Residual force enhancement due to active muscle lengthening allows similar reductions in neuromuscular activation during position-and force-control tasks
2020-12-18RhiannonMarionGeoffreyPower
Rhiannon Marion,Geoffrey A.Power*
Department of Human Health and Nutritional Sciences,College of Biological Sciences,University of Guelph,Guelph,Ontario,N1G 2W1,Canada
Abstract Background:Residual torque enhancement(rTE)is the increase in torque observed during the isometric steady state following active muscle lengthening when compared with a fixed-end isometric contraction at the same muscle length and level of neuromuscular activation.In the rTE state,owing to an elevated contribution of passive force to total force production,less active force is required,and there is a subsequent reduction in activation.In vivo studies of rTE reporting an activation reduction are often performed using a dynamometer,where participants contract against a rigid restraint,resisting a torque motor.rTE has yet to be investigated during a position task,which involves the displacement of an inertial load with positional control.Methods:A total of 12 participants(6 males,6 females;age=22.8±1.1 years,height=174.7±8.6 cm,mass=82.1±37.7 kg;mean±SD)completed torque-and position-matching tasks at 60%maximum voluntary contraction for a fixed-end isometric contraction and an isometric contraction following active lengthening of the ankle dorsi flexors.Results:There were no significant differences in activation between torque-and position-matching tasks(p=0.743),with~27%activation reduction following active lengthening for both task types(p<0.001).Conclusion:These results indicate that rTE is a feature of voluntary,position-controlled contractions.These findings support and extend previous findings of isometric torque-control conditions to position-controlled contractions that represent different tasks of daily living.
Keywords:Eccentric;Electromyography;History dependence offorce;Position tasks;Residual force enhancement
1.Introduction
The contractile history of a muscle,termed the“history dependence offorce”,is known to increase a muscle's forceproducing capacity following active lengthening(residual torque enhancement;rTE)compared with a fixed-end isometric contraction at the same muscle length and level of activation.1Following active muscle lengthening,a given submaximal isometric force can be achieved with less neuromuscular activation than before the lengthening stimulus.2-4This activation reduction during submaximalcontractionsrangesfrom 7%-25%2and has been observed across multiple muscles and muscle groups,such as the adductor policis brevis,3,5,6tibialis anterior,4,7plantar flexors,8-10knee extensors,11and elbow flexors.12Some studies have reported differences in the control of isometric force between conditions that require the subject either to hold a rigid load or to hold an equivalent inertial load in the same anatomical position(i.e.,position task)but in free space.13We were interested in investigating how this taskdependency may carry over to isometric tasks following active lengthening.Despite marked differences in neuromuscular activation during these tasks,13the implications of rTE during position tasks has not yet been explored.
The history dependence offorce was first observed by Abbott and Aubert,1and since then it has been observed from the single sarcomere,14at the single- fiber level in humans,15and up toin vivostudies of human voluntary muscle contraction.2,16Despite these observations,the mechanisms of the history dependence offorce are still under debate,and the implications on voluntary control offorce are not fully understood.The magnitude of rTE has been shown to be dependent on the amplitude of muscle lengthening and is thought to involve 2 components:a cross-bridge-based component17and a passive component related to the giant molecular spring,titin.Titin contributes to an increase in passive force following active lengthening.18,19Following active lengthening,in the rTE state,an increased contribution from passive force to total force allows for reduced neuromuscular activation(i.e.,activation reduction)through less motor unit recruitment and a lower firing rate.2,4,5,20,21
In vivostudies of activation reduction in the rTE state use a dynamometer to control the joint excursion and presumed rate of stretching that a muscle is subjected to during the protocol.The participant contracts against a rigid restraint,receiving torque feedback in order to match torque between the rTE and isometric states.To date,rTE has not been studied during a position task that requires the participant to hold an inertial load in the same anatomical position as a rigid load(i.e.,position-matching task)but with unconstrained movement.Position-matching tasks are known to differ from torque-matching tasks in activation strategies of the motor neuron pool22-25in that position-matching tasks are associated with decreased presynaptic inhibition23and increased muscle spindle sensitivity compared with torquematching tasks.24During eccentric contractions,there is a distinction to be made between the 2 separate actions of displacing a load during a position-matching task and resisting a torque motor in a torque-matching task,26which lead to different control strategies during dynamic contractions.27,28During eccentric contractions,lowering an inertial load involves a reduction in corticospinal excitability and a reduction in motor unit discharge rates compared with resisting a torque motor.26It has been proposed that increased gain in stretch re flex is,in part,responsible for these differences in motor unit discharge rates.27
Therefore,the purpose of the present study was to determine whether activation reduction is present following active lengthening of the ankle dorsi flexors for a position-matching task.Based on reports of rTE and activation reduction in previousin vivostudies,we hypothesized that activation reduction would still occur during position-matching tasks.
