者为异常蛋白,以β-折叠为主,具有致病性并抑制蛋白酶K[13];甲酸和高温水浴对PrP的暴露具有很好的修饰作用[14]。PrPC作为细胞表面的糖蛋白,主要存在于中枢神经系统中,此外在淋巴细胞、胃上皮细胞、心、肾和肌肉等许多非神经组织中也有表达,已有研究表明,朊蛋白不仅存在于哺乳动物体内,而且在禽类、两栖类动物、鱼类、酵母中均有表达[15-16]。PrPC也可通过促进细胞增殖和抑制凋亡而在口腔癌中发挥重要的作用[17]。
朊蛋白是由第20号染色体短臂上的朊蛋白基因编码,由253个氨基酸组成。N端除了信号肽负责蛋白外分泌,还有5个八肽重复序列(octarepeat),可结合一定数量的铜离子,与朊蛋白构象转变密切相关[18]。Prnp基因突变会造成朊蛋白构象的改变,进而影响它的稳定性,缺失则会导致相关基因的调控系统发生紊乱。PrPC在发育上是受调控的,PrPC在未成熟的大脑组织中高度表达可能与它能够调控神经形成和细胞增殖有关。最近的一项研究表明,PrPC在调节海马体中的蛋白质激酶 A(PKA)突触可塑性方面起着至关重要的作用。
PrP尽管在哺乳动物和禽类中有很多研究,但其功能还不是很明确[19]。虽然,不同物种朊蛋白的一级结构有所不同,但还存在保守结构域。对青蛙、乌龟、鸡、哺乳动物的朊蛋白进行核磁共振研究分析发现,它们都存在类似的球形结构域,C-端球形结构域由3个α-螺旋、一个短的β-螺旋和一个短的反向平行β-折叠组成,N-端是无规则的可变区;而已知哺乳动物和禽类的朊病毒同源性很低,这种重要的结构域和肽段是保守的,禽类没有自发感染的病例[20-21]。多年来,人们对朊蛋白的结构、生理功能和致病机制进行了大量研究,PrPC最重要的功能是参与细胞信号转导,由于PrPC在细胞膜外定位,PrPC能够将环境中的分子信号传递到细胞内;朊蛋白还参与了多种生理过程,如应激保护、细胞分化、神经元兴奋性、髓鞘维持、昼夜节律和金属离子稳态等[21-22]。朊蛋白在肿瘤的形成中具有一定作用,不仅干扰肿瘤形成的通路,还调控肿瘤细胞的分化与凋亡。朊蛋白的多态性影响了朊病毒的敏感性和种间屏障[23]。本实验拟构建鸡Prnp基因原核表达体系,并进行初步分析,为制备鸡朊蛋白单克隆抗体提供材料。
1 材料与方法
1.1 材料
1.1.1 质粒和菌株
表达载体pET-28a;E.coliTop10、E.coliBL21(DE3)保藏于甘肃农业大学微生物实验室。
1.1.2 酶和试剂
