竹材细胞壁水分研究进展
2020-12-16方长华张淑琴陈美玲罗俊吉费本华
袁 晶,方长华,张淑琴,陈美玲,陈 琦,刘 嵘,罗俊吉,费本华
(1.国际竹藤中心,北京100102;2.国家林业与草原局/北京市共建竹藤科学与技术重点实验室,北京100102;3.加拿大不列颠哥伦比亚大学,加拿大)
研究以竹材纤维饱和点为切入点,讨论了细胞尺度下竹材中水分变化对细胞壁的影响,并归纳了分子尺度的水分类型、水分位置等研究成果,总结了近年来宏观及微观尺度上测试水分的相关联用仪器、测试方法。通过对细胞壁水分的研究综合认知,探讨竹材中水分研究重要性,提出竹产业发展所面临的机遇和挑战。
图1 竹材中水分存在状态及相关成果Fig.1 Status ofwater in bamboo and summary of relevant literature on bamboo
1 竹材纤维饱和点
纤维饱和点是指植物细胞腔没有水分,而细胞壁水分处于饱水状态时材料的含水率,是生物材料物理力学性能变化的一个拐点[5]。早在1988年,周芳纯[6]利用力学法测试竹材纤维饱和点为30% ~35%。关明杰[7]利用同样的方法测得毛竹(Phyllostachys edulis)的纤维饱和点为36.82%~42.55% (图2a)。水分的存在除了影响材料的力学性能,对材料的干缩湿胀性、介电性、质量等性质也存在一定的影响。因此,干缩法、热重法等手段也被应用于竹材纤维饱和点的测试。不同测试方法测得的纤维饱和点不同(图2a),陈森等[8]利用干缩法、力学法及电学法测试材料的纤维饱和点,指出干缩法测定木材的纤维饱和点是最精确的。关明杰[7]利用干缩法测试的纤维饱和点,发现纤维饱和点范围较大,介于19.76% ~50.34%之间。核磁技术可通过表征微观水分的状态对竹材的纤维饱和点进行测试,发现竹材的纤维饱和点在31.11% ~39.24%[9](图2a)。利用动态水蒸气吸附仪(DVS)对14种成熟竹材的吸湿等温线拟合发现,竹材的纤维饱和点在16.37%~27.91%,略低于木材22%~35%[10-11]。目前,通过质量变化获得竹材吸湿等温线的方法测试的纤维饱和点明显低于核磁,热重等手段[7]。力学法、干缩法所得到纤维饱和点的范围相比核磁法测得的大[6-7,12],这种范围偏差可能是由于竹材细胞结构的差异性导致的。
竹材主要由纤维和薄壁细胞组成,为进一步了解竹材的吸湿性,江泽慧等[13]首次提出针对竹材组成单元的吸湿性进行测试,通过离析获得竹材纤维及薄壁细胞,利用饱和盐溶液测定薄壁细胞的纤维饱和点为44.24%,显著大于纤维的纤维饱和点34.07%。但离析破坏了材料天然的化学组分和结构,因此获得的竹材纤维及薄壁细胞的纤维饱和点均大于天然竹材。通过物理分离竹材的纤维和薄壁细胞,发现高纯度的纤维组织和薄壁细胞组织的含水率存在一定的差异[10]。当纤维和薄壁细胞以一定的梯度规律排列在竹材中时,由于相同温湿度条件下纤维和薄壁细胞之间的湿度差,水分会发生一定的扩散,使材料表面的湿度平衡。这种扩散可能会导致天然竹材中薄壁细胞中的实际水分低于纯薄壁细胞的纤维饱和点,而纤维中的实际水分高于纯纤维组织的纤维饱和点。因此,在一定的温湿度条件下,竹材中不同的细胞类型其水分存在位置极有可能存在一定差异,即不同细胞,纤维饱和点不一样。