2.Methods
A total of 14 healthy adults(7 males,7 females;age=23.4±2.6 years;height=175.5±8.3 cm;mass=81.9±35.1 kg;mean±SD)participated in a single data-collection session.Participants were free of neurological,orthopedic,or cardiovascular conditions.Participants gave written informed consent prior to testing.All procedures involving human subjects were approved by the Research Ethics Board of the University of Guelph(15NV008)and,with the exception of registration in a database,conformed to the Declaration of Helsinki.
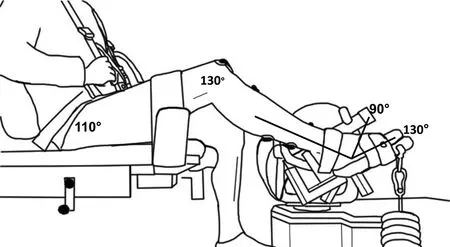
Fig.1.Depiction of the experimental set-up on the HUMAC NORM dynamometer.Participants sat on the dynamometer with a hip angle of 110°and a knee and ankle angle of 130°.Active lengthening occurred through a 40°range of motion about the ankle joint.For position tasks,a load equivalent to 60%maximum voluntary contraction at a joint angle of 130°,corresponding to 40°plantar flexion,was hung from the dynamometer foot plate.
2.1.Experimental arrangement
Participants were seated in a HUMAC NORM dynamometer(CSMi Medical Solutions,Stoughton,MA,USA),which recorded all torque,angular velocity,and position(joint angle)data.Participants were seated with a hip angle of 110°,a knee angle of 130°(relative to 180°as full extension),and an ankle angle of 130°of plantar flexion(from 90°as neutral).The knee was supported by a padded cushion and immobilized by a strap.The right foot was fixed to the dynamometer foot plate with one strap placed over the ankle and one strap placed at the mid-distal portion of the metatarsals(for experimental set up,see Fig.1).
Electromyography(EMG)was recorded from the tibialis anterior(TA)and soleus muscles using silver-silver chloride(Ag/AgCl)electrodes(1.5 cm×1.0 cm)(Kendall,Mans field,MA,USA).Prior to electrode placement,the skin was shaved and cleaned with alcohol.A common ground electrode was placed on the patella;an electrode was placed 7 cm inferior,2 cm lateral to the tibial tuberosity over the muscle belly of the TA near the TA motor point;and an electrode was placed distally over the tendon of the TA with an inter-electrode distance of~8 cm.The tendon of the TA was located by palpating the ankle and instructing participants to dorsi flex against the strap.A large electrode distance was used to optimize evoked potentials.Two more electrodes were placed on the soleus,one being placed 2 cm inferior to the midline of the gastrocnemii and one being placed on the calcaneal tendon.
Torque,angular position,and stimulus trigger data were sampled at 1000 Hz using a 12-bit analog-to-digital converter(PowerLab System 16/35;ADInstruments,Bella Vista,Australia).EMG data were sampled at 2000 Hz and bandpass- filtered online between 10 Hz and 1000 Hz.All data were analyzed using Labchart(Labchart,Pro Modules 2014,Version 8;ADInstruments)software.