GreenTaqMix DNA聚合酶,T4 DNA连接酶,10×K Buffer,10×M Buffer,DNA凝胶回收试剂盒(EasyPure GeL DNA Extration Mini kit)均购自南京诺唯赞(Vazyme)生物科技有限公司;酶BamH I和Hind Ⅲ、GeLRed核酸染料、Loading Buffer、0.1%BSA(牛血清白蛋白)均购自TaKaRa;卡那青霉素(Kan)、提质粒试剂盒、纯化试剂盒均购于全式金生物技术有限公司;DL2000、DL5000 DNA Marker、彩虹180广谱蛋白Marker(Solarbio)、异丙基-β-D-硫代半乳糖苷(IPTG)、三羟甲基氨基甲烷(Tris)、十二烷基硫酸钠(SDS)均购于BBI公司。
1.1.3 主要仪器、设备
恒温培养振荡器ZHWY-200D(ZHICHNG);电热恒温培养箱HG202-3K(南京电气三厂);TG16-WS台式高速离心机(Cence湘仪);梯度基因扩增仪(Eppendorf,德国);Centrifuge 545小型高速离心机(eppendorf,德国);PCR扩增仪(美国伯乐BIO-RAD);WTL-6K迷你离心机(Cence湘仪);超净工作台(海尔生物医疗);紫外、可见分光光度计(GE Healthcare,英国);恒温金属浴MK-20(AllSHENG);凝胶成像分析仪(BIO-RAD);超低温冰箱(-70、-80 ℃);台式高速冷冻离心机(Cence)。
1.2 方法
1.2.1 引物设计与合成
根据GenBank已报道的鸡Prnp基因cDNA及基因组序列和原核表达载体pET-28a质粒图谱设计了如下1对引物,由金唯智(天津)生物科技有限公司合成。
上游引物F:5′-CGGGATCCATGGCTAGGCTCCTCACCACC-3′;
下游引物R:5′-CCCAAGCTTGTGCATGGCAAAAAGGGTGG-3′。
上述合成引物中下划线部分为引入的BamH I和Hind Ⅲ酶切位点。
1.2.2 鸡血液中基因组的提取
采取健康三黄鸡新鲜血液于抗凝管中,并根据全血提取基因组试剂盒的操作步骤进行基因组的提取,总DNA由1%琼脂糖凝胶电泳检测。
1.2.3 鸡Prnp目的DNA片段的PCR扩增
