广西人群LncRNA-GAS5基因rs6790A/G和rs1951625A/G位点多态性分布①
2020-12-25刘纯宏陆玉兰黄华佗王春芳韦叶生
刘纯宏 王 艳 石 凤 陆玉兰 黄华佗 王春芳 韦叶生 蓝 艳
(右江民族医学院附属医院检验科,百色533000)
长链非编码RNA(long non-coding RNA,LncRNA)是一类长度大于200个核苷酸且缺乏开放阅读框的新型非编码RNA[1]。最初,LncRNA被认为是“基因垃圾”“暗物质”或“假基因”,不具有生物学功能。最新研究表明,LncRNA在转录及转录后水平调控基因表达,调控细胞增值、分化及凋亡过程。生长停滞特异性转录本5(growth arrest specific transcript 5,GAS5)最初被发现积聚于生长停滞细胞中的LncRNA,位于人类染色体1q25.1,由12个外显子构成,可编码10个C/D snoRNA,全长651个核苷酸。最新研究表明,GAS5与系统性红斑狼疮、哮喘、类风湿关节炎和骨关节炎等免疫相关疾病的发生密切相关,提示GAS5可能是某些免疫炎症疾病的防治靶标或生物标记物[2-5]。目前,关于GAS5基因rs6790A/G和rs1951625A/G遗传多态性在广西人群中分布特征的研究尚未见报道。因此,本研究采用SNPscan技术对广西健康人群LncRNA-GAS5基因rs6790A/G和rs1951625A/G位点进行基因分型检测,并分析其多态性与其他地区人群的分布差异,为阐明GAS5基因多态性在不同种族之间的分布特征及其与疾病的关系提供群体遗传学参考。
1 资料与方法
1.1资料 随机选取2017年5月~2018年9月我院健康体检者289例,其中男性106例,女性183例,年龄19~76岁。血常规及生化指标均在正常参考范围内,结合影像学检查及临床医师诊断,排除自身免疫、神经系统、肿瘤、肝脏、肾脏、心脏等疾病;研究对象均为世居于广西地区且相互间无血缘关系的个体。本研究方案获得我院伦理委员会批准,所有受试者知情同意。
1.2方法
1.2.1基因组DNA提取 用EDTA-K2抗凝管采集各受试者静脉血3.0 ml,采用离心柱法提取基因组DNA,-70 ℃保存备用。连接引物的设计与合成从NCBI(https://www.ncbi.nlm.nih.gov/)数据库中查找LncRNA GAS5基因rs6790A/G和rs1951625A/G位点的碱基序列,采用primer3在线软件(http://bioinfo.ut.ee/primer3-0.4.0/)辅助设计实验引物序列(表1),委托上海天昊生物科技有限公司合成。
1.2.2连接和多重PCR反应及基因分型检测 连接反应体系总体积为20.0 μl,其中含样本DNA 8.0 μl、2× 连接缓冲液 10.0 μl、耐热DNA连接酶0.2 μl、连接探针混合液0.8 μl、 剩余体积用灭菌双

表1 GAS5基因rs6790A/G和rs1951625A/G引物序列
蒸水补足。连接反应参数:98℃ 2 min;95℃ 30 s,58℃ 3 h;循环5次;94℃ 2 min;72℃ forever。将连接产物进行多重荧光PCR扩增,多重PCR反应体系的总体积为20.0 μl,其中包含连接产物1.0 μl,2×PCR缓冲液A 10.0 μl,TaqDNA聚合酶0.4 μl,多重荧光扩增引物0.5 μl,灭菌双蒸水8.1 μl。多重PCR反应参数:95℃ 2 min;94℃ 20 s,62℃ 40 s(每个循环降低0.5℃),72℃ 1.5 min,循环9次;94℃ 20 s,57℃ 40 s,72℃ 1.5 min,循环25次;68℃ 60 min;4℃ forever。将多重PCR产物以10倍ddH2O稀释,振荡混匀,取1.0 μl PCR产物与 0.1 μl LIZ500和8.9 μl Hi-Di混匀,95℃变性 5 min,后ABI3500测序仪检测,采用GeneMapper 4.1软件进行数据分析。
1.3统计学分析 采用SPSS22.0软件对实验数据进行统计学分析。采用Hardy-Weinberg遗传平衡定律检验样本的群体代表性。基因型和等位基因频率采用直接计数法计算,不同组间基因型和等位基因频率的比较用行χ2检验和四格表χ2检验,以P<0.05为差异有统计学意义。
2 结果
2.1广西人群GAS5基因rs6790A/G和rs19516 25A/G分型检测结果 结果显示,rs6790A/G位点存在AA、AG和GG 3种基因型,频率分别为15.6%、48.1%和36.3%,A、G等位基因频率分别为39.6%和60.4%。rs1951625A/G位点存在AA、AG和GG 3种基因型,频率分别为8.3%、38.1%和53.6%,A、G等位基因频率分别为27.3%和72.7%。2个位点的基因型分布均符合Hardy-Weinberg遗传平衡定律(P>0.05),表明本研究所选取的人群具有群体代表性。基因分型检测结果见图1。
2.2rs6790A/G和rs1951625A/G位点多态性在广西人群不同性别间的分布 广西人群GAS5基因rs6790A/G和rs1951625A/G位点分别以AG和GG基因型为多见,其频率分别为48.1%和53.6%;G等位基因频率最高,分别为60.4%和72.7%。2位点的基因型和等位基因频率在男性和女性间的分布差异无统计学意义(P>0.05),提示2个位点基因型和等位基因频率在广西人群中的分布与性别无关,见表2。
2.3广西人群rs6790A/G和rs1951625A/G多态性与其他地区人群比较 将广西人群rs6790A/G和rs1951625A/G位点的基因型和等位基因频率分别与国际千人基因组计划数据库公布的CHS、JPT、CEU、AFR和AMR人群进行比较,结果显示,广西人群rs6790A/G位点的基因型和等位基因频率与JPT、CEU、ARF和AMR人群比较差异均有统计学意义(P<0.05),与CHS人群比较差异无统计学意义(P>0.05);广西人群rs1951625A/G位点的基因型和等位基因频率与AFR人群比较差异有统计学意义(P<0.05),与CHS、JPT、CEU和AMR人群比较差异均无统计学意义(P>0.05),结果见表3、4。