2.2.Experimental protocol
In order to normalize EMG root mean square(EMGRMS),maximal compound action potentials(M-max)were recorded from the TA and soleus muscles by stimulating the deep fibular and tibial nerves,respectively.A standard 30-mm clinical bar electrode(Chalgren Enterprises,Gilroy,CA,USA)was placed over the nerve,and stimuli were delivered with a single pulse from a constant-current high-voltage stimulator(Model DS7AH,Digitimer,Welwyn Garden City,Hertfordshire,UK).Voltage was set to 400 V and pulse width to 200 μs.Current was increased incrementally until a plateau in peak-to-peak M-wave amplitude was reached.To ensure consistent activation,this current was increased to 120%of the current required to generate M-max.
Participants performed up to 3(a minimum of 2)maximum isometric voluntary contractions(MVC)and were instructed to perform ankle dorsi flexion as hard and as fast as possible.For each MVC,participants received visual feedback,provided as a torque trace on a computer monitor,as well as strong verbal encouragement from the researcher.Each MVC was separated by 5 min of rest.The first MVC was used as a practice trial.Subsequent MVCs involved the use of the interpolated twitch technique(ITT)to ensure≥95%voluntary activation(VA).29All participants were capable of≥ 95%VA.After≥ 95%VA was achieved,participants performed a 10-s maximum effort fixed-end isometric contraction.Torque was obtained and averaged from 8.0 s to 8.5 s of this contraction.This time point was used because it corresponded to the isometric steady state of the rTE contractions.A total of 60%of this value was used as the target load for both the position-and torque-matching tasks.All tasks were completed twice,with 5 min of rest in between each task.All participants were familiar with these experiments and had participated in similar experiments before;therefore,we did not expect an ordered learning effect.The order of contractions was chosen to minimize the effects offatigue and bias our results toward lower rTE values.The isometric contraction was performed first,followed by the rTE contraction.With this approach,we can be fully con fident that any effect of active lengthening on subsequent activation reduction was,indeed,due to the history-dependent properties of the muscle and not to some extraneous factor.
The position-matching tasks were performed first to ensure the load would be matched during the isometric steady state of both the position and torque tasks.With the foot plate set to 130°plantar flexion,a load consisting of 0.1 kg,0.5 kg,and 1.0 kg plates was hung from the foot plate until 60%MVC was reached.Once 60%MVC load was attached to the foot plate,the dynamometer's angular velocity was unrestricted to ensure free movement,and the range of motion was increased from approximately 85°to 133°to ensure that the participant could lower to 130°unhindered.Because the load was suspended from the dynamometer,and the participant was actively holding the load,the torque value,which the load cell registers,would essentially be 0 N⋅m.Therefore,during the position tasks,the participant matched angular position.The participant was given position feedback for both the isometric position-matching task and the rTE position-matching task as a prescribed angle shown as a line on the computer monitor.For the isometric task,the participant's foot was moved to 130°,and the participant was instructed to “take the weight”to initiate contraction,holding the position at the joint angle that matched the position guideline.For the rTE position-matching task,the researcher moved the participant's foot to 90°,and the participant was instructed to “take the weight”to initiate contraction in the 90°position and then carefully control the lowering of the foot to the 130°guideline over 3 s.It should be noted that,because the weight stack was hung from the foot plate,the moment arm and,thus,the load imposed on the ankle dorsi flexors would vary throughout the range of motion during the active lengthening contraction.To obtain an isometric steady state,the participant held this contraction for at least 5 s following the active lengthening.