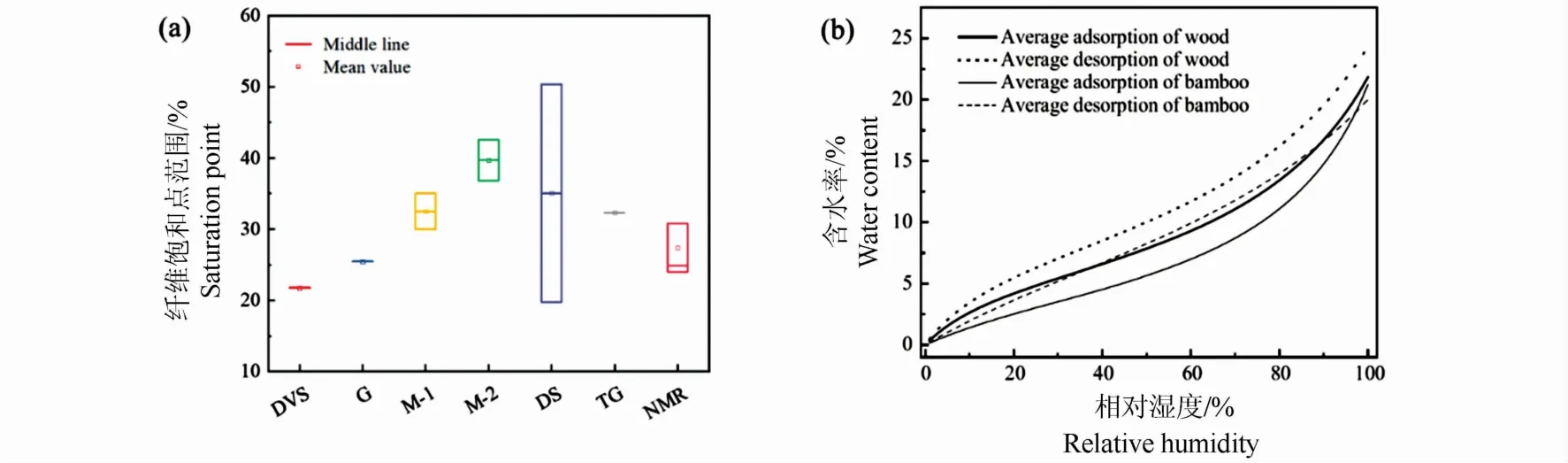
图2 竹材纤维饱和点范围及GAB模型模拟竹木材吸湿等温线Fig.2 The bamboo fiber saturation point range and GABmodel simulated the water absorption isotherm of bamboo wood
纤维饱和点不仅出现在竹材物理力学试验方法、竹材干燥标准等行业标准中,在实际加工生产中的作用也不可小觑[14]。例如,竹材的贮藏及运输过程中竹材的霉变程度随着含水率的增加而加剧[15]。目前竹材纤维饱和点的研究较少且测试结果存在较大的差异,制约了竹材的加工利用及各类加工标准的形成。因此,对于毛竹纤维饱和点的进一步确认是有必要的。
2 宏观尺度竹材水分特征
宏观尺度下,竹材中水分含量随环境湿度的变化而变化。吸湿解吸等温线是表现相同温度下材料质量随环境湿度的变化。吸湿等温线是宏观尺度下描述材料吸湿解吸过程主要的表征手段之一。图2(b)中竹材与木材这种“S”形的吸湿等温线为Ⅱ型曲线,该曲线类型发生在非多孔性固体表面或大孔固体上自由的单一多层可逆吸附过程[16-17]。吸湿等温线不仅可以在宏观上表现材料的含水率,也可以通过分区和模型分析微观水分子的存在方式,因此材料吸湿等温线的确定对于吸湿性能的研究十分重要。
2.1 吸湿过程
吸湿过程中,通过吸湿等温线的增加趋势,被分为3个区域。第1阶段,主要是不冻结水,对材料没有明显的增塑作用;第2阶段,水分吸附会引起材料骨架的溶胀;第3阶段,吸附水接近于自由水,有利于化学反应的进行和微生物的生长[18]。
利用相应的数学模型分析可以得到材料中水分类型,将宏观尺度中材料质量的变化与微观水分的类型建立联系。其中GAB和H-H模型在竹材中的应用最为广泛。在不同的环境下,材料提供模型会有不同的参数回应[11]。通过GAB对材料吸湿等温线进行非线性拟合发现竹材平均单分子层含水量为4.47%,低于木材的平均单分子层含水量[9]。H-H模型得到的单位水分子对应吸附位点的物质的量范围在111.37~231.38。由于数学表达和物理意义的差异,相比于H-H模型,GAB模型拟合得到的单分子吸附水的含量最大[19]。在宏观层面,不同位置,不同方向竹材的吸湿性不同。对竹材竹间及竹节的吸湿速度进行测试,发现竹材的竹间的吸湿速度大于竹节的吸湿速度。且竹材纵向吸湿膨胀率小于径向吸湿膨胀率和弦向吸湿膨胀率[10,20]。
濮阳市职业中等专业学校卢巧真在《中职学前教育专业多元化人才培养模式探索》中提出:“要根据学生发展的可能性及学生的自身需求来设立有短期就业需要、有专业发展需要、有学历教育需要等多元化培养方案来满足他们对就业的多元化需求。”多元化的学前教育人才主要包括应用型幼儿教师、儿童读物研 发及其他儿童文化工作者等复合型学前教育专业人才,要培养这样的人才,首先,政府要高调介入,制定政策及制度来保障高校教学管理的顺利进行。其次,高校要保障课程与教学运行机制的畅通。最后,形成以政府、高校、教师、学生、用人单位为主体的学前教育本科专业多元化人才培养机制。
2.2 解吸过程
材料的解吸过程相当于干燥过程,其本质为材料中含水量逐渐减少的过程。竹材解吸过程的研究可为竹材干燥工艺提供基础的理论数据。Zhang X等[10]通过GAB模型对解吸过程也进行了拟合,发现其单层水分含量为9.07%,显著大于吸湿阶段的4.47%。这是由于GAB模型中的重要参数均与材料吸附时的分子相互作用及温度有关,解吸过程中水分与材料的相互作用与吸湿过程恰好相反,因此导致其相关参数值存在差异[21]。更多研究学者利用了材料干缩特性对其尺寸进行解吸过程进行分析。徐兆军等[22]基于短程扫描成像技术对木材纤维饱和点以上的水分分布及迁移进行了研究,提出干燥过程中材料的自由水水分分布不均匀,腔大壁薄,细胞间隙较大的早材中自由水含量较多,且在干燥过程中木材横截面上不同区域的水分变化不同。这种现象由于木材结构及水分梯度引起的。在纤维饱和点之下,吕黄飞[23]利用CT对红竹的解吸性能进行测试,发现竹材顶部的干缩率大于底部。这种现象可能是由于竹材中纤维和薄壁细胞的分布不均匀造成,竹材顶部的薄壁细胞含量明显低于底部,薄壁细胞的干缩率较小。材料的细胞类型及梯度结构,对解吸过程材料的尺寸变化有着重要的影响。
2.3 吸湿滞后
吸湿滞后性是指在同一温湿度下,吸湿过程中材料所达到的含水率低于解吸过程中材料的含水率。生物质材料的吸湿滞后曲线均呈现先增加,后下降的趋势。滞后现象可以通过吸附位点的不完全再水化过程及压缩应力的变化来解释[24]。滞后现象的产生被认为是侧向羟基键作用导致吸附过程中吸附位点减少造成的[25]。也有研究者指出,滞后现象与材料细胞壁的刚度有关,并证实木质素含量越多,滞后现象越严重[26-27]。此外,解吸和吸湿过程中水分所需能量的变化也是产生滞后性的主要原因。解吸和吸湿阶段能量的主要来源为材料的毛细管力及水分子的浓度差。解吸过程水分与材料直接形成的氢键断裂时需要吸收一定的能量使水分子脱离束缚,水分向外扩散仍需要能量。而吸湿过程中,水分子与材料聚合物之间,水与水之间的氢键结合会释放出热量,会为水分的运动提供一定的热量,使得其扩散范围增加。因此,滞后性的产生可能与水分吸附解吸过程中的能量变化也有一定的关系。然而,还有研究表明,当温度高于75℃时,木材不再存在滞后现象,这可能是由于材料软化点的变化引起的[28]。
通过对比11种竹材的吸湿滞后性,发现绿竹显示出较高的吸湿滞后峰值,而龙竹的吸湿滞后峰值较低。且在细胞层面,竹材薄壁细胞的吸湿滞后值在高湿度下明显低于纤维的吸湿滞后值,这表明不同结构及化学组分对材料的滞后性也有一定的影响[9]。
3 细胞尺度水分特征
水分被分为自由水、吸着水和化合水3类。其中,吸着水是重要的研究部分。吸着水是细胞壁中微毛细管的水。这部分水分与细胞壁中的纤维素,半纤维素及木质素等物质形成氢键结合,影响材料的物理力学,防霉防腐性能及耐久性等。
在细胞壁水平,水分对细胞壁存在一定的塑化作用。Yu Y等[29]通过测试不同湿度下细胞壁S2层的纵向弹性模量,发现随着含水率的增加,木材细胞壁S2层的纵向弹性模量降低。随着含水率的增加,竹材次生壁S2层的压入深度及残余变形分别增加了28.33%和5.69%,通过压入深度的变化可得材料的硬度也随含水率的增加而减少[12]。这些力学性能变化的主要原因是水分子与生物质材料分子之间的相互作用,因此,水分对材料性能的研究机理需要对分子层面水分与材料的相互作用进行了解。吸附水分子后,细胞壁中的孔体积和孔面积都会增加,较小的孔会合并为较大的孔,从而形成连通性良好的孔隙网络以便于水分的传输[30]。
成像技术也被应用于细胞壁尺度下水分对材料变形润胀情况的研究。原位成像技术的应用能够直接表征材料对水分变化的反应机制,是十分重要的研究手段。Chen Q等[31]人利用激光共聚焦测试了不同湿度下竹材细胞壁的湿胀情况,并指出细胞壁的润胀应变取决于细胞类型,与细胞的位置及方向无关,其中纤维细胞的绝对润胀最高。这是由于润胀与细胞壁的密度有直接的关系,不同细胞其细胞壁密度的差异导致细胞的润胀程度不同。此外,细胞的润胀是由亚细胞尺度驱动的,细胞壁S2层中存在各向异性膨胀,且孔隙率对于细胞壁膨胀的各向异性有积极的影响[32]。Derome O等[33]通过分子模拟技术指出,S2微型柱的膨胀系数表明其体积膨胀率远高于早材或晚材的细胞组织。在细胞尺度上,S1和S3层发挥了抑制作用。因此,关于细胞层面,水分对细胞壁润胀情况的影响主要归因于3个方面,密度、细胞壁孔隙结构及纤丝排列方向。
4 分子尺度水分特征
竹木材的构成分子组分及结构均存在一定的差异,但其主要的电负性官能团一致。在分子尺度,材料中具有电负性的基团均可与水分子形成氢键,因此吸附水具有不同的能量和存在状态。其中,高分子聚合物-水、水-水之间的相互作用所形成的氢键也存在相应的能量差。因此在分子层面上,水分子的存在形式较为丰富。此外,不同的化学物质对于水分的吸附位置也存在差异。
4.1 水分类型
早在20世纪初,Hailwood and Horrobin将植物体水分主要分为2类,一类是“Polylayer water”,第二类是“Monolayer water”,这类水和材料表面的活性基团形成氢键结合[19]。近年来,研究学者分别利用核磁共振技术及红外技术对不同湿度下水分的作用官能团进行测试。马尔妮等[34]利用核磁表征了干燥过程中水分状态的变化,自由水在含水率下降至20%左右才蒸发殆尽,对于单分子层吸附水的变化点则出现在含水率为6%左右。
利用低场核磁对4年生毛竹进行测试,推测毛竹单层吸附水的弛豫时间低于16 ms,多层吸附水在16~35 ms之间[9]。此外,Guo X等[35]利用红外技术将单层吸附水分为3种且只受到较少的官能团作用力,其中水分子结合官能团有C-O、C-O-C、C=O、-OH,最后1种氢键结合方式是-H2O-H2O之间的强氢键结合。如图4所示,半纤维素与水分子,纤维素与水分子,水分子之间的氢键类型都不同,这也证明了水分子在微观层面的类型不同[36]。
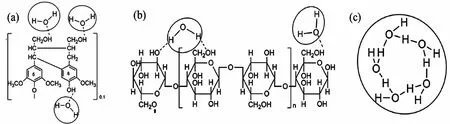
图3 部分氢键结合位点示意图[36]Fig.3 Schematic diagram of partial hydrogen bond binding sites[36]
此外,数学方法也被用来揭示水分的吸附机理。通过数学模拟,得到微观尺度下水分与材料的分子模型及相互作用场,其距离精度可以到达埃(Å),时间精度可以到达纳秒级别[37]。由于细胞壁上水分变化速度极快,故分子模拟对于水分吸附机理的揭示十分重要。Youssefian S等[38]通过分子模拟技术,利用氢键的键长对吸附水进行了分类,主要有3个峰,其中第一个峰为1.75Å处的氢键表明水分子与聚合物链上的羟基之间存在短距离的强氢键。第2个峰出现在2.45Å处,这是由于水分子与聚合物中侧向的氢原子之间的距离。在2.65Å处的第3个峰表示水分子上氧与羟基上的氧原子之间的距离。且随着含水率的增加,水分子更倾向与其他水分子形成氢键。竹材与木材的主要电负性官能团类似,因此在竹材主要组分中结合水类型应与木材基本相同。但竹材中存在大量的抽提物,这些抽提物可能对材料中的水分类型存在一定的影响[39]。在水分类型的研究中,竹材中的水分类型还有待于进一步的实验证实。
4.2 结合位置
水分吸附过程中的潜在吸附位点主要是羟基和羧基,在低含水率下,水分子往往会破坏聚合物链之间的氢键,在聚合物链之间形成新的氢键。在高含水率下,水分子倾向于与其他水分子形成氢键,并在材料内部形成纳米液滴水[38]。
水分的结合位置与化学物质的结构及化学键均有一定的关系。水分与材料官能团的结合是不断变化的。Kulasinski K等[40]发现水分子在半纤维素中被吸附,并且在纤维素与半纤维素的界面处出现过度吸附现象。尽管半纤维素含有更多的羟基,但木质素的羟基比半纤维素的羟基更易与水分子接触并形成氢键。半纤维素相比于木质素具有更加密集的氢键网络,因此水分子会渗透到半纤维素分子结构中,并使其自由体积增加,此时半纤维素聚合物内的氢键部分会遭到破坏[38]。聚合物位点之间氢键的破坏使得聚合物基团间距增加,基团之间的作用力变成长距离相互作用,从而降低了材料的机械模量并促进了润胀过程。
Guo X等[36]利用对木材中水分的存在类型进行测试,发现强、中、弱3种氢键结合水的对应红外波峰为3 149、3 496和3 602 cm-1,并指出当湿度大于55%时,强结合水的增长趋势明显且结合水为四面体结构。随着湿度的增加羰基C=O和C-O基团优先与水分子结合形成氢键[41]。Olsson A等[42]测得当环境相对湿度大于80%时,羧酸基团水分吸附速度高于羟基水分吸附速度。因此,水分的吸附基团不同及吸附阶段的不同对氢键的形成速度和键能的影响较大。竹材与木材均是由纤维素、半纤维素、木质素组成的高分子物质。然而,竹材中半纤维素主要的糖单元与木材存在一定的差异,因此在氢键形成时,其结合的部位及键长与木材不可能完全相同[43-44]。因此对于竹材微观层面水分的相关测试需要进一步的完善。
5 细胞壁水分测定方法
5.1 类饱和盐溶液法及成像技术联用
宏观尺度下,通过不同湿度下材料的质量变化测试材料中的水分。饱和盐溶液法是通过调节环境湿度从而影响材料的含水率,然而饱和盐溶液测试时需要将样品从密封的干燥器中移出再进行质量测试,这个移动的过程对样品本身的含水率结果存在一定的影响[45]。目前,动态水蒸气吸附仪(DVS)因其高重复率、高精度及操作简单而备受关注。DVS装置采取氮气及水分的混合气体对样品仓内的湿度进行调节,通过精密天平对样品质量实时记录[46],如图4a。然而,DVS的样品原始质量较小,对于变异性较大的生物质材料其测试结果具有一定的特殊性。
针对以上宏观细胞壁水分的测定方法,提出类饱和盐溶液法与成像技术的联用,既可以满足样品尺寸的不同选择及质量的实时检测,也可以对湿度变化下材料的干缩/润胀性进行观察(图4b)。通过对饱和盐溶液法及动态水吸附方法的认识,可以利用大的天平并建立相应的可调节湿度变化环境,进而测试大样品的吸湿性能,其中类饱和盐溶液法是指利用空气和水分的混合物调节空气湿度。如图4b所示,通过加湿器及温湿度控制器对玻璃器皿中的环境湿度进行调控。此外,在该装置中加入CCD或者利用实体显微镜对其端面的变化进行捕捉。其中CCD摄像倍数范围相比于实体显微镜较小,在精度要求较低的情况下CCD摄像头更为简便[48]。同时,也有研究学者将CCD摄像头替换为红外摄像头,并对材料吸湿解吸过程中材料端面的热学变化进行了测试,观察到木材表面的温度变化呈现各向异性[49]。
5.2 红外光谱技术与湿度调节装置联用
微观尺度材料中水分子的检测主要依赖核磁共振,红外光谱等技术[35,50]。核磁共振技术主要是利用电磁波使粒子从低能级向高能级跃迁时能级变化的对应时间测试水分子类型,但核磁共振测试成本较高。
红外光谱技术花费小,易操作分析,因此被广泛的应用于材料中水分的测试。红外光谱技术是利用共振原理对化学物质中分子的伸缩振动及弯曲振动进行测试。由于H-OH键的强烈伸缩震动,水分与细胞壁产生氢键结合时,氢键的存在对材料出峰位置及峰型展宽均会造成一定的影响,氢键相互作用的增强会导致红外波数的降低[51]。
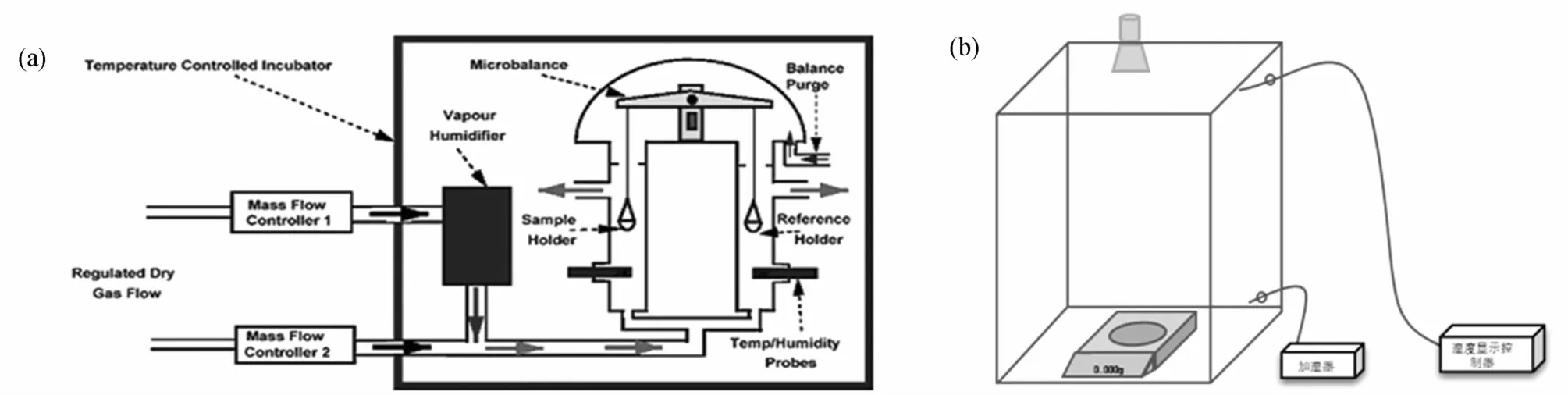
图4 宏观尺度材料水分的测试装置Fig.4 Test device for water ofmaterials atmacro scale
红外光谱技术与湿度调节装置联用,可以保证材料测试过程中含水率的稳定,从而提高实验数据的可靠度。图5为红外光谱技术与湿度调节联用装置示意图。通过给傅里叶显微红外提供一定的湿度环境,测试不同含水率下材料中的水分。参考DVS中的湿度调节,利用氮气和水分的混合气体对ZnSe玻璃容器内的相对湿度进行调节,并利用湿度计对玻璃容器内的湿度进行测定。傅里叶显微红外可直接测定不同细胞细胞壁区域组分红外光谱的差异,并进行材料微区分析。红外光谱技术与湿度调节装置联用技术可以在测试材料中水分类型的同时,也可以了解不同含水率下,材料官能团的变化,进而了解水分对材料化学组分的影响。

图5 红外与调湿装置的联用示意图[41]Fig.5 Schematