苏姜猪SIRT3 基因多态性及其组织表达定位分析
2020-12-21陈章言黄欣爱
陈章言,王 洁,黄欣爱,张 伟*
(1.江苏农牧科技职业学院,江苏泰州 225300;2.江苏姜曲海种猪场,江苏泰州 225300)
沉默信息调节因子(Silence Information Regulator,Sirtuin)是一类高度保守的烟酰胺腺嘌呤二核苷酸(Nicotinamide Adenine Dinucleotide,NAD+)依赖的去乙酰化酶,在动物进化过程中其高度保守,自细菌到人类的生物机体内均存在其同源体。Sirtuins家族共包含7 个成员(SIRT1~SIRT7),它们具有不同的亚细胞定位和功能[1-2]。SIRT3 主要定位于生物细胞线粒体中,通过与2 个关键的去乙酰化底物——线粒体超氧化物歧化酶(SOD2)和异柠檬酸脱氢酶2(IDH2)结合从而使相关的蛋白质脱乙酰基,以增加活性氧自由基(ROS)清除酶活性,抑制ROS 蓄积,调节ATP 生成速率,稳定线粒体功能[3-4],进而参与调控动物机体的三羧酸循环、尿素循环以及脂肪酸β-氧化等代谢活动过程,因此SIRT3在动物机体的能量代谢和氧化应激过程中具有重要的作用[5-6]。然而,相关研究发现,SIRT3基因序列存在多态性,这种多态性与人(Homo saplens)的衰老和寿命的调控以及代谢综合症紧密关联[7-9]。而针对家畜生产性能,有研究表明秦川牛(Bos taurus)SIRT3基因多态性与背膘厚、眼肌面积和肌内脂肪含量显著相关[10];在大围子猪SIRT3基因的第5 外显子上存在一处A/G 突变位点,形成AA、AG 和GG 3 种基因型,AA 基因型的24 h 滴水损失、失水率和肌内脂肪含量与AG、GG 基因型相比差异显著,而湘村黑猪仅有肌肉色值中的亮度值在AG 和GG 基因型之间存在显著性差异[11]。截至目前,有关猪组织脏器中SIRT3基因表达、分布定位的研究较少。本研究以苏姜猪(Sus scrofa)为研究对象,检测SIRT3基因的多态性与屠宰性状和肉质性状的相关性,并对野生型(CC)公母苏姜猪组织中SIRT3基因表达分布进行分析,以期为苏姜猪屠宰和肉质性状的分子标记辅助选育提供依据,为丰富猪的SIRT3基因研究提供参考。
1 材料与方法
1.1 实验材料及设计 实验用苏姜猪均来自江苏姜曲海种猪场。采集60 头成年苏姜猪的耳组织样,-80℃保存,用于DNA 提取及SIRT3基因多态性检测。
根据SIRT3基因多态性检测结果,随机选取日龄相同(205 日龄)、健康无病的CC 型和CT 型成年苏姜公猪(去势)各3 头,进行屠宰,测定屠宰性状(眼肌面积、屠宰率、背膘厚、前区瘦肉率、中区瘦肉率、后区瘦肉率)及肉品质指标(pH、剪切力、系水力);采集野生型即纯合型(CC)成年苏姜公猪(去势)和母猪各3 头的心脏、肝脏、脾脏、肺脏、肾脏、胃、大肠、小肠、腹脂、背最长肌、腿肌、卵巢(母猪)、输卵管(母猪)、子宫角(母猪)14 个组织样,每个组织3 个重复,分装在1.5 mL Eppendorf 管中,贮存于液氮中,用于实时荧光定量检测;同时,再采集所有上述14 个脏器组织样置于10%中性福尔马林溶液中固定,用于免疫组织化学检测。
1.2SIRT3基因多态性检测 根据NCBI 中SIRT3基因序列信息(NC_010444.4:c55074-38465),使用Primer3在线软件和Oligo 6.31 设计引物(表1)。将苏姜猪耳组织样送上海翼和应用生物技术有限公司进行SIRT3基因多重PCR 高通量测序。
一轮PCR 扩增总体系为10 μL:ddH2O 3.1 μL、10×Buffer 1 μL、Prime(r50 nmol/L)2 μL、dNTP(2.5 mmol/L)0.8 μL、酶(5 U/μL)0.1 μL、Sample 2 μL、Mg2+(100 mM)1 μL。PCR 程序为:95℃15 min;94℃30 s,60℃10 min,72℃30 s,4 个循环;94℃30 s,60℃ 1 min,72℃30 s,20 个循环。二轮PCR 扩增总体系为21 μL:ddH2O 3.5 μL、10×Buffer 2 μL、Barcode(2 μmol/L)3.6 μL、dNTP(2.5 mmol/L)0.8 μL、酶(5U/μL)0.1 μL、Sample 10 μL、Mg2+(100 mmol/L)1 μL。PCR 程序为:95℃ 15 min;94℃ 30 s,60℃ 4 min,72℃ 30 s,5 个循环;94℃ 30 s,65℃ 1 min,72℃ 30 s,10 个循环。

表1 SIRT3 基因多态性检测引物序列及扩增区域
1.3 实时荧光定量PCR 根据GenBank 中猪SIRT3(登录号:NM_001110057.1)基因序列和β-actin(登录号:KU672525.1)作为内参基因,采用Primer Premier 6.0和Beacon designer 7.8 软件设计实时荧光定量PCR 引物(表2),由上海英骏生物技术有限公司合成。根据说明书,使用TRIzol(TaKaRa)提取所有受试猪各组织样品中的总RNA。取1 000 ng 总RNA,以20 μL 反应体系反转录成cDNA。实时荧光定量PCR 反应体系20 μL:Power SYBR®Green Master Mix 10.0 μL、Forward Primer(10 μmol/L) 0.5μL、Reverse Primer (10 μmol/L)0.5 μL、cDNA 1.0 μL。反应条件为:95 ℃ 1 min;40个循环(95℃ 15 s,63℃ 25 s),每个样品重复3 次。利用2-ΔΔCT法计算各基因的相对表达量。

表2 实时荧光定量PCR 检测用引物序列及扩增产物长度
1.4 免疫组织化学染色 将组织样固定于10%中性福尔马林溶液,经常规石蜡包埋,5 μm 厚切片。石蜡切片经脱蜡和水化处理后,用Tris-EDTA(pH=9)抗原修复,微波加热10 min,自然冷却至室温后,TBS 洗涤3 次,用3% H2O2封闭内源性过氧化物酶之后以5% BSA 于37℃封闭30 min,弃掉封闭液,使用SIRT3 抗体(CST,1:250)作为一抗4℃孵育过夜。使用HRP 标记的羊抗兔IgG(南京生兴生物技术有限公司,sn134,1:100)作为二抗识别特异性一抗,DAB 显色液显色,之后,以苏木精复染细胞核后,梯度酒精脱水,中性树胶封片。
1.5 统计分析 各组织中SIRT3mRNA 相对表达量以β-actin作为内参基因,矫正不可控制因素,归一化起始组织量,并定义心脏组织中的表达水平为1,以对SIRT3mRNA 在不同组织中的表达水平进行相对定量。所有数据以平均值±标准差表示,使用SPSS 19.0 软件的单因素方差分析(ANOVA)、独立样本t检验和χ2检验进行差异显著性和群体遗传平衡性分析,P<0.05表示差异显著。
2 结果
2.1 苏姜猪SIRT3基因多态性及群体遗传分析
2.1.1 苏姜猪SIRT3基因多态性检测 通过对60 头苏姜猪耳组织样进行SIRT3基因重测序检测,分别在外显子1 区的314 位点(T →A)和内含子6 区的14 589位点(C →T)存在碱基突变(图1)。将外显子1 处的突变位点与GenBank 中猪XM_005652976.3 序列比对,结果显示这一碱基位点位于5’非翻译区(5’UTR)。
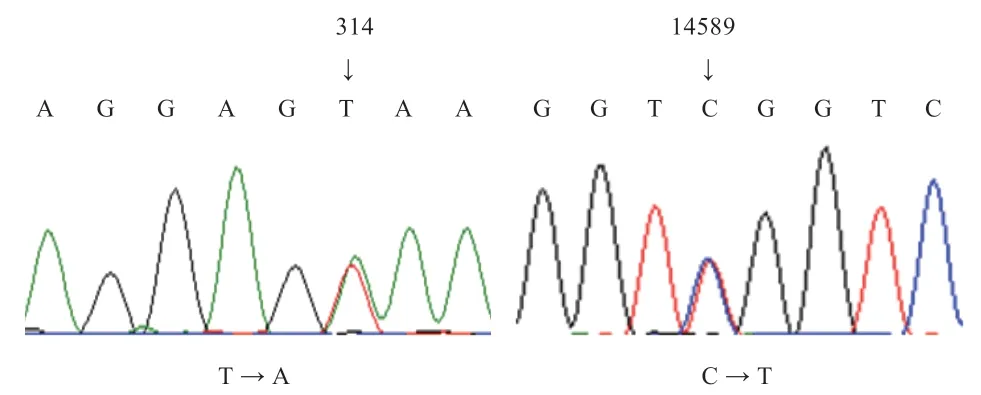
图1 苏姜猪SIRT3 基因碱基突变位点