采用50 μL体系:上、下游引物各2.0 μL,TaqDNA聚合酶25 μL,模板2.0 μL,加灭菌蒸馏水至50 μL;离心混匀后至PCR扩增仪中,94 ℃预变性5 min,94 ℃ 30 s、51.1 ℃ 30 s和72 ℃ 2 min,共35个循环,72 ℃再延伸5 min,最后4 ℃储存。PCR产物经1%的琼脂糖凝胶电泳鉴定后,将符合预期大小的目的条带切下,按DNA凝胶回收试剂盒的说明回收其目的DNA片段。
1.2.4 目的基因与pET-28a质粒的酶切
将回收的目的DNA片段和pET-28a质粒DNA分别用限制性内切酶BamH Ⅰ和Hind Ⅲ进行双酶切。第一次酶切体系(50 μL):目的DNA片段/质粒30 μL,10×K Buffer 5.0 μL,BamH Ⅰ 3.0 μL,加灭菌蒸馏水至50 μL;37 ℃水浴3 h,酶切产物经1%琼脂糖凝胶电泳验证。第二次酶切体系(50 μL):第一次酶切产物3.0 μL,10×M Buffer 5.0 μL,BSA 5.0 μL,Hind Ⅲ 3.0 μL,加灭菌蒸馏水至50 μL;37 ℃水浴3 h,酶切产物经1%琼脂糖凝胶电泳验证后,切下目的条带,用DNA凝胶回收试剂盒纯化回收酶切目的片段和质粒片段。
1.2.5 重组表达载体的构建与鉴定
将双酶酶切的鸡Prnp目的DNA片段和pET-28a质粒DNA片段进行连接,转化至E.coliTop10感受态细胞中进行克隆,通过BamH Ⅰ和Hind Ⅲ双酶切鉴定并测序,挑选出阳性的重组表达菌。
表达载体片段和目的DNA片段的连接与转化。(1)感受态细胞的制备。①分别从37 ℃过夜培养的新鲜培养基上挑取E.coliTop10和E.coliBL21(DE3)单菌落,接种于普通的LB液体培养基(3 mL)中;②将过夜培养的细菌扩增至100 mL的LB液体培养基(1∶100)中,37 ℃ 250 r·min-1,振荡培养3 h至D600为0.45~0.60;③在超净台下,将细菌培养物转移至2个冰冷的50 mL离心管中,冰浴10 min,然后将离心管放4 ℃ 5 000×g离心10 min;④弃上清,将冰冷的100 mmol·L-1CaCl2溶液在离心管中加入5 mL,混匀后再加入20 mL CaCL2溶液重悬(两管合一),4 ℃ 5 000×g离心10 min;⑤弃上清,将冰冷的100 mmol·L-1CaCl2溶液用25 mL吹打混匀管底的菌体,冰浴45 min,放4 ℃ 5 000×g离心10 min;⑥弃上清,用5 mL冰冷的含15%甘油的100 mmol·L-1CaCl2溶液混匀菌体,每100 μL一管分装在冰冷的1.5 mL无菌离心管中,置于-70 ℃冰箱冻存。