淡翅小花蝽对黄胸蓟马的捕食功能反应与搜寻效应
2020-12-28邱海燕付步礼马晓彤
邱海燕 付步礼 马晓彤


摘要 为评价淡翅小花蝽Orius tantillus对黄胸蓟马Thrips hawaiiensis的捕食作用及控害潜能,在室内条件下研究了淡翅小花蝽对黄胸蓟马2龄若虫的捕食功能反应、搜寻效应和干扰反应。结果表明:淡翅小花蝽对黄胸蓟马若虫的捕食功能反应符合HollingⅡ型,其捕食量随着猎物密度的增加而提高,其搜寻效应随着猎物密度的增加而下降。淡翅小花蝽成虫具有较强的种内干扰作用,随着自身密度的增加,其平均捕食量减少,捕食作用率降低。根据Beddington模型E=a/[1+aThN+bTw(P-1)]拟合获得淡翅小花蝽成虫捕食黄胸蓟马2龄若虫的干扰模型E=0.968 5/(0.011 0N+1.383 7P-0.383 7),表明淡翅小花蝽的搜寻效应随其自身密度和黄胸蓟马密度的增大而下降。研究结果表明淡翅小花蝽对黄胸蓟马具有较大防治潜力。
关键词 淡翅小花蝽; 黄胸蓟马; 功能反应; 搜寻效应; 干扰反应
中图分类号: S 476.2
文献标识码: A
DOI: 10.16688/j.zwbh.2019478
Abstract In order to evaluate the predatory function and potential management capacity of Orius tantillus against Thrips hawaiiensis, the functional response, searching efficiency and interference response of O.tantillus against T.hawaiiensis were studied under laboratory conditions. The predation of T.hawaiiensis by O.tantillus fitted to the Holling-type Ⅱ functional response. The predation rate was positively correlated with the prey density, while the searching efficiency was negatively correlated with the prey density. The mean predation rate decreased with increasing predator density, suggesting that there was a strong intraspecific interference in O.tantillus. The relationships between the searching efficiency (E), O.tantillus density (P) and T.hawaiiensis density (N) could be described by the Beddington model E=a/[1+aThN+bTw(P-1)]: E=0.968 5/(0.011 0N+1.383 7P-0.383 7), suggesting that the searching efficiency declined when the densities of O.tantillus and T.hawaiiensis increased. This study indicates that O.tantillus has the potential as a biological control agent against T.hawaiiensis.
Key words Orius tantillus; Thrips hawaiiensis; functional response; searching efficiency; intraspecific interference
黄胸蓟马Thrips hawaiiensis (Morgan)隶属于缨翅目锯尾亚目蓟马科,又名香蕉花蓟马、夏威夷蓟马[1-2],主要分布在亚洲的热带及亚热带地区,环太平洋地区、北美南部和欧洲一些国家[3],在我国主要分布在海南、广东、广西、云南、福建和台湾等省区,为香蕉、芒果、花卉等作物的重要花期害虫[4-6]。一直以来,黄胸蓟马的防治以化学防治为主,但由于该虫具隐蔽性,且繁殖能力强、生长发育历期短,化学防治较难获得理想的防治效果,而长期不合理地使用化学药剂导致其对很多杀虫剂的敏感性下降,甚至对部分杀虫剂产生了抗性[7-9]。化学药剂的大量使用也造成生态环境的破坏和农产品农药残留问题日益突出。为了响应国家绿色农业的发展要求,害虫的生物防治是解决这些问题的关键,因此有必要开展黄胸蓟马的生物防治研究。
小花蝽是一类对蓟马具有较大防治潜能的天敌昆虫,目前已在北美、澳洲、欧洲的一些国家以及日本成功应用于蓟马防治[10]。随着蓟马类害虫危害日益严重,抗药性问题愈来愈突出,近年来蓟马防治的研究热点转向了如何合理利用当地天敌控制蓟马,并开展了大量的基础研究工作。然而,目前的研究主要集中在东亚小花蝽Orius sauteri、南方小花蝽O. strigicollis、微小花蝽O. minutus对西花蓟马、棕榈蓟马、花蓟马捕食功能和控害潜能的研究[11-15]。而针对我国南部地区的重要蓟马类害虫——黄胸蓟马的生物防治研究还比较缺乏,目前仅付步礼等[16]开展了东亚小花蝽对黄胸蓟马的捕食功能研究,结果表明东亚小花蝽对黄胸蓟马的捕食能力较强。然而东亚小花蝽主要分布在我国的中部和北部,而在海南、广西、云南等地区暂无分布的报道[17-18]。淡翅小花蝽O.tantillus (Motschulsky)主要分布在海南、廣东、广西、福建、云南、四川、贵州等地,为棕榈蓟马、黄胸蓟马、花蓟马、玉米螟等重要害虫的捕食性天敌[19]。但关于淡翅小花蝽对害虫的控害潜能的研究较少,目前仅Mituda等[20]与Navasero等[21]分别开展了淡翅小花蝽对棕榈蓟马和亚洲玉米螟捕食功能的研究,表明其对棕榈蓟马和亚洲玉米螟均具有较强的捕食作用。鉴于淡翅小花蝽在36℃高温下仍可完成发育,较其他一些小花蝽的热敏感性差[22],且其分布区域覆盖了黄胸蓟马的大部分分布区域,因此开展淡翅小花蝽对黄胸蓟马的捕食功能研究,可为正确评价淡翅小花蝽对黄胸蓟马的防治潜能提供理论依据,为当地天敌的开发、利用奠定基础。
1 材料与方法
1.1 材料
供试昆虫:黄胸蓟马采自海南省澄迈县一香蕉园,在室内以新鲜香蕉花进行继代饲养,并挑取1日龄的2龄若虫作为供试猎物。淡翅小花蝽采自海南省儋州市宝岛新村试验场,在温度(26±1)℃、相对湿度(70±5)%,光周期L∥D=16 h∥8 h的人工气候箱内以豆大蓟马进行继代饲养,待其发育至1龄、2龄、3龄、4龄、5龄和成虫时,单头移入指形管中,饥饿24 h作为供试天敌。
儀器:PYX-400Q-A型人工气候箱,韶关市科力实验仪器有限公司。
1.2 试验方法
1.2.1 淡翅小花蝽对黄胸蓟马2龄若虫的捕食功能反应
分别将10、20、30、40、50头黄胸蓟马2龄若虫和1头饥饿24 h的淡翅小花蝽各龄期若虫与成虫放入柱形玻璃管(高80 mm,直径20 mm,下同)内,每管放入1片香蕉花萼片,玻璃管用保鲜膜封口并用0号昆虫针插20个通气孔,每处理重复6次,于24 h后检查记录淡翅小花蝽捕食的黄胸蓟马数。
1.2.2 淡翅小花蝽自身密度对黄胸蓟马搜寻效应的影响
分别将1、2、3、4、5头淡翅小花蝽成虫和120头黄胸蓟马2龄若虫组合放入柱形玻璃管,其他处理同1.2.1。
1.2.3 干扰效应对淡翅小花蝽捕食黄胸蓟马的影响
黄胸蓟马2龄若虫与饥饿淡翅小花蝽成虫以20∶1、40∶2、60∶3、80∶4、100∶5比例组合放入柱形玻璃管中,其他处理同1.2.1。
1.3 数据处理及统计分析
1.3.1 捕食功能反应
捕食功能反应采用HollingⅡ型公式[23]Na=a′TN/(1+a′ThN),其中,Na为被捕食猎物数,N为猎物密度,T为试验持续时间,本试验持续时间为T=1 d(24 h),a′为瞬时攻击率,Th为处置1头猎物所花费的时间。
1.3.2 搜寻效应估计[24]
搜寻效应方程为S=a′/(1+a′ThN),S为搜寻效应,其余同上。
1.3.3 干扰反应[23, 25-26]
干扰反应方程为E=QP-m=Na/(N×P)=a/[1+aThN+bTw(P-1)],其中,E为捕食作用率,Q为搜寻常数,P为捕食者密度,m为干扰常数,a为干扰下的攻击率,Th为处理时间(可从Holling圆盘方程中求得),b为捕食者之间的相遇率,Tw为每个捕食者一次相遇消耗的时间,bTw可作为一个整体,其余同上。
1.3.4 分摊竞争强度[27]
分摊竞争强度为I=(E1-EP)/E1,式中I为分摊竞争强度,E1、EP分别表示1头和P头天敌的捕食作用率。
1.3.5 数据处理
所有数据采用Excel 2010及SPSS 21.0进行统计分析,并用卡方测验来检验各方程的拟合度。
2 结果与分析
2.1 淡翅小花蝽对黄胸蓟马2龄若虫的捕食功能反应
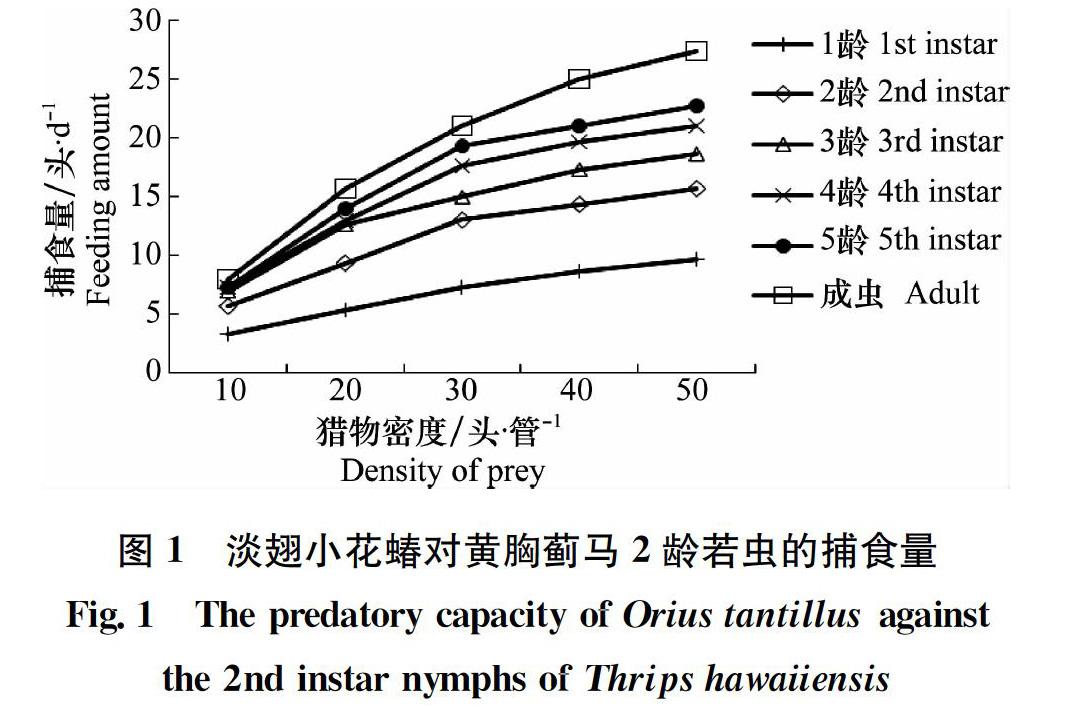
淡翅小花蝽各龄期若虫与成虫对黄胸蓟马2龄若虫的捕食情况见图1,结果显示淡翅小花蝽各龄期若虫与成虫的捕食量均随着黄胸蓟马若虫密度的增加而提高,但当猎物密度增加到一定水平后,其捕食量增速减缓,表明淡翅小花蝽对黄胸蓟马的捕食功能反应属于HollingⅡ型。根据HollingⅡ型圆盘方程拟合得到淡翅小花蝽对黄胸蓟马若虫的捕食功能反应模型及相关参数见表1。结果显示,所得方程的相关系数均较高(r=0.995 3~0.999 1),表明淡翅小花蝽各龄期若虫与成虫的捕食量均与黄胸蓟马若虫的密度显著相关。用拟合模型计算淡翅小花蝽对黄胸蓟马的理论捕食量,与实际捕食量进行卡方检验,所得χ2值(0.052~0.170)均远小于χ2(0.01,4)=13.277,表明所拟合的HollingⅡ型圆盘方程可反映不同黄胸蓟马密度下淡翅小花蝽捕食量的变化规律。
2.2 淡翅小花蝽对黄胸蓟马2龄若虫的搜寻效应
根据搜寻效应方程及表1相关参数,可计算得到淡翅小花蝽捕食黄胸蓟马的搜寻效应(图2)。结果表明淡翅小花蝽各龄期若虫与成虫对黄胸蓟马2龄若虫的搜寻效应均随猎物密度的增加而下降。此外,在相同猎物密度下,淡翅小花蝽成虫的搜寻效应最高,随着若虫虫龄的增大搜寻效应提高。
2.3 淡翅小花蝽的种内干扰
在黄胸蓟马2龄若虫密度不变的情况下,不同密度的淡翅小花蝽成虫对黄胸蓟马的捕食情况见表2。
结果表明淡翅小花蝽的总捕食量随其自身密度的增大而提高,但平均捕食量和捕食作用率均随其自身密度的增大而降低。根据干扰反应模型E=QP-m计算得到淡翅小花蝽成虫捕食黄胸蓟马2龄若虫过程中的自我干扰方程为E=0.357 3P-0.874 3,相关系数r=0.995 8,表明淡翅小花蝽的捕食作用率与其自身密度显著相关。经卡方检验,χ2=0.390(P=0.983)远小于χ2(0.01,4)=13.277,表明上述干扰方程可用于描述淡翅小花蝽成虫捕食黄胸蓟马2龄若虫过程中的自我干扰情况。