Torque-matching tasks were performed following positionmatching tasks.The participant received torque feedback during the isometric and rTE torque tasks.To ensure tasks were controlled for torque within-participants,the torque-matching guidelines were matched to the inertial load used during the position task.During the isometric torque task,the foot plate was set to an ankle angle of 130°.The participant was instructed to match the torque guideline for 10 s.During the rTE torque task,the participant initiated the contraction at 90°and actively lengthened,at 13°/s over ~3 s,to 130°,and was instructed to maintain a constant effort while matching the target torque guideline.The participant held the contraction for 5 s thereafter.We make the assumption that torque reaches an isometric steady state following the dissipation of torque transients.Therefore,for both position and torque tasks,torque and EMG data from the tibialis anterior and soleus muscles were taken from 8.0 s to 8.5 s and averaged for data analysis.
Following 2 trials for each task,the participant performed a final MVC with ITT to ensure that the participant was not fatigued and was capable of near maximal activation.For a depiction of the experiment's time line,see Fig.2.
2.3.Data and statistical analyses
Of 14 participants tested,12 were used in analysis.Two participants were excluded because the post-MVC was reduced>15%,indicating fatigue.The 15%value was chosen as a cut-off point owing to apost hocanalysis of torque declines following the task.Indeed,there was some variability,but most participants declined<10%,and only 2 participants declined>15%.Of the 12 participants used in analysis(6 males,6 females;age=22.8±1.1 years,height=174.7±8.6 cm,mass=82.1±37.7 kg;mean±SD),a single trial that best matched 60%MVC(e.g.,<±5%deviation from the target trace)from each task was used in analysis.Position trials were not used if the participant's ankle angle movement went past the target ankle angle;this limited corrective muscle shortening to avoid any confounding factors of shortening-induced torque depression(for a review,see Chen et al.30)on rTE.Time-matched data during the isometric steady state of contraction,between 8.0 s and 8.5 s or 9.0 s and 9.5 s,was analyzed for mean torque,mean TA,and soleus EMGRMS.Activation reduction of the TA was calculated as a percent from EMGRMSas follows:
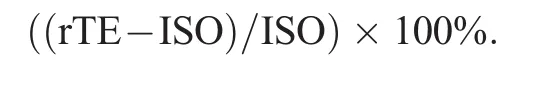
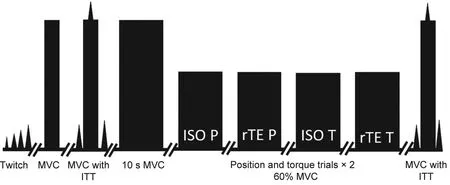
Fig.2.Depiction of the experiment's time line.The protocol began with twitch determination so that MVCs could be completed with the interpolated twitch technique.One 10-s MVC was completed,where 60%MVC was taken from the 8.0 s to 8.5 s steady-state time epoch.All tasks were performed twice at an intensity of 60%MVC with 3 min of rest,culminating with a final MVC to monitor any strength loss throughout the protocol.ISO=isometric contraction;ITT=interpolated twitch technique;MVC=maximum voluntary contraction;P=position matching task;rTE=residual torque enhancement;T=torque matching task.
Neuromuscular economy was also used to assess torque per unit of EMGRMSduring rTE tasks.This measure eliminates differences and variability in the EMG values that may occur due to difficulties in matching torque.3Antagonist coactivation was determined by normalizing soleus EMG during dorsi flexion to the resting Mmax;this value was then divided into the normalized agonist activation to account for the presumed greater activation(and cross-talk)in the isometric conditionvs.the rTE condition.All data were analyzed using Labchart(Labchart,Pro Modules 2014,Version 8;ADInstruments),Excel(Microsoft,Redmond,WA,USA),and SPSS Version 26.0 software(IBM Corp.,Armonk,NY,USA).ttests were used to analyze differences in strength,as well as voluntary activation pre-and postprotocol.There are no reports of a sex difference for the history-dependence offorce;however,a one-way analysis of variance(ANOVA)was used to assess any differences owing to sex,before collapsing across sex.A two-way ANOVA with repeated measures was used to assess differences in activation reduction,neuromuscular economy,and soleus coactivation(history dependence×task).Effect size was calculated as partial eta squared().All data in the text are reported as mean ±SD and are reported in the figures as mean±SEM.Significance was set top≤0.05.For a depiction of one participant's raw data traces,see Fig.3.
