饥饿胁迫和再投喂对俄罗斯鲟形体和血液指标的影响
2020-12-21鹿王成志蒲红宇王晓雯朱华
鹿王成志 蒲红宇 王晓雯 朱华

摘 要:为探讨饥饿胁迫对俄罗斯鲟(Acipenser gueldenstaedtii)形体和血液生化指标的影响,以室内养殖俄罗斯鲟为研究对象进行饥饿胁迫和再投喂实验,分别在饥饿7、14、21、28 d和恢复投喂后20 d,分析形体和血液指标变化情况。研究发现,饥饿胁迫和复投喂过程中,俄罗斯鲟体长和体质量变化不显著,肝指数和肥满度显著下降(P<0.01),肝质量在短期饥饿胁迫后相对稳定,而随着饥饿时间延长显著下降(P<0.01)。饥饿胁迫条件下,俄罗斯鲟血清谷丙转氨酶、谷草转氨酶和碱性磷酸酶活性以及总蛋白和白蛋白含量保持稳定。饥饿胁迫和再投喂过程中,血清肌酐和尿素显著下降(P<0.01),长期饥饿胁迫和复投喂后甘油三酯水平显著下降(P<0.01)。结果表明,饥饿胁迫对俄罗斯鲟肝脏的影响主要表现为肝质量和肝指数下降,肝细胞结构未受损伤,肝脏合成贮存功能未受影响;饥饿胁迫对俄罗斯鲟能量代谢的影响主要体现在嘌呤代谢、肌肉代谢和脂肪代谢过程。
关键词:俄罗斯鲟(Acipenser gueldenstaedtii)饥饿胁迫;形体指标;血液生化指标
俄罗斯鲟(Acipenser gueldenstaedtii)属鲟形目,鲟科,鲟属,为营底栖生活的冷水性鱼类[1]。鲟鱼是目前世界上最古老的鱼类之一,距今已有两亿多年的历史,素有“活化石”之称。鲟鱼浑身上下都是宝,其卵做成的鱼子酱有“黑色黄金”的美誉[2]。我国自20世纪90年代开展鲟鱼人工养殖研究,目前,随着鲟鱼养殖技术的推广,我国鲟鱼商品鱼年产量占世界总养殖产量的80%左右,成为世界上最重要的鲟鱼生产大國[3]。
人工养殖过程中,鱼类体型大小差异、养殖密度过大、投喂饲料成分不当、投喂密度不均等因素都会造成鱼类饥饿现象[4]。目前,关于养殖过程中鲟鱼受饥饿影响的研究尚未见报道。为此,本研究以俄罗斯鲟为研究对象,探讨饥饿胁迫和复投喂对俄罗斯鲟形体指标和血液生化指标的影响,旨在丰富俄罗斯鲟生理学的基础指标,为俄罗斯鲟的健康养殖提供科学依据。
1 材料和方法
1.1 实验材料
俄罗斯鲟在北京市水产科学研究所鲟鱼养殖室养殖。挑选体长相近、体重均一的活泼健康个体用于实验。随机挑选40条俄罗斯鲟,转移到1.8 m×1 m×0.6 m养殖箱内,驯化14 d后,进行28 d饥饿胁迫处理,28 d后恢复投喂。养殖箱保持不间断自动水循环,水温(4±2)℃,pH 值7.5±0.5。
1.2 实验方法
样本采集:分别在饥饿胁迫第0、7、14、21、28 d,以及恢复喂食第20 d时,随机取3尾俄罗斯鲟,立即用MS-222(Sigma,CAS 886-86-2)溶液麻醉,无菌注射器尾静脉采血2 mL。血液样本室温下静置1 h,4 ℃过夜,随后3 000 r/min离心5 min,吸取上层血清,-80 ℃保存备用。
形体指标检测:采血前对用于采样的俄罗斯鲟进行称重,测量全长,其中全长是指从吻端至尾鳍未端的长度。采血后进行解剖,记录内脏器官总质量和肝脏质量,计算形体指标。其中:
肥满度=体质量/全长
脏体比=内脏质量/体质量
肝体比=肝质量/体质量×100
能量含量(kJ/g)=脂肪(g)×39.5(kJ/g)+蛋白质(g)×23.6(kJ/g)
水分含量用常压干燥法(GB/T 5009.3-2003)测定,粗蛋白含量用微量凯氏定氮法(GB/T 5009.5-2003)测定,粗脂肪含量用全脂肪法(GB/T 5009.6-2003)测定,灰分含量用灼烧称重法(GB/T 5009.3-2003)测定。