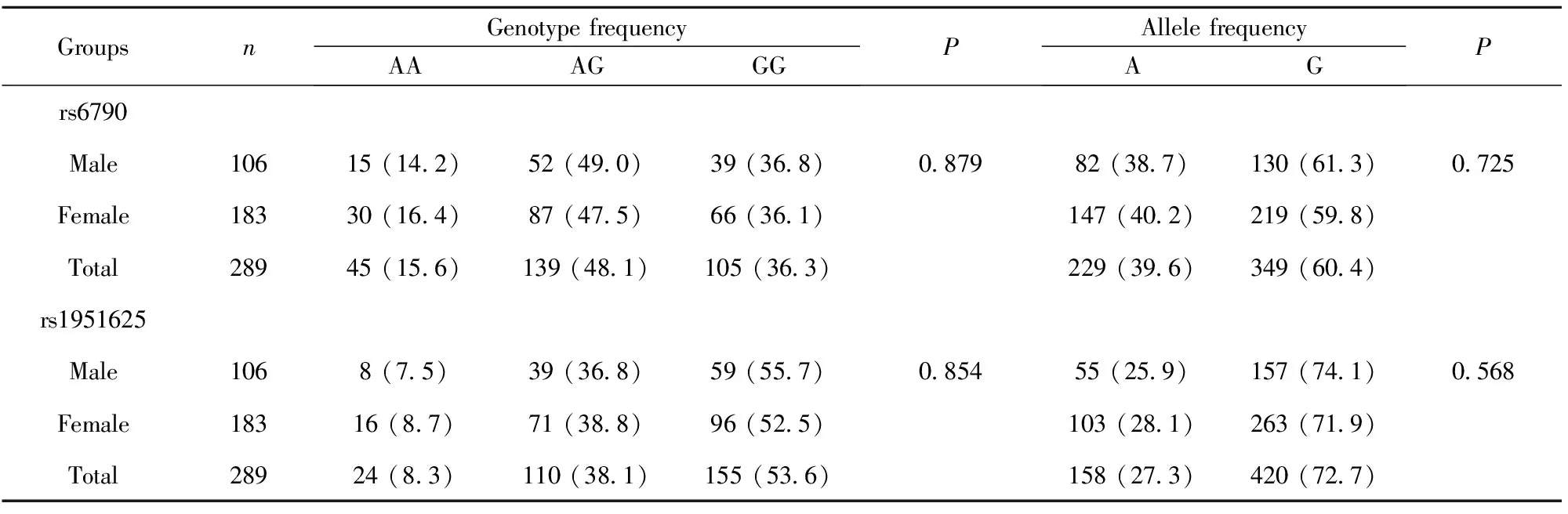
表2 rs6790A/G和rs1951625A/G位点基因型和等位基因频率在广西人群不同性别间比较[例(%)]
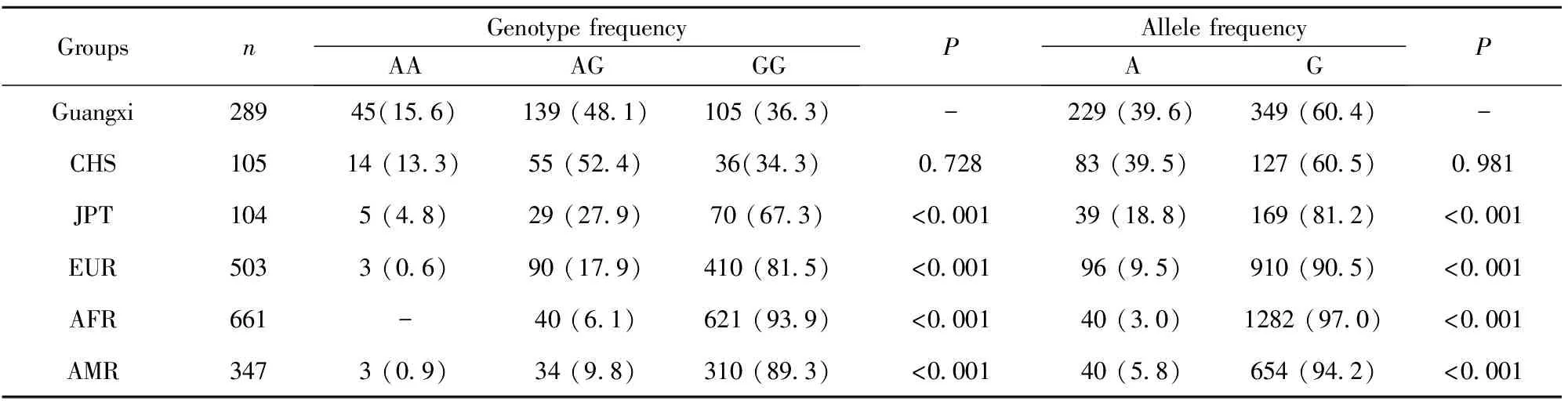
表3 广西人群GAS5基因rs6790A/G遗传多态性与不同人群的比较[例(%)]

表4 广西人群GAS5基因rs1951625A/G遗传多态性与不同人群的比较[例(%)]
3 讨论
LncRNA-GAS5最初被发现与细胞生长阻滞密切相关,全长651个核苷酸,其编码基因GAS5属于5′-top家族成员,其外显子不具有编码能力,而内含子可编码10box C/D的miRNAs并发挥生物学作用[6]。GAS5第400~598个核苷酸含有6对短发夹结构,且第5对发夹结构含有2条糖皮质激素受体反应元件 (glucocorticoid receptor response element,GRE) 序列,称为GAS5 GRE-mimic[7]。由于GAS5具有相对复杂的二级结构,可能是其具有复杂生物学功能的结构基础。近年GAS5基因与疾病的关系引起研究者关注,成为基因研究的新兴领域。Wang等[7]研究发现GAS5基因启动子区的rs55829688位点突变与结直肠癌发生相关,并通过影响转录因子YY1与GAS5的结合亲和力降低GAS5表达。Tang等[8]研究发现GAS5基因rs145204276位点del等位基因可能通过与转录因子特异性蛋白1结合,诱导启动子活性,从而抑制乳腺癌发生,导致GAS5水平升高。伊朗学者Rakhshan等[9]研究发现GAS5基因rs2067079位点隐性遗传模型(TT vs.CC + CT)及rs2067079和rs6790位点单倍型组合T-G可能增加膀胱癌发病风险。
遗传多态性可因自然选择、环境因素、饮食习惯和不同种族呈现不同程度的差异,现有研究表明,不同遗传背景的人群对相同疾病易感性不同。如埃及学者Shaker等[10]研究发现LncRNA-CASC8基因rs6983267位点多态性可能提高大肠癌遗传易感性(GG vs.GT/TT:OR=2.13,95%CI=1.146~3.937,P=0.017);而西班牙学者Carla等[11]研究发现rs6983267位点多态性可能降低大肠癌发病风险(GG vs.GT/TT:OR=0.69,95%CI=0.50~0.95,P<0.001)。出现此差异性结果的原因可能是研究对象遗传背景不同。因此,研究不同地区和族群间正常人群的基因多态性分布特征,可阐明不同地区人群的遗传背景资料,也为进一步研究不同人群基因多态性与某些疾病的遗传易感性提供理论依据和数据支持。