3.Results
3.1.Peak torque and voluntary activation
Voluntary activation was not different between males(99.6%±0.9%)and females(99.2%±0.9%)(p=0.157)during the initial MVC(99.4%±0.9%)and final MVC(99.3%±0.8%)(p=0.659).Males were stronger than females(p=0.050).There was no difference(p=0.926)in torque generation from the initial to final MVC for males(24.6 ± 7.5 N⋅m to 20.9± 6.7 N⋅m)and females(17.3±2.0 N⋅m to 20.7± 5.5 N⋅m).
3.2.Activation reduction and neuromuscular economy
Activation reduction did not differ between males and females(p=0.113,=0.232)and was,thus,collapsed across sex.There was no interaction between history dependence and task(p=0.882,=0.002).However,there was a main effect of activation reduction between isometric and rTE states in that activation reduction was present in both the position-matching(26.9%±14.3%)and torque-matching(27.1%±9.4%)tasks(p< 0.001,=0.704);however,the magnitude of activation reduction did not differ between the position-and torque-matching tasks(p=0.743,=0.010)(Fig.4).For neuromuscular economy,there was no interaction between history dependence and task(p=0.199,=0.145).Neuromuscular economy increased similarly for both tasks(p=0.047,=0.312),from 2.8±1.5 during the isometric to 3.4±2.1 in the rTE state for the position tasks,and from 2.9±1.2 during the isometric to 3.4±1.5 in the rTE state during the torque tasks,with no difference observed between tasks(p=0.971,=0.000).
3.3.Soleus coactivation
For antagonist coactivation,there was no interaction between history dependence and task(p=0.892,=0.000).There was an effect of history dependence(p=0.006,=0.048),in that soleus coactivation was increased from 22.6%±11.6%in isometric to 29.0%±17.2%in the rTE state during the position tasks and from 21.0%±10.4%in isometric to 26.4%±14.4%in the rTE state during the torque tasks.There was no significant difference between tasks(p=0.591,=0.007).
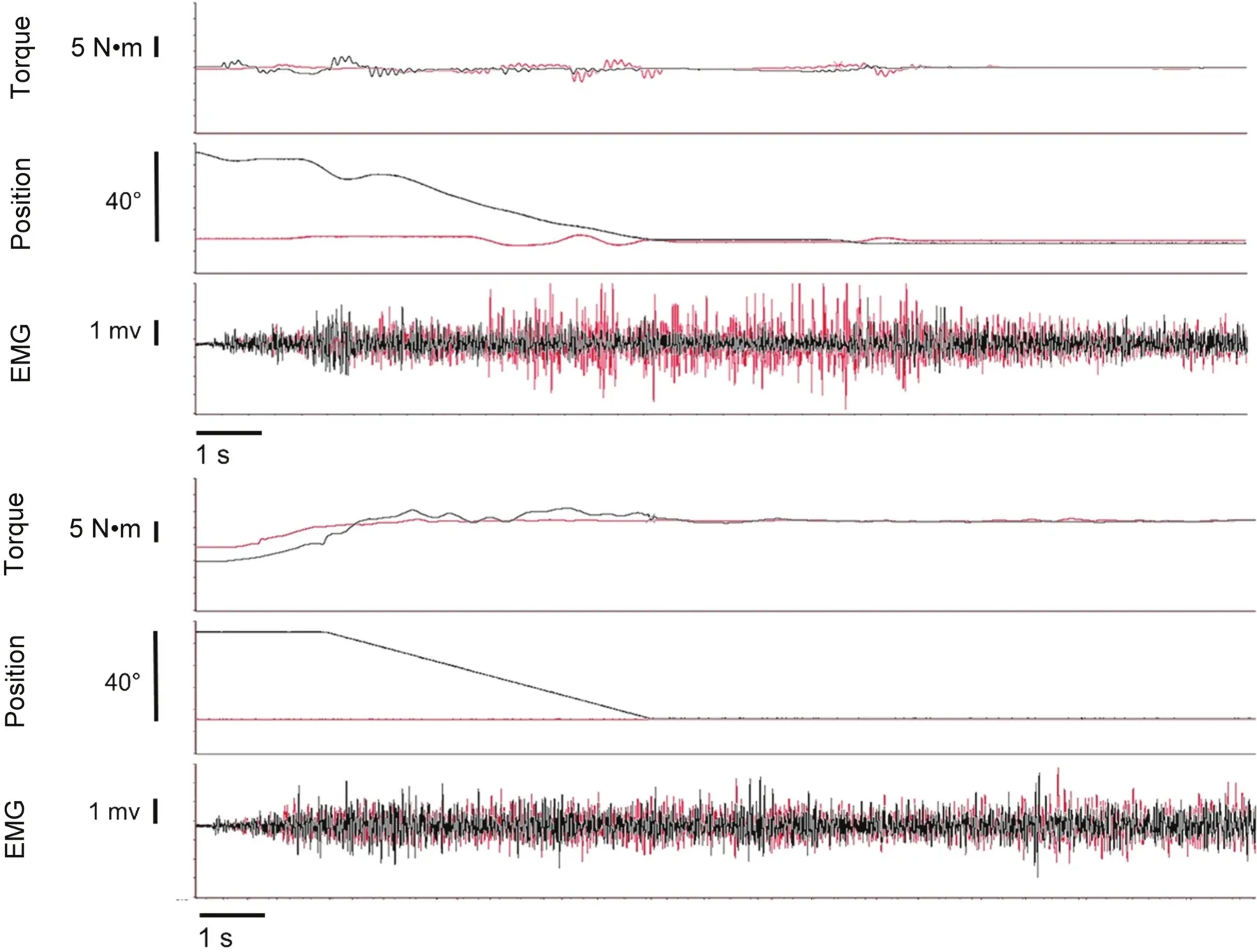
Fig.3.Raw data traces displaying torque,position,and electromyography(EMG)for position(top)and torque(bottom)tasks.Residual torque enhancement(rTE)is overlaid in black over fixed-end isometric conditions in red.rTE conditions involved an isometric contraction,followed by active lengthening to a subsequent isometric contraction.Please note that other than some minor adjustments,the load during the position-matching task registers as 0,owing to the participant's holding and matching the load.