利用分摊竞争强度公式计算淡翅小花蝽成虫捕食黄胸蓟马2龄若虫所产生的分摊竞争强度(I)见表2和图3,淡翅小花蝽的分摊竞争强度(I)与其自身密度对数(lgP)之间的关系式为I=1.078 8lgP+0.074 5,相关系数r=0.966 1,表明两者显著相关。经卡方检验,χ2=0.051 9远小于χ2(0.01,3)=11.34,表明上述关系式能较好地描述淡翅小花蝽成虫捕食黄胸蓟马2龄若虫过程中的分摊竞争情况。由图3可知,淡翅小花蝽成虫捕食黄胸蓟马若虫时的种内分摊竞争强度随其自身密度对数值的增加而呈直线增加。
2.4 相互干扰对淡翅小花蝽成虫取食黄胸蓟马2龄若虫的影响
一定空间下,在淡翅小花蝽与黄胸蓟马密度同倍增加的情况下,根据搜寻效应与猎物密度及捕食者密度的函数关系E=Na/(N×P)计算淡翅小花蝽成虫对黄胸蓟马2龄若虫的搜寻效应,结果见表3。结果表明淡翅小花蝽的总捕食量随着捕食者密度、猎物密度的增加而升高,而捕食作用率却随着捕食者密度、猎物密度的增加而降低。
Beddington[25]提出,在考虑捕食者对猎物处理时间的情况下,搜寻效应(E)与猎物密度(N)、捕食者密度(P)之间的关系式为:E=a/[1+aThN+bTw(P-1)],将方程线性化为1/E=bTwR/a+1/a,用表3参数进行拟合后得到方程1/E=1.428 7R+1.032 5,(r=0.999 1),由此可求得a=0.968 5,bTw=1.383 7,所得数据代入上述模型得E=0.968 5/(0.011 0N+1383 7P-0.383 7),根据拟合的模型可求出淡翅小花蝽搜寻效应的理论值,与实际值进行卡方检验,χ2=0.051 4(P=0.999 7)远小于χ2(0.01,4)=13.277,表明所得模型能较好描述淡翅小花蝽的搜寻效应。由模型亦可知,淡翅小花蝽的搜寻效应随着自身密度和猎物密度的增加而下降,且受自身密度的影响较大。
3 讨论
天敌的捕食功能反应可用于评价天敌对害虫控害效能,其中天敌对害虫的瞬时攻击率与处理时间之比(a′/Th)可作为衡量参数之一,a′/Th越大,表明天敌的捕食能力越强,其对害虫的控制能力越强[28]。本研究结果显示淡翅小花蝽对黄胸蓟马2龄若虫具有较强的捕食能力,成虫的捕食能力最强,若虫随虫龄增大而提高,即成虫>5龄>4龄>3龄>2龄>1龄。相关研究结果表明东亚小花蝽对黄胸蓟马若虫的捕食能力也随其虫龄的增大而提高,3龄东亚小花蝽对黄胸蓟马若虫的捕食能力即达9200头[16],高于淡翅小花蝽所有参试虫态的捕食能力,表明东亚小花蝽较淡翅小花蝽对黄胸蓟马若虫的控制能力强。但鉴于淡翅小花蝽与黄胸蓟马的分布区域具有较大重叠,而与东亚小花蝽的分布区域大不相同,因此淡翅小花蝽在黄胸蓟马的生物防治上仍具有较大的潜能。
关于天敌的搜寻效应,目前大多数研究主要考虑天敌的搜寻效应与天敌自身密度关系或与害虫密度的关系[29-30],而仅有少数研究报道了天敌搜寻效应与天敌自身密度和害虫密度的关系[31-33]。本研究采用Beddingotn[25]推导出的搜寻效应与猎物密度和捕食者密度之间的关系式E=a/[1+aThN+bTw(P-1)],拟合得到淡翅小花蝽的搜寻效应模型E=0.968 5/(0.011 0N+1.383 7P-0.383 7),表明淡翅小花蝽搜寻效应随自身密度和猎物密度的提高而下降。故在田间释放淡翅小花蝽防治黄胸蓟马时,除了考虑蓟马的密度外,还应考虑淡翅小花蝽的释放密度,以达到最佳的控害效能。
本研究只开展了室内条件下一定空间内淡翅小花蝽对黄胸蓟马的捕食作用,但在自然条件下,天敌对害虫的控制能力除了受到天敌自身的密度和龄期、害虫的密度影响外,还受到天敌性别[34]、害虫龄期[16]、环境温度[35]、湿度[36]、农药使用[37]、寄主植物种类[38-39]等诸多因素的影响,因此,为了弄清淡翅小花蝽对黄胸蓟马的控制作用,今后仍需开展更深入的研究。
参考文献
[1] OSTMARK H E. Economic insect pests of banana [J]. Annual Review of Entomology, 1974, 19: 161-176.