2.1.2 苏姜猪SIRT3基因多态性群体遗传分析 由表3 可以看出,在所检测的60 头苏姜猪群体中,SIRT3基因314 位点仅检测出1 头苏姜猪为杂合型TA,其他苏姜猪均为纯合型TT;14 589 位点检测到纯合型CC(48 头)和杂合型CT(12 头)个体。2 个多态位点的He 均较低,PIC 值均小于0.25,为低度多态,表明该群体遗传变异不高。经χ2检验,这2 个碱基突变位点在所选择的苏姜猪群体中均处于哈迪-温伯格遗传平衡(P>0.05)。

表3 苏姜猪群体中SIRT3 基因多态分布
2.2 苏姜猪SIRT3多态性与屠宰性能及肉品质间相关性分析 因SIRT3基因314 位点仅检测出1 头苏姜猪为杂合型TA,故本研究未进行该位点不同基因型与屠宰性能和肉品质间相关性分析,只进行SIRT3基因14 589 位点纯合型CC(48 头)和杂合型CT(12 头)苏姜猪屠宰性能和肉品质间相关分析。
由表4 可以看出,6 项屠宰指标在纯合型(CC)和杂合型(CT)苏姜猪公猪间差异均不显著,表明2种基因型苏姜猪的产肉性能无明显差别;在肉品质指标中,纯合型(CC)苏姜猪肉的剪切力显著高于杂合型(CT),表明杂合型苏姜猪猪肉嫩于纯合型,然而系水力和pH 则在不同基因型苏姜猪肉间差异不显著。

表4 苏姜猪SIRT3 基因14 589 位点不同基因型屠宰性状和肉品质指标
2.3 苏姜猪组织中SIRT3mRNA 表达量分析 因CC型和CT 型苏姜猪在产肉性能方面无显著差别,因此不进行这2 种基因型苏姜猪组织中SIRT3mRNA 表达量检测分析,只进行野生型(CC)公母苏姜猪组织中SIRT3mRNA 表达量差异检测分析。由图2 可知,在公猪的11 个组织和母猪的14 个组织中均存在SIRT3mRNA 表达。经统计分析,SIRT3mRNA 在公猪的胃组织中表达量相对最高,显著高于心脏、肝脏、大肠、背最长肌、腿肌组织;母猪则肝脏组织中的表达量相对最高,显著高于除心脏、大肠、背最长肌、胃组织外的其他组织。其中,公母猪的脾脏组织中SIRT3mRNA表达量均相对最低。除肝脏和脾脏组织外,公猪各组织中SIRT3mRNA 相对表达量均较高于母猪,其中,公猪的肾脏和胃组织中表达量显著高于母猪。
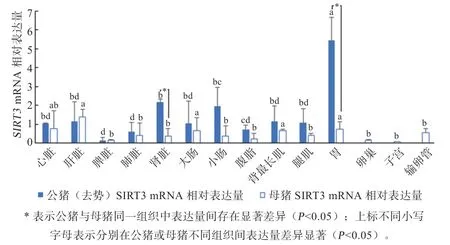
图2 苏姜猪不同组织脏器中SIRT3 mRNA 相对表达量
2.4 SIRT3 蛋白在苏姜猪组织中的定位分布 经免疫组织化学检测,在苏姜猪各脏器组织中均检测到SIRT3免疫阳性颗粒。由图3 可知,SIRT3 免疫阳性颗粒主要分布于各组织细胞的细胞质中。在肌肉组织(心肌、腿肌和背最长肌)的肌细胞、肝脏的肝细胞、肾脏的近曲小管和远曲小管细胞、肺脏组织的肺泡细胞以及细支气管粘膜层细胞中有较丰富的SIRT3 免疫阳性颗粒分布;脾脏组织中SIRT3 免疫阳性颗粒分布较少,仅在红髓细胞中有分布;在消化系统的胃、小肠和大肠组织中,SIRT3 免疫阳性颗粒主要分布于黏膜层上皮细胞及固有层腺细胞中;在母猪生殖系统中,SIRT3 免疫阳性颗粒主要分布于卵巢卵泡的颗粒细胞和卵母细胞,输卵管和子宫组织的粘膜层上皮细胞中。


图3 苏姜猪组织中SIRT3 免疫阳性颗粒分布
3 讨 论
SIRT3 为NAD+依赖的去乙酰化酶,通过使蛋白质被乙酰化修饰参与机体的三羧酸循环、尿素循环、脂肪酸β-氧化等代谢过程,在动物机体物质代谢和能量代谢中具有重要的作用。有研究报道表明,SIRT3基因存在碱基突变,这种突变会影响秦川牛和湘村黑猪的产肉性能[10-11]。本研究在苏姜猪SIRT3基因的外显子1 和内含子6 区分别检测到T →A 和C →T 的碱基突变,而与大围子猪和湘村黑猪的研究结果[11]不同,未在第5 外显子区检测到碱基突变。本研究结果显示,仅纯合型(CC)苏姜猪肉的剪切力显著高于杂合型(CT),其他屠宰指标及肉品质指标均无显著差异,表明杂合型(CT)苏姜猪的肉质嫩于纯合型(CC),可作为后续苏姜猪肉质进一步选育提高的候选基因。