(2)目的DNA片段与表达载体片段的连接。将双酶切后纯化回收的Prnp目的DNA片段和载体pET-28a质粒 DNA片段于16 ℃连接8 h后,4 ℃冰箱保存,次日进行转化。连接体系如表1所示。(3)重组质粒的转化与培养。将3个不同连接体系的连接产物转化至E.coliTop10感受态细胞中,接种于不含有卡那青霉素(Kan)的890 μL LB培养液的无菌EP管中,37 ℃ 220 r·min-1振荡培养1 h,再将EP管5 000×g离心2 min,弃上清液后用枪头将剩余的细胞沉淀和培养液轻轻混匀,分别打到LB固体培养基(含Kan)上,用L棒涂抹均匀,37 ℃恒温箱过夜培养。
重组表达质粒的鉴定:(1)PCR鉴定。用灭菌接种环分别从3个平皿中挑选单个菌落,接种到3 mL含Kan的LB液体培养基的试管中,37 ℃ 220 r·min-1振荡过夜,3个试管分别做菌液PCR扩增,用1%琼脂糖凝胶电泳观察结果。菌液PCR体系(15 μL):上、下游引物各0.6 μL,TaqDNA聚合酶7.5 μL,菌液1.8 μL,加灭菌蒸馏水至15 μL。(2)双酶切鉴定。根据试剂盒说明从菌液中提取重组质粒,经BamH Ⅰ和Hind Ⅲ进行双酶切。双酶切体系(20 μL):重组质粒 15 μL,1×K Buffer 1.0 μL,酶BamH Ⅰ 1.0 μL,酶Hind Ⅲ 1.0 μL,加灭菌蒸馏水至20 μL。

表1 连接体系
重组表达质粒的序列测定:将PCR扩增和双酶切鉴定均为阳性克隆的重组质粒,各取15 μL送金唯智(天津)生物科技有限公司进行测序。
1.2.6 重组表达质粒的诱导表达
转化:经DNAstar软件比对序列,将测序结果正确的重组质粒pET-28a-ChPrnp转化至E.coliBL21(DE3)感受态细胞。
Prnp蛋白的诱导表达:从转化后的重组菌株pET-28a-ChPrnp(BL21)中挑取单菌落接种于4 mL的LB液体培养基中,37 ℃ 220 r·min-1过夜培养,取1 mL菌液于-20 ℃保存(未诱导的菌液)。加入0.10 mmol·L-1的IPTG诱导剂分别在16、28、37 ℃下诱导12 h,确定最佳诱导温度为16 ℃;然后分别将IPTG诱导剂的终浓度为0.08、0.09、0.10、0.11、0.12 mmol·L-1的菌液,16 ℃ 220 r·min-1诱导6 h,确定诱导剂的最佳浓度;在已确定最佳诱导浓度时,16 ℃ 220 r·min-1诱导5和7 h,以确定最佳诱导时间。