血清葡萄糖、甘油三酯、胆固醇、尿素、尿酸、肌酐、总蛋白和白蛋白含量采用自动生化分析仪(贝克曼,Cx-4)测定。谷丙转氨酶、谷草转氨酶、碱性磷酸酶、乳酸脱氢酶等酶活力的检测,采用试剂盒法,试剂盒购买自南京建成生物有限公司。
1.3 统计分析
所有数据采用平均值±标准差表示。单因素方差分析法进行组间显著性分析,Duncans方法进行事后多重检验,P<0.05 为差异显著,P<0.01为差异极显著。
2 实验结果
2.1 俄罗斯鲟形体指标的变化分析
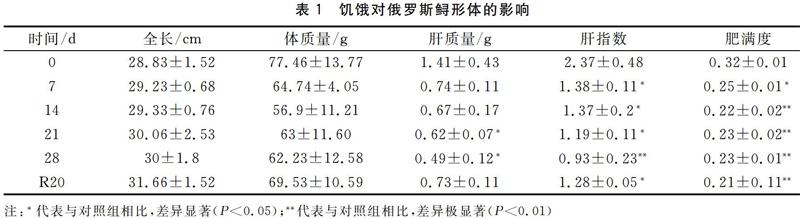
饥饿胁迫和恢复投喂期间,俄罗斯鲟全长和体质量变化不显著。体重变化不显著。肝质量在饥饿胁迫21 d和28 d时显著下降(P<0.05),恢复投喂20 d后有所回升。肝指数在饥饿胁迫7 d开始显著下降(P<0.05),28 d后极显著低于对照组(P<0.01);值得注意的是,恢复投喂20 d后肝指数依然显著低于对照组(P<0.05)。肥满度在饥饿胁迫7 d后开始显著下降(P<0.05),随后在饥饿胁迫期以及恢复投喂后20 d时依然极显著低于对照组(P<0.01)。研究结果表明,饥饿胁迫显著影响俄罗斯鲟肝指数和肥满度。见表1。
2.2 俄罗斯鲟肝脏酶活性变化分析
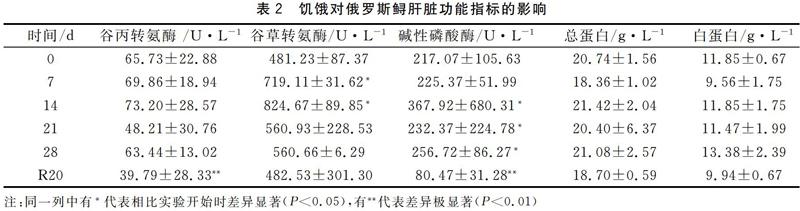
在饥饿胁迫和恢复喂食阶段,俄罗斯鲟血清谷丙转氨酶(ALT)活性在恢复投喂后20 d后极显著低于对照组水平(P<0.01)。碱性磷酸酶(ALP)活性在第14 d,21 d,28 d显著高于对照组水平(P<0.01),在恢复投喂后极显著低于对照组水平(P<0.01)。谷草转氨酶(AST)在实验第7 d,第14 d相比实验开始显著高于对照组(P<0.05)。饥饿胁迫和恢复投喂后血清总蛋白和白蛋白含量维持稳定,没有明显变化。结果表明,饥饿胁迫对俄罗斯鲟AST、ALT和ALP酶活性有显著影响,对血清总蛋白和白蛋白含量没有显著影响。见表2。
2.3 俄罗斯鲟能量代谢指标的变化分析
饥饿胁迫处理中和恢复投喂后20 d时,俄罗斯鲟血糖含量维持稳定,表明饥饿胁迫对俄罗斯鲟血糖含量没有显著影响。饥饿胁迫14 d后尿素含量显著下降,在恢复投喂20 d后依然显著低于对照组(P<0.05)。饥饿胁迫处理过程中血清中肌酐含量极显著低于对照组(P<0.01),恢复投喂20 d后依然显著低于对照组(P<0.05)。饥饿胁迫前期,甘油三酯含量变化不显著,饥饿28 d和恢复投喂20 d时甘油三酯含量显著低于对照组(P<0.05)。饥饿胁迫和恢复投喂后,乳酸脱氢酶第14 d显著高于对照组(P<0.05),活性在恢复投喂20 d后极显著低于对照组水平(P<0.01)。尿酸除了第14 d之外,都低于对照组水平,饥饿胁迫28 d与恢复投喂后含量显著低于对照组(P<0.01)。见表3。
3 讨论
3.1 饥饿胁迫和再投喂影响俄罗斯鲟形体指标
鱼类的肥满度在很大程度上决定了鱼类的品质好坏,从而影响其商品价值。阐明饥饿胁迫对俄罗斯鲟肥满度等形体指标的影响,对俄罗斯鲟的人工养殖有重要的指导意义。本研究发现,随着饥饿时间的延长,俄罗斯鲟的肥满度逐渐下降,饥饿14 d后肥满度下降31.25%。饥饿28 d并恢复投喂后相比初始值依然下降了34.37%。研究报道,西洋鲑魚(Salmo salar)经饥饿83 d后体重降低11.3%[5]。刀鲚(Coilia nasus)饥饿20 d体重降低3.8%,肥满度下降23.33%[6]。鳙(Aristichthys nobilis)饥饿25 d后肥满度下降34.45%[7]。结果表明,俄罗斯鲟的饥饿耐受能力明显低于大西洋鲑、刀鲚和鳙等。