SIRT3在猪组织中的表达具有广谱性,如通城猪、湘村黑猪和大围子猪的心脏、肝脏、脾脏、肺脏、肾脏、肌肉等组织中均检测到SIRT3mRNA 的表达[11-12]。本研究在苏姜猪的心脏、肝脏、脾脏、肺脏、肾脏、胃、大肠、小肠、腹脂、背最长肌、腿肌、卵巢、输卵管、子宫等14 个组织中均检测到SIRT3mRNA 的表达,其中,在胃、心脏、腿肌、背最长肌和肝脏组织中的表达量相对较丰富,脾脏组织中SIRT3mRNA 的表达量相对最低,这与相关报道中提出的在动物代谢旺盛的组织如肌肉、肝脏、心脏中有较高的表达结果相同[5-6]。对比公母苏姜猪各组织中SIRT3mRNA 表达量差异,结果显示除苏姜母猪的肝脏和脾脏组织中SIRT3mRNA表达量稍高于公猪外,其他组织中的表达量均相对低于公猪,特别是在母猪胃和肾脏组织中的表达量显著低于公猪,表明SIRT3mRNA 在动物机体组织中的表达量与性别有关。
SIRT3 蛋白主要定位于动物机体组织细胞质的线粒体中,其表达对于ATP 的生成具有重要作用[4,11]。如在肝脏组织中,SIRT3 参与动物尿素循环和脂肪酸的合成与分解等代谢活动,通过去乙酰化增强限速酶K88 活性,进一步增强尿素循环,而在能量限制或饥饿时,SIRT3可以在线粒体中直接脱掉OTC 上的乙酰基,上调其活性,增加尿素生成[13]。特异性敲除肝脏SIRT3基因的小鼠体内,发现脂肪酸氧化中间产物和甘油三酯显著升高,LCAD(Long-chain acy1 CoA dehydrogenase) 第42 号赖氨酸呈现高度乙酰化,酶活性显著降低,最终导致ATP 水平下降,进一步促进肝脏中脂肪沉积,使肝细胞的死亡速度加快[14]。而肌肉组织中缺失SIRT3的小鼠则表现出耗氧量减少,并出现氧化应激,导致胰岛素信号受损,心脏、肾脏和肝脏中的ATP 水平下降幅度超过50%[15]。本研究通过免疫组织化学方法在苏姜猪14 个脏器组织的细胞中均检测到SIRT3 免疫阳性颗粒,特别是在SIRT3mRNA 表达量相对较丰富的心脏、胃、肌肉、肝脏和肾脏组织细胞质中有丰富的SIRT3 免疫阳性颗粒。这些组织中ATP 水平较高,SIRT3 可通过调节代谢酶来决定ATP 的生成速率[5-6,11]。在苏姜猪肾脏组织中,SIRT3 蛋白主要分布于需要较多能量进行吸收和分泌功能的近曲小管和远曲小管细胞中,而在具有血液过滤器作用的肾小球中免疫阳性颗粒较少。动物消化器官的消化腺细胞从其周围的血液中摄取原料,合成分泌物后贮存或在受到适宜刺激时排出分泌物,这一分泌过程为主动活动过程,整个分泌过程需要消耗能量,这些能量主要来自腺细胞内的ATP。本研究在苏姜猪消化系统的胃、小肠和大肠组织的腺细胞中均检测到丰富的SIRT3 免疫阳性颗粒,表明SIRT3与细胞的分泌功能有关。在苏姜猪母猪生殖系统组织中,SIRT3 免疫阳性颗粒主要分布于卵巢的卵泡颗粒细胞、卵母细胞和生殖道的输卵管、子宫组织的粘膜层上皮细胞中,这表明SIRT3参与卵子的成熟过程,且与生殖道黏膜层细胞的分泌活动有关联。
4 结 论
本研究发现,苏姜猪SIRT3基因在外显子1 区和内含子6 区分别存在1 处碱基突变,其中内含子6 区的碱基突变显著影响苏姜猪猪肉的剪切力,表明SIRT3基因为进一步提高苏姜猪肉品质的候选基因。SIRT3mRNA 在苏姜猪脾脏组织中的表达量相对最低,在检测的器官组织中,除肝脏和脾脏外公猪的表达量相对高于母猪;SIRT3 免疫阳性颗粒主要分布于苏姜猪的肝脏细胞、肌肉细胞和黏膜层细胞,在母猪卵巢颗粒细胞和卵母细胞中也有丰富表达,表明SIRT3主要参与动物机体细胞的物质、能量代谢以及卵子的成熟过程。