处理样品:将未诱导的菌液和诱导的菌液各取50 μL,再加入50 μL的上样缓冲液混匀煮沸10 min,冷却至室温后12 000×g离心5 min,取上清进行电泳。
SDS-PAGE:配置好蛋白胶(制胶配方见表2),然后分别取处理好的样品各40 μL加入样品孔中,先用80 V电压电泳,观察样品电泳到分离胶时,再将电压调至120 V,观察样品电泳至分离胶边缘时,关闭电源。
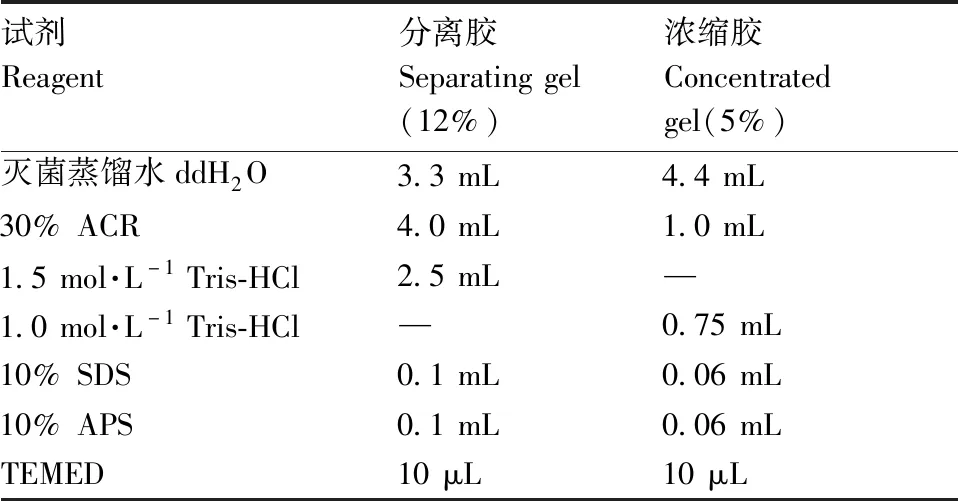
表2 蛋白胶配方
染色及脱色:电泳完毕,将蛋白胶从玻璃板上小心取下,用R-250考马斯亮蓝染色液在脱色摇床上染色3 h后,弃去染色液加入脱色液进行脱色,期间更换脱色液3~4次,直到出现清晰的条带,将凝胶放入蒸馏水中终止脱色。
2 结果与分析
2.1 鸡Prnp基因目的DNA片段的获得
以健康三黄鸡的全血总DNA作为PCR扩增的模板,同Prnp基因的上游引物和下游引物,及其耐高温的Taq酶进行扩增,经1%琼脂糖凝胶电泳检测,得到了约为822 bp的条带(图1),与预期的目的DNA相对分子质量大小相符。
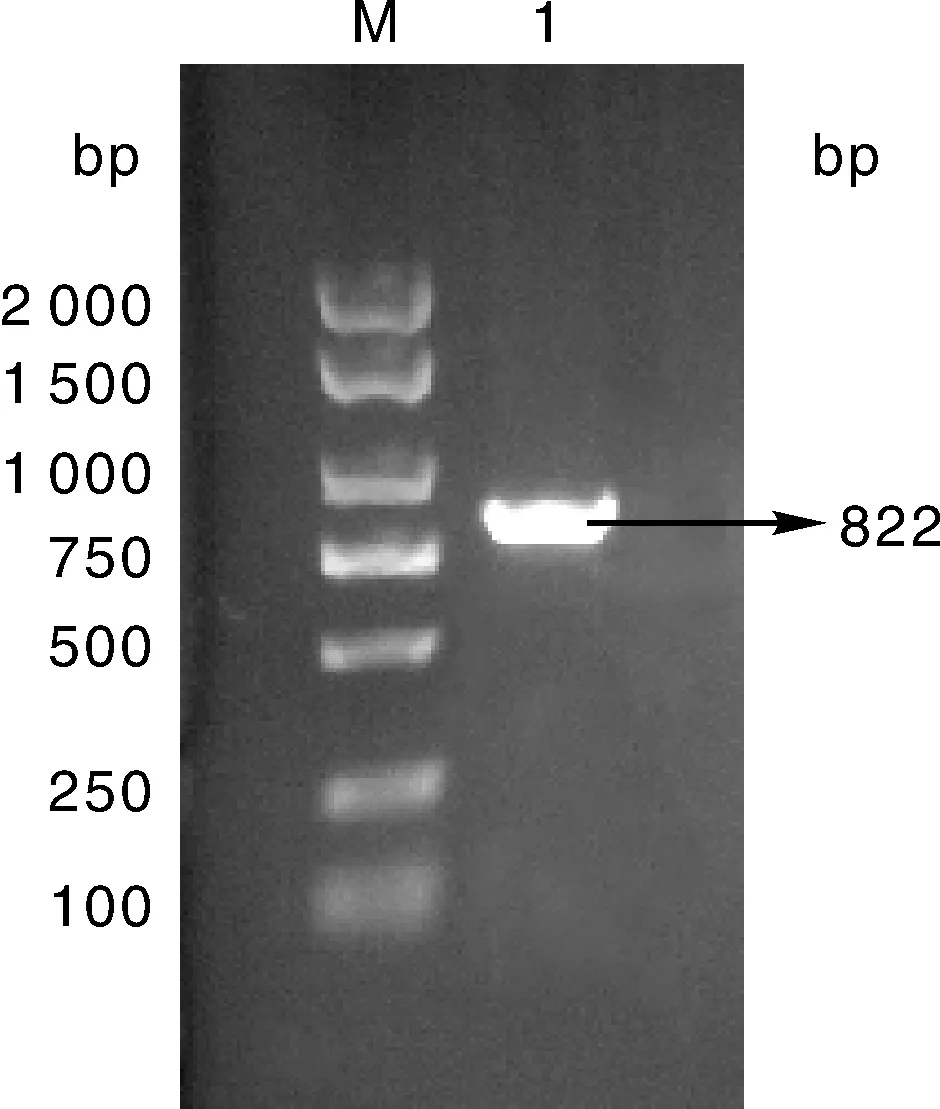
M,DL2000 DNA marker;1,PCR扩增产物ChPrnp(822)。M, DL2000 DNA marker;1, PCR amplification product of ChPrnp(822).图1 目的基因PCR扩增产物Fig.1 PCR amplification product of target gene
2.2 目的DNA与pET-28a质粒的酶切分析
双酶切得到目的基因片段(822 bp)和质粒DNA片段(5 369 bp),形成可以互补配对的粘性末端,为重组质粒的连接提供了条件(图2)。
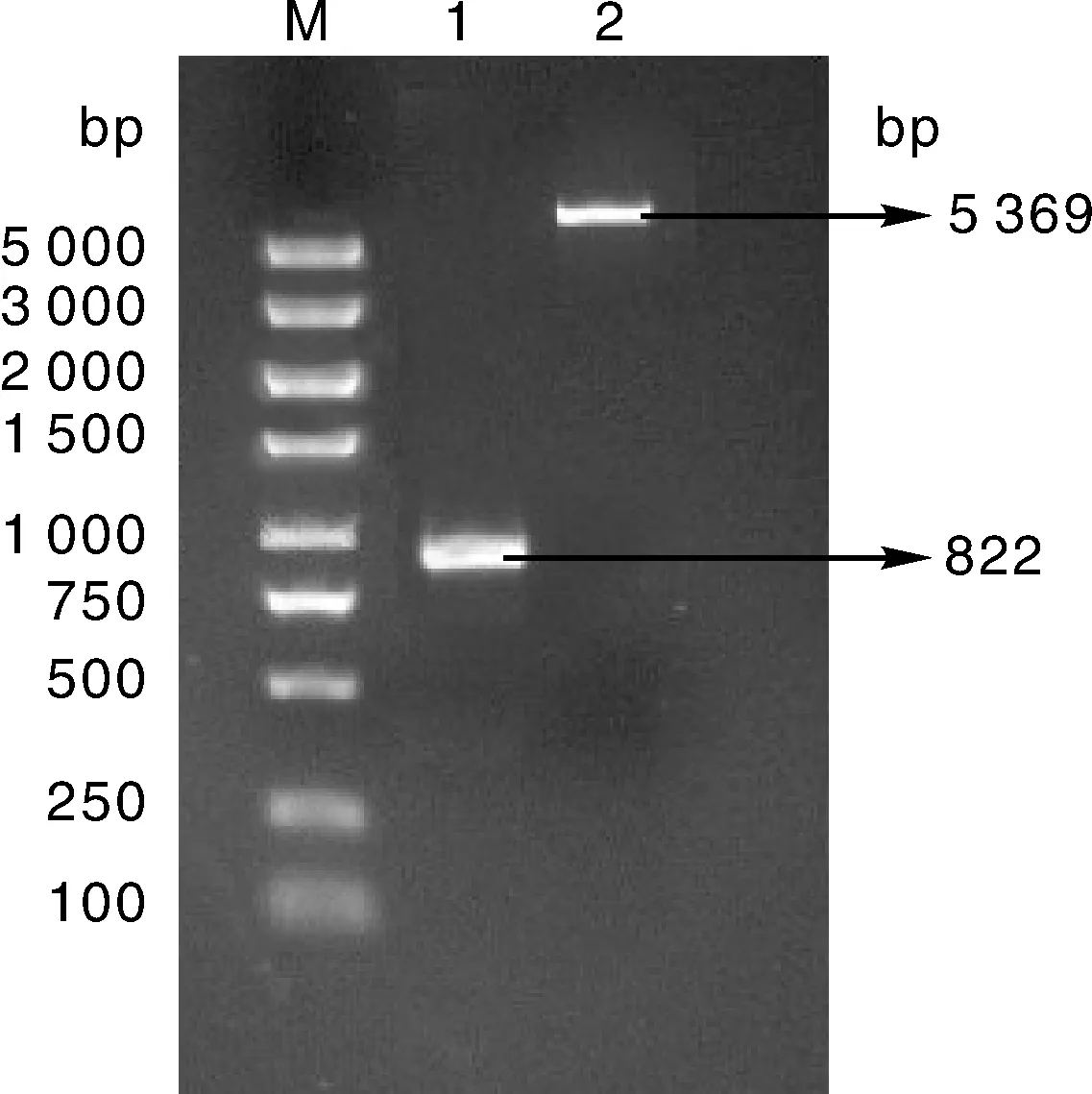
M,DL5000 DNA marker;1,目的DNA酶切产物;2,pET-28a质粒酶切产物。M, DL5000 DNA marker; 1, Digested product of target DNA; 2, Digested product of pET-28a plasmid.图2 目的DNA与pET-28a质粒的酶切产物Fig.2 Digested product of target DNA and pET-28a plasmid