肝脏是鱼类重要的代谢器官,也是主要的营养储藏器官,在营养不良时,肝质量将发生显著变化。本研究发现,饥饿28 d后俄罗斯鲟肝质量下降65.24%,肝指数下降60.75%,表明长期饥饿胁迫对俄罗斯鲟肝质量具有显著影响,这可能与肝脏是俄罗斯鲟储能器官有关。
3.2 饥饿胁迫和再投喂下俄罗斯鲟肝脏功能变化
血清生化指标的变化是鱼类生理状态最重要的指示因子,血清酶活性影响鱼类多种生化反应过程,因此血清酶活性可以用于评估鱼类生理状态。其中,AST、ALT和ALP酶活性变化是细胞膜损伤的敏感性分子标志物[8]。AST、ALT分别参与氨基酸和碳水化合物的三羧酸循环,ALP在水生动物骨骼组织矿物质转运过程中发挥重要作用,其活性的变化表征肝功能的变化[9]。本研究发现,在饥饿胁迫阶段,俄罗斯鲟血清ALT活性几乎不变,AST与ALP活性有较大提升,但是在恢复投喂之后都下降,其中ALT与ALP显著下降。表明饥饿胁迫后再投喂的俄罗斯鲟肝脏功能可以恢复正常,这可能是由于28 d饥饿胁迫尚不造成俄罗斯鲟肝脏细胞结构水平的损伤。
血清总蛋白和白蛋白主要由肝脏合成[10],其含量变化反映了肝脏的合成和贮备功能。一般认为,饥饿会导致血清总蛋白和白蛋白含量下降[11-12]。然而近来研究发现,饥饿条件下鱼类血清总蛋白和白蛋白含量变化具有物种特异性。钱云霞等[13]研究发现,养殖鲈鱼血清总蛋白和白蛋白随着饥饿时间的延长,表现出先上升后下降的趋势。本研究发现,饥饿胁迫期间俄罗斯鲟血清总蛋白和白蛋白含量没有显著变化,表明长期饥饿胁迫(28 d)对俄罗斯鲟肝脏合成贮备功能没有显著影响。
3.3 饥饿胁迫和再投喂影响俄罗斯鲟鱼能量代谢
尿素是鱼类嘌呤代谢的最终代谢产物[14-15]。嘌呤代谢生成尿酸,在尿酸氧化酶的作用下被氧化生成尿囊素,然后经尿囊素酶作用生成尿囊酸。对大多数鱼类,尿囊酸还需经尿囊酸酶的作用水解生成尿素而排出体外[16]。因此血清尿素和尿酸含量可以反映俄罗斯鲟嘌呤代谢情况。饥饿胁迫下俄罗斯鲟尿素含量显著下降,表明饥饿胁迫可能导致俄罗斯鲟嘌呤代谢能力下降。
肌酐是肌酸和磷酸肌酸代谢最终产物,肌酸主要通过不可逆的非酶脱水反应缓缓地生成肌酐,再释放到到血液中,血清肌酐含量反映肌肉代谢情况,表明体内含氮性代谢产物的产生[17]。饥饿胁迫下俄罗斯鲟血清肌酐含量显著下降,表明饥饿胁迫导致俄罗斯鲟肌肉代谢能力下降,该结果可能进一步导致肌肉活动减少。
甘油三酯是长链脂肪酸和甘油形成的脂肪分子。脂肪组织中的甘油三酯在一系列脂肪酶的作用下,分解生成甘油和脂肪酸,并释放入血液供其它组织利用。本研究发现,长期饥饿胁迫(28 d)下,俄罗斯鲟血清甘油三酯水平显著下降。该结果表明,饥饿胁迫下俄罗斯鲟可能持续利用体内存储的脂肪为机体提供能量,饥饿持续时间过长,存储脂肪含量下降,可利用的甘油三酯含量降低,俄罗斯鲟难以通过脂肪代谢为机体提供充足能量。复投喂20 d后甘油三酯水平保持继续下降趋势,表明俄罗斯鲟体内脂肪的合成和存储是一个缓慢的过程,长期饥饿后难以通过短暂投喂恢复正常水平。血清甘油三酯含量变化进一步解释了饥饿期间俄罗斯鲟肥满度持续下降的原因。
4 结论
本实验中研究了饥饿胁迫对俄罗斯鲟形体指标和血液指标的影响,发现长期饥饿胁迫俄罗斯鲟体长和体质量的影响较小,但是导致肝质量和肝指数下降,肝脏在细胞结构上和合成贮存功能上没有显著变化。饥饿胁迫显著影响俄罗斯鲟能量代谢,长期饥饿胁迫后嘌呤代谢和肌肉代谢能力下降。饥饿胁迫过程中俄罗斯鲟大量消耗脂肪,导致甘油三酯含量大幅下降。
参考文献:
[1] 朱婷婷,李琦,朱浩拥,等.饲料脂肪水平对俄罗斯鲟幼鱼生长、血液生化指标及抗氧化性能的影响[J].海洋渔业.2017,39(01):58-67.