本研究采用SNPscan测序技术检测广西人群LncRNA-GAS5基因rs6790A/G和rs1951625A/G位点多态性,结果显示,广西人群GAS5基因rs6790A/G和rs1951625A/G位点基因型和等位基因频率分布在不同性别间差异均无统计学意义(P>0.05),提示GAS5基因rs6790A/G和rs1951625A/G位点在广西人群中的遗传多态性分布与性别无关。进一步将广西人群rs6790A/G和rs1951625A/G位点多态性与千人基因组计划数据库公布的CHS、JPT、CEU、AFR和AMR人群的SNP分型数据进行比较,发现广西人群rs6790A/G位点的基因型和等位基因频率与JPT、CEU、ARF和AMR人群比较差异均有统计学意义(P<0.05),与CHS人群比较差异无统计学意义(P>0.05);广西人群rs1951625A/G位点的基因型和等位基因频率与AFR人群比较差异有统计学意义(P<0.05),与CHS、JPT、CEU和AMR人群比较差异均无统计学意义(P>0.05)。国内外多项研究已证实基因多态性与地缘关系、生活习性及种族等多种因素密切相关,与本研究结果相符[12-14]。广西人群与CHS和JPT人群同属于亚洲人群,地缘关系相对较近,因此,这3种人群的遗传多态性分布差异相对较小,与“亲缘关系越近,其基因型的分布越相似,反之则差异越显著”的遗传学定律相符。与此同时,本研究发现JPT人群与广西地区人群虽然都是亚洲人群,但由于广西是多民族聚集的偏远山区,地理环境及气候独特,生活方式及饮食习惯与其他地区也存在较大差异,因此,2种人群的遗传多态性也呈现差异性。而广西人群与CEU、AFR和AMR人群属于不同种族人群,3者亲缘关系较远,地理及生态隔离阻碍了种族间遗传物质的交流,随着时间积累,各隔离群体发生的遗传变异朝着不同的方向发展,可能是造成3种人群存在较明显的遗传多态性差异的原因之一。rs1951625A/G位点多态性与AFR人群差异有统计学意义,而与CHS、JPT、CEU和AMR人群差异均无统计学意义的原因之一可能与“人类起源学说”有关,研究认为人类在非洲具有1个共同祖先,而随着气候、地理等因素人类逐渐走出非洲,到达世界各地[15]。现代人类经过漫长的迁移,与不同地区原有的近缘同类混血通婚,并独立进化,形成不同族群,随着时间的积累,不同群体与非洲人群间的差异越来越明显。因此,本研究认为出现此差异结果的原因可能与人类历史上大规模的迁徙、自然的选择及基因的随机漂变有关。提示广西人群存在GAS5基因多态性,且与其他地区人群比较存在不同程度的差异。
本研究初步探讨了广西人群GAS5基因rs6790A/G和rs1951625A/G位点多态性分布特征,并与国际千人基因组计划公布的不同地区人群的SNP分型数据进行了比较,不仅为GAS5基因多态性的群体遗传学普查提供了数据资料,同时也为其与相关疾病易感性的研究奠定了遗传学基础。但广西是多民族聚集区,本研究未能具体到民族,有待进一步研究。