2.3 重组表达质粒的菌液PCR鉴定
菌液PCR扩增获得目的DNA片段(图3),证明在不同连接体系下,目的DNA片段与载体片段都可以成功连接。
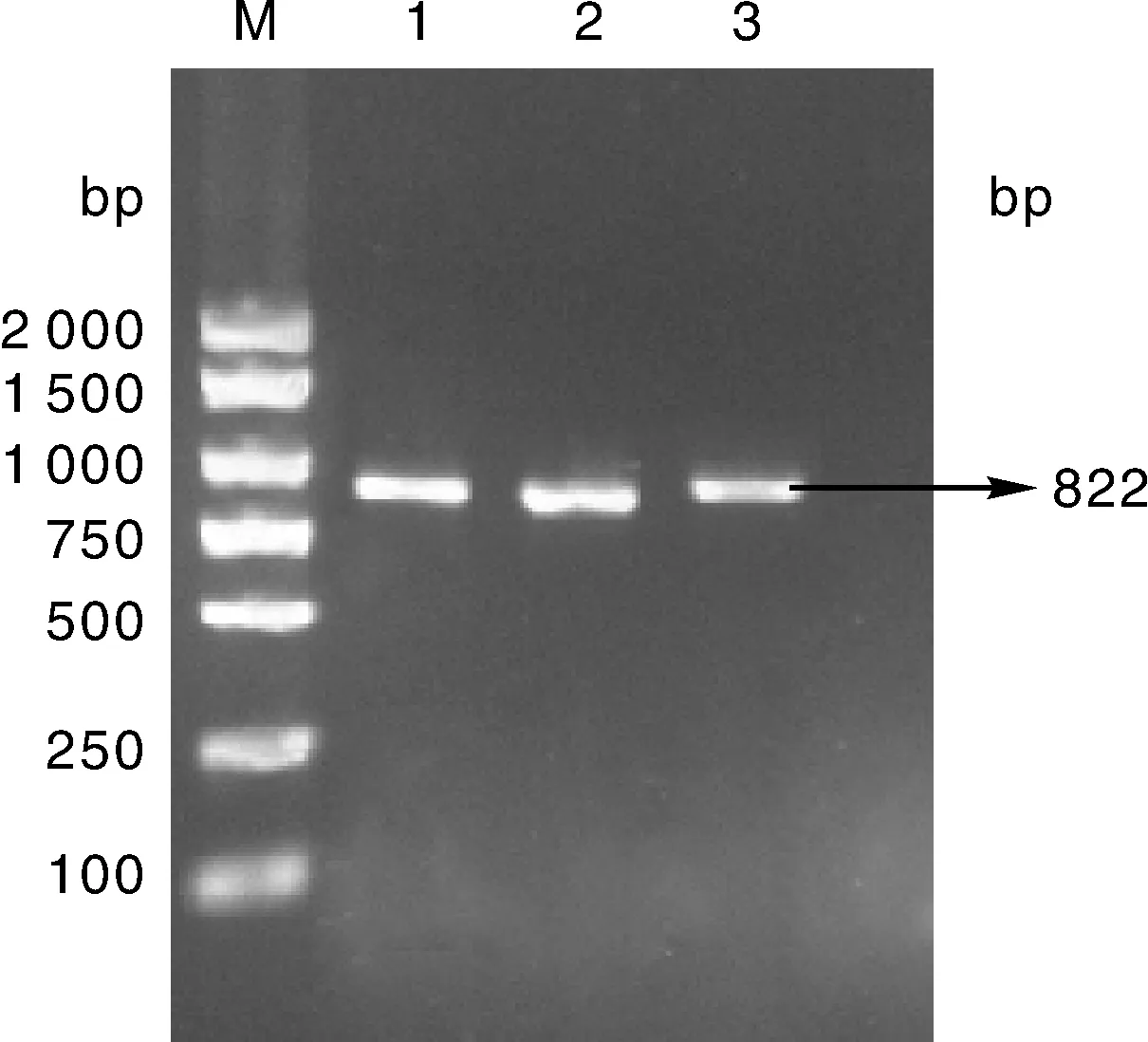
M,DL2000 DNA marker;1~3,1、2、3号菌液PCR扩增产物。M, DL2000 DNA marker; 1-3, PCR amplification products of bacteria solution 1, 2 and 3.图3 重组表达质粒的菌液PCR鉴定Fig.3 PCR identification of recombinant plasmid
2.4 重组表达质粒的双酶切鉴定
双酶切得到大约5 369 bp和822 bp的两条片段(图4),证明重组表达载体中含有预期大小的目的DNA片段,初步证明构建了重组表达载体。
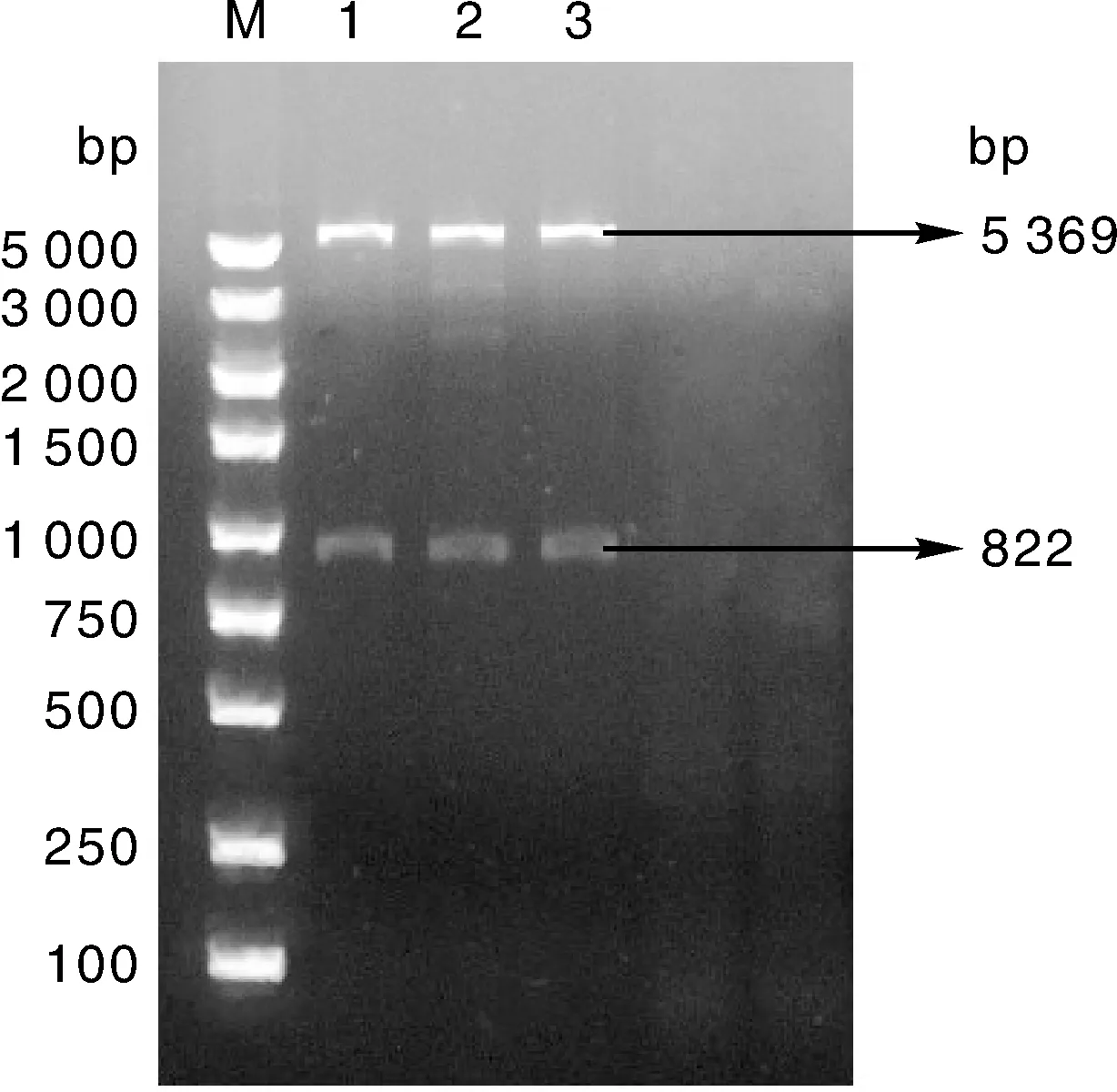
M,DL5000 DNA marker;1~3,1、2、3号重组质粒双酶切产物。M, DL5000 DNA marker; 1-3, Double enzyme digestion products of recombinant plasmids 1, 2 and 3.图4 重组表达载体双酶切分析Fig.4 Double enzyme digestion analysis of recombinant expression vector
2.5 重组表达载体的序列分析
将PCR扩增和双酶切鉴定均为阳性克隆的3管不同连接体系的重组质粒,分别取15 μL送金唯智(天津)生物科技有限公司测序,DNAstar软件进行序列比对,1号管的基因序列方向正确,提示1号管提取的重组表达质粒符合要求,插入了相对分子质量大小约为822 bp的目的DNA片段,证明成功构建了重组表达载体,将其命名为pET-28a-ChPrnp(822)。
2.6 重组菌表达产物的SDS-PAGE分析
经表达,IPTG诱导剂在16 ℃下诱导蛋白表达量最高(图5),加入IPTG的终浓度为0.08 mmol·L-1
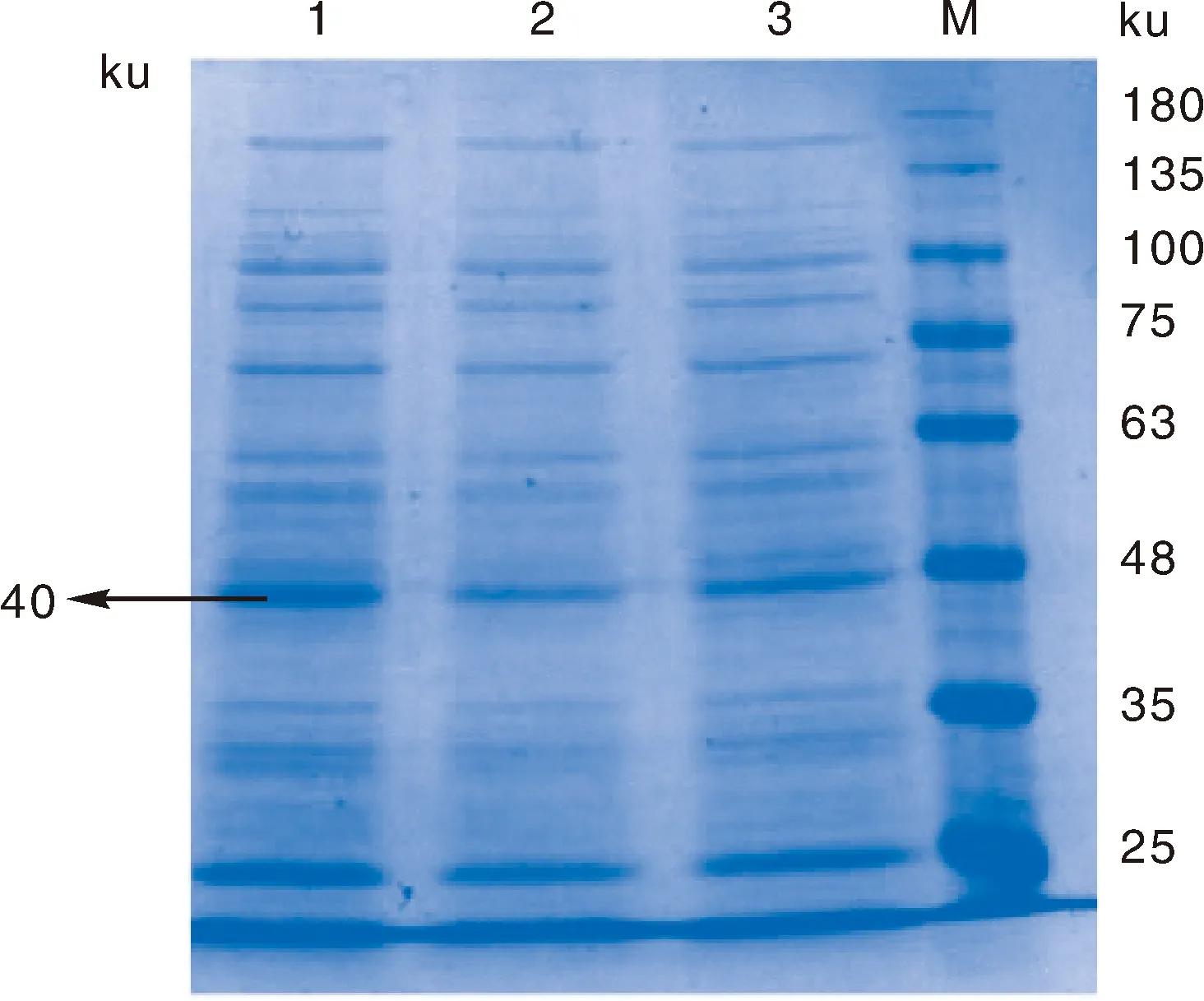