Fig.4.Activation reduction during position and torque tasks.Bars represent the mean and standard error.Individual circles represent individual participants.Activation was reduced in both position and torque tasks(*p<0.001)but was not different between tasks(p=0.743).
4.Discussion
The purpose of the present study was to determine the relevance of rTE as it relates to a position-matching task with the use of an inertial load in the same anatomic position(i.e.,position task)as compared with a static rigid load.Our hypothesis was con firmed,because activation reduction occurred in both tasks,with no differences observed between the position-and torque-matching tasks.There was a~27%reduction in EMG activity between the isometric contraction and isometric steady states following active lengthening for both position tasks and torque tasks.Thus,rTE is a feature of controlling a positional contraction(i.e.,position-matching task),which supports and extends previous findings of isometric torque control conditions to position matching that represent various tasks of daily living.
When studying the effects of prior lengthening contractions on torque,an activation or torque-matching task may be imposed,resulting in a subsequent increase in torque or reduction in activation,respectively.Thus,activation reduction can serve as a proxy for rTE.Activation reduction observed in this study is not different from rTE(range:7%-25%)observed in previous studies of the ankle dorsi flexors.Power and others31,32observed 7%-25%and 10%-25%rTE during maximal dorsi flexion contractions.Previous studies of rTE in the ankle dorsi flexors found a 7%-15%increase in torque following active lengthening.8,33Neuromuscular economy,the amount of torque produced per unit of EMG,is a measure that accounts for variability in torque and EMG when attempting to match a target torque.In both position and torque tasks,neuromuscular economy was enhanced in the rTE state following active lengthening,indicating that less neuromuscular activity was required to produce the same amount of torque.