[2] 蔡云鹏, 黄明道, 陈新评. 香蕉园内花蓟马之发生及其为害[J]. 中华昆虫, 1992, 12(4): 231-237.
[3] REYNAUD P, BALMS V, PIZZOL J. Thrips hawaiiensis (Morgan, 1913)(Thysanoptera: Thripidae), an Asian pest thrips now established in Europe [J]. EPPO Bulletin, 2008, 38(1): 155-160.
[4] 曾鑫年, 林进添. 黄胸蓟马对香蕉的危害及其防治[J]. 植物保护, 1998, 24(6): 15-17.
[5] 韩冬银, 邢楚明, 李磊, 等. 芒果花果期蓟马寄主植物间迁移对其种群数量增长的影响[J]. 昆虫学报, 2017, 60(6): 723-730.
[6] CAO Yu, LI Can, YANG Wenjia, et al. Effects of temperature on the development and reproduction of Thrips hawaiiensis (Thysanoptera: Thripidae)[J]. Journal of Economic Entomology, 2018, 111(2): 755-760.
[7] 張帆, 付步礼, 刘奎, 等. 温度对香蕉花蓟马发育和存活的影响[J]. 生态学报, 2014, 34(14): 3895-3899.
[8] 付步礼, 刘俊峰, 邱海燕, 等. 海南省香蕉黄胸蓟马田间种群的抗药性监测[J]. 应用昆虫学报, 2016, 53(2): 403-410.
[9] FU Buli, LI Qiang, QIU Haiyan, et al. Resistance development, stability, cross-resistance potential, biological fitness and biochemical mechanisms of spinetoram resistance in the Thrips hawaiiensis (Thysanoptera: Thripidae)[J]. Pest Management Science, 2018, 74(7): 1564-1574.
[10]王大生, 张帆. 国内外天敌昆虫产业现状[C]∥罗晨, 季延寿. 第五届生物多样性保护与利用高新科学技术国际研讨会论文集.北京: 北京科学技术出版社, 2005: 44-48.
[11]张鲁民, 刘志诚, 孙兴全, 等. 南方小花蝽和花蓟马种群季节动态及捕食功能反应[J]. 中国生物防治, 2008, 24(S1): 21-27.
[12]李向永, 陈福寿, 赵雪晴, 等. 微小花蝽的发生及其对西花蓟马的捕食作用[J]. 环境昆虫学报, 2011, 33(3): 346-350.
[13]莫利锋, 郅军锐, 田甜. 南方小花蝽在不同空间及笼罩条件下对西花蓟马的控制作用[J]. 生态学报, 2013, 33(22): 7132-7139.
[14]王然, 王晓灵, 王甦, 等. 东亚小花蝽对西花蓟马的控害能力评价[J]. 环境昆虫学报, 2014, 36(6): 983-989.
[15]吕兵, 孙猛, 翟一凡, 等. 基于捕食功能反应评价麦蛾卵饲养东亚小花蝽对棕榈蓟马的控害效果[J]. 植物保护学报, 2017, 44(5): 875-876.
[16]付步礼, 邱海燕, 李强, 等. 东亚小花蝽对黄胸蓟马的室内捕食作用研究[J]. 应用昆虫学报, 2019, 56(1): 91-98.
[17]梁旻雯. 中国花蝽科与锤角叶蜂科生物地理研究 [D].长沙: 中南林业科技大学, 2006.
[18]张骏, 郅军锐, 杨成源, 等. 贵州省农田小花蝽种类调查及鉴定[J]. 山地农业生物学报, 2015, 31(1): 36-40.
[19]CALILUNG V J, MALABAYABASARCELO M, CALILUNG M V J. Occurrence of Thrips palmi Karny (Thysanoptera: Thripidae) and its predator, Orius tantillus (Motschulsky)(Hemiptera: Anthocoridae)in the Philippines [J]. Philippine Agriculturist, 1999, 80(1/2): 33-42.
[20]MITUDA E C, CALILUNG V J. Biology of Orius tantillus (Motschulsky)(Hemiptera: Anthocoridae)and its predatory capacity against Thrips palmi Karny(Thysanoptera: Thripidae)on watermelon [J]. Philippine Agriculturist, 1989, 72(2): 165-184.
[21]NAVASERO M M, MORALLO-REJESUS B, NAVASERO M V. Field evaluation of the predatory bug, Orius tantillus (Mots), (Anthocoridae:Hemiptera) for the control of the Asian corn borer, Ostrinia furnacalis (Guenée)[J]. Philippine Entomologist, 2004, 7(2): 185-186.
[22]BALLAL C R, GUPTA T, JOSHI S. Effect of constant temperature regimes on the biological parameters of an anthocorid predator Orius tantillus (Motsch) [J].Journal of Biological Control, 2017, 31(3): 146-158.
[23]HOLLING C S. Some characteristics of simple type of predation and parasitism [J]. Canadian Entomologist, 1959, 91(7): 385-398.
[24]丁岩钦. 昆虫数学生态学[M]. 北京: 科学出版社, 1994: 257-258, 303-304.
[25]BEDDINGTON J R. Mutual interference between parasites or predators or predators and its effect on searching efficiency [J]. Journal of Animal Ecology, 1975, 44(1): 331-340.
[26]HASSELL M P, VARLEY G C. New inductive population model for insect parasites and its bearing on biological control [J]. Nature, 1969, 223(5211): 1133-1137.
[27]邹运鼎, 耿继光, 陈高潮, 等. 异色瓢虫若虫对麦三叉蚜的捕食作用[J]. 应用生态学报, 1996, 7(2): 197-200.
[28]周集中, 陈常铭. 拟环纹狼蛛对褐飞虱的捕食作用及其模拟模型的研究Ⅰ. 功能反应[J].生物防治通报, 1986, 2(1): 2-9.
[29]王圣印, 牛雨佳, 唐睿, 等. 短翅蚜小蜂对桃蚜的取食和寄生功能反应[J]. 植物保护学报, 2016, 43(2): 267-274.
[30]唐良德, 王晓双, 赵海燕, 等. 大草蛉幼虫捕食豆大蓟马和豆蚜的功能反应及生长发育[J]. 中国生物防治学报, 2017, 33(1): 49-55.
[31]杨燕燕, 李照会, 王如刚, 等. 异色郭公虫对柏肤小蠹捕食作用的研究[J]. 山东农业科学, 2004(6): 40-42.
[32]刘萍, 沈平, 吴建华, 等. 迭球螋成虫对棉蚜的捕食作用[J]. 植物保护, 2014, 40(5): 70-74.
[33]马艳芳, 张永强, 常承秀, 等. 迭球螋对落叶松球蚜成虫的捕食作用[J]. 植物保護, 2016, 42(1): 116-118.
[34]金剑雪, 李凤良, 程英, 等. 七星瓢虫对豆蚜的功能反应[J]. 植物保护, 2011, 37(4): 68-71.
[35]贾静静, 符悦冠, 张方平, 等. 温度对加州新小绥螨捕食东方真叶螨功能反应的影响[J]. 中国生物防治学报, 2019, 35(3): 382-389.
[36]李鸿筠, 刘浩强, 姚廷山, 等. 不同条件下日本方头甲对矢尖蚧捕食功能的反应[J]. 植物保护学报, 2009, 36(6): 491-496.
[37]李锐, 李娜, 刘佳, 等. 低剂量杀虫剂对星豹蛛捕食效应的影响及其机理[J]. 生态学报, 2014, 34(10): 2629-2637.
[38]王甦, 渠成, 李济航, 等. 大草蛉幼虫对不同寄主植物上烟粉虱卵的捕食功能反应与搜寻效应[J]. 植物保护学报, 2016, 43(1): 149-154.
[39]DE PATRICK C, MOHAGHEGH J, LUC T. Effect of host plant on the functional response of the predator Podisus nigrispinus (Heteroptera: Pentatomidae)[J]. Biological Control, 2000, 18(1): 65-70.
(责任编辑:田 喆)