1~3,16、28、37 ℃的温度下诱导表达的蛋白产物;M,蛋白质相对分子质量标准。1-3, Protein products induced at 16, 28 and 37 ℃; M, Protein marker.图5 重组菌诱导表达IPTG温度优化Fig.5 Temperature optimization of induced expression of IPTG by recombinant bacteria
进行诱导时,蛋白表达量最高(图6),在已确定诱导剂最佳浓度时,16 ℃ 220 r·min-1诱导表达7 h,为正式表达优化条件(图7)。SDS-PAGE电泳分析其蛋白表达量,发现相对分子质量大约为40 ku的一条外源蛋白表达条带,表达的蛋白量大小和预测的Prnp蛋白分子量相一致。
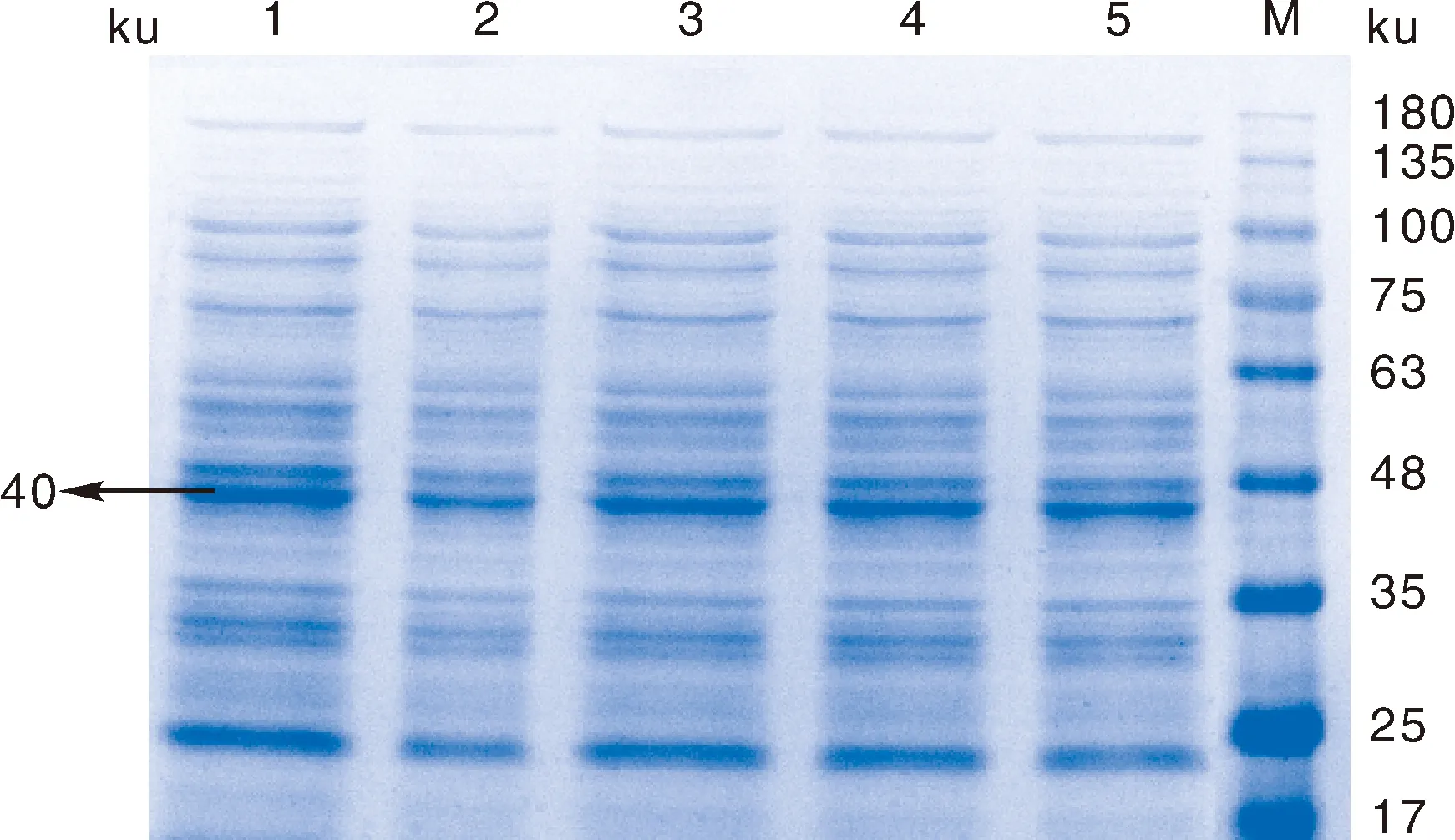
1~5,分别加入IPTG的终浓度为0.08、0.09、0.10、0.11、0.12 mmol·L-1;M,蛋白质相对分子质量标准。1-5, Adding 0.08, 0.09, 0.10, 0.11, 0.12 mmol·L-1 IPTG, respectively; M, Protein marker.图6 重组菌诱导表达IPTG浓度优化Fig.6 Optimization of concentration of IPTG induced by recombinant bacteria
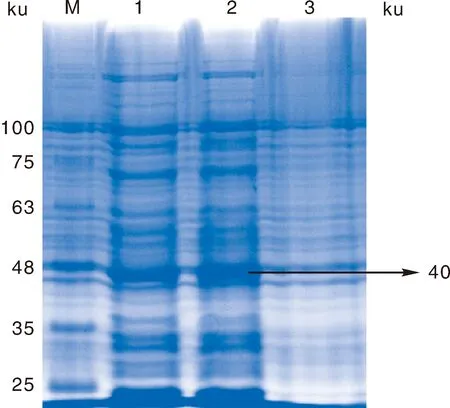
M,蛋白质相对分子质量标准;1~2,诱导5、7 h的重组菌表达产物;3,未诱导的重组菌。M, Protein marker; 1-2, Expression products of recombinant bacteria induced for 5 and 7 h; 3, Uninduced recombinant bacteria.图7 重组菌诱导表达时间优化Fig.7 Optimization of expression time of IPTG induced by recombinant bacteria
3 讨论
一般来说,异源表达更容易获得特定的蛋白,它通过分子生物学技术将外源基因克隆到人工载体上,构建出异源表达体系,分析其结构和特性。大肠埃希菌遗传背景清楚、易培养、生长廉价、繁殖速度快、蛋白高表达,因此它是很多异源蛋白表达的首选。本实验的表达宿主菌是不能修饰表达产物的BL21,因此重组表达产物没有糖基化[24]。
传染性海绵状脑病(TSEs)也称朊病毒病,在脑内淀粉样沉积,引起中枢神经病变,导致人和许多动物出现致死性神经退行性疾病,Prnp是调控TSEs的主要基因,基因结构简单,大多含有3个或5个外显子,无内含子[24-25]。Prnp基因在神经元中的表达量很高,并且具有高度保守性,为朊病毒跨物种传播提供了分子依据[26]。
目前,朊病毒在牦牛、驼峰、水牛、奶牛、山羊、绵羊、梅花鹿等哺乳动物中已有多方面研究[24,26-27]。在已研究的哺乳动物中,Prnp基因ORF中的DNA序列和PrP氨基酸序列的相似性分别达到90%和95%以上[27]。鸡PrP氨基酸序列的相似性同哺乳动物相比只有30%[24]。水牛朊蛋白基因氨基酸序列同牛科动物相比同源性在97.1%以上,不同品种鹿之间朊蛋白基因氨基酸序列之间的同源性较高[28]。牛与绵羊间Prnp基因与其氨基酸序列的同源性分别为97.3%和96.5%;牛与人间Prnp基因与其氨基酸序列的同源性分别为86.7%和90.3%;人与绵羊间Prnp基因与其氨基酸序列的同源性分别为87.1%和90.7%[29]。Prnp基因很大程度上影响了山羊对瘙痒病的易感性[30]。Prnp基因同源性的高低在一定程度上也影响朊病毒种间屏障的形成,但还需要其他辅助因子共同作用[12]。
国内外研究朊蛋白主要从它的基因构型、蛋白结构、生理功能、致病机制方面展开,为朊病毒病的预防和治疗提供基础。本研究用pET-28a质粒作为表达Prnp基因的原核表达载体,其相对分子质量较小,N端带有一个His/凝血酶/T7标签蛋白,而在C端带有His标签蛋白,在蛋白诱导表达时,蛋白相对分子质量可能会比实际大10 ku左右,因此最终外源蛋白表达量为40 ku。
利用IPTG诱导重组蛋白表达时,为使蛋白表达量较高,对诱导条件进行了多方面的探索。37 ℃是菌体生长的适宜温度,菌体蛋白本身也需要生长,在此温度下它本身的蛋白急需表达,没有多余的能力去表达外源蛋白,因此诱导表达需要降低其温度,实验表明16 ℃下蛋白表达量较高。而诱导表达时的诱导剂浓度和诱导时间也是影响蛋白表达量的重要因素,经过比对分析,发现在IPTG诱导剂浓度为0.08 mmol·L-1时诱导7 h蛋白表达量较高。用SDS-PAGE电泳验证可以得到与预期大小相符的蛋白条带,初步表达了重组的蛋白。
4 小结
本研究利用pET-28a质粒成功构建了鸡Prnp基因的原核表达载体,并在大肠埃希菌BL21(DE3)中初步表达重组的蛋白,为Prnp朊蛋白的结构、生理功能和致病机制研究提供方法。