In the present study,all participants displayed rTE,indicating that our participant pool had no nonresponders.When investigating rTE in humans,there is often an incidence of nonresponders,or participants who do not exhibit increased torque following active lengthening.Our lack of nonresponders could,in part,be due to our study design.Our study involved a high load of 60%MVC,and it appears that the incidence of nonresponders is reduced with increasing contraction intensity.4,5,16However,the mechanism for increased responders during higher contraction intensities is currently not fully understood.2Another factor that may affect the responder/nonresponder phenomenon is the magnitude of active muscle fiber lengthening.The range of motion of 40°in our study was likely large enough to induce sufficient active muscle fiber lengthening and take advantage of the passive component relating to rTE.1,2,16An important consideration when investigating rTE in humans is controlling for the amount of muscle fiber versus tendon lengthening,which is ultimately governed by the amount of in-series compliance of the musculotendinous unit.34,35This,in turn,affects the magnitude of active muscle lengthening during the protocol by each participant.By having a large range of motion,some differences in tendon compliance that may affect rTE can,it is hoped,be attenuated.
During the position task,because lowering the load was completed by the participant,angular velocity was at times variable,while terminal angle was consistent across participants.It should also be noted that the load imposed on the ankle dorsi flexors varied throughout the range of motion owing to a changing moment arm during the active lengthening contractions.This variability is,in part,a limitation,but it is also an aspect of task differences that can occur with position-controlled movement,and it adds further ecological validity to the relevance of rTE during position-controlled tasks.Despite some potential variability in torque during the eccentric phase of the tasks,there were no significant differences in activation reduction during position and torque tasks.The lack of differences in activation reduction,despite some variability in tasks,is likely because rTE is velocity independent.36Another factor to consider in this study is the use of only surface EMG to observe activation strategies during position-and torque-matching tasks.Surface EMG is relevant for the study of rTE;however,specificneuromuscular-activation differences in position and torque tasks are better observed at the motor unit level.37If differences exist between position-and torque-matching tasks during dynamic contraction,surface EMG may not be sensitive enough to detect them.38,21Motor unit recordings could better determine whether any differences exist in motor neuron output between position and torque tasks during rTE protocols.As well,to investigate the adaptability of the neuromuscular system,39future studies should investigate activation reduction and potential changes to muscle energetics40following lengthening contractions in older adults.
5.Conclusion
Activation reduction in the isometric steady state following active lengthening is a feature of both torque-matching and position-matching tasks.Until the present study,the history dependence offorce in humans has been investigated via electricity-stimulated or voluntary contractions,with the active lengthening component controlled by a dynamometer motor,which is represented by our torque-matching experiments.Positon-matching tasks are inherently more complex owing to their unconstrained nature.Given that activation reduction was still present during the position-matching task,the data from our study provide us with further insight into the relevance of rTE in tasks that better represent voluntary human movement.
Data accessibility
Individual values of all supporting data are available upon request.
Acknowledgment
This project was supported by the Natural Sciences and Engineering Research Council of Canada(NSERC),Grant number:03829.Infrastructure was provided by the University of Guelph start-up funding.
Author contributions
GAP and RM contributed equally.Both authors have read and approved the final version of the manuscript,and agree with the order of presentation of the authors.
Competing interests
The authors declare that they have no competing interests.
杂志排行

Journal of Sport and Health Science的其它文章
- Isokinetic trunk flexion-extension protocol to assess trunk muscle strength and endurance:Reliability,learning effect,and sex differences
- Effects of compression garments on surface EMG and physiological responses during and after distance running
- Postural control quantification in minimally and moderately impaired persons with multiple sclerosis:The reliability of a posturographic test and its relationships with functional ability
- Health-related fitness knowledge growth in middle school years:Individual-and school-level correlates
- Habitual physical activity levels and sedentary time of children in different childcare arrangements from a nationally representative sample of Canadian preschoolers
- Walking-friendly built environments and objectively measured physical function in older adults