[2] 东天,胡红霞.鲟鱼育种技术之分子标记辅助育种的应用与展望[J].中国水产.2018(05):102-104.
[3] 董颖,胡红霞.鲟鱼种质鉴定技术的发展及应用[J].中国水产.2014(10):70-72.
[4] 张怡波,吕慧明,袁子,等.饥饿再投喂对黑鱾幼鱼消化酶活性与血液生理生化指标的影响[J].宁波大学学报(理工版).2019,32(05):12-16.
[5] EINEN O,WAAGAN B,THOMASSEN M S.Starvation prior to slaughter in Atlantic salmon (Salmo salar) I:Effects on weight loss,body shape,slaughter- and fillet-yield,proximate and fatty acid composition[J].Aquaculture.1998,166(1-2):1-104.
[6] 金鑫,徐钢春,杜富宽,等.饥饿胁迫对刀鲚形体、体成分及血液生化指标的影响[J].动物学杂志.2014,49(06):897-903.
[7] 钟金香,李俊伟,颉晓勇,等.饥饿处理对鳙鱼形态和肌肉营养成分的影响分析[J].基因组学与应用生物学.2019,38(08):3515-3520.
[8] TAVANA G Z B,BANAEE M,JOURDEHI Y A,et al.Effects of selenium (Sel-Plex) supplement on blood biochemical parameters of juvenile Siberian sturgeon (Acipenser baerii)[J].Iranian Journal of Fisheries Sciences.2018,17(2):300-312.
[9] GHOLAMI-SEYEDKOLAEI S J,MIRVAGHEFI A,FARAHMAND H,et al.Effect of a glyphosate-based herbicide in Cyprinus carpio:Assessment of acewlcholinesterase activity,hematological responses and serum biochemical parameters[J].Ecotoxicology and Environmental Safety.2013,98:135-141.
[10] 孙委委.斑马鱼早期胚胎/幼鱼急相期反应基因表达模式研究及血清淀粉样蛋白A(SAA)功能分析[D].中国海洋大学,2014.
[11] 周兴华, 欧仁建, 向桢,等. 饥饿和再投喂对中华倒刺鲃幼鱼生长和血清指标的影响[C]// 厦门:2011年中国水产学会学术年会.
[12] 杨严鸥,姚峰,余文斌,等.饥饿时间对越冬期黄颡鱼血清及鱼体生化成分的影响[J].长江大学学报(自科版)农学卷.2007,4(04):42-44.
[13] 钱云霞,陈惠群,孙江飞.饥饿对养殖鲈鱼血液生理生化指标的影响[J].中国水产科学.2002(02):133-137.
[14] ROZEMA E,KIERSZNIOWSKA S,ALMOG-GABAI O,et al.Metabolomics reveals novel insight on dormancy of aquatic invertebrate encysted embryos[J].Sci Rep.2019,9(1):8878.
[15] NOGUCHI T,FUJIWARA S,TAKADA Y,et al.Metabolism of urea and glyoxylate,degradative products of purines in marine animals[J].Journal of Biochemistry.1982,92(2):525-529.
[16] 张宇,丛艺,张士璀.文昌鱼嘌呤代谢酶基因:结构与演化[J].中国海洋大学学报(自然科学版).2009,39(01):96-104.
[17] 孔庆辉,晁燕,夏明哲,等.黄河裸裂尻鱼肌酸激酶M-CK cDNA的克隆及組织表达分析[J].动物学杂志.2016,51(01):84-94.
(收稿日期:2020-10-13)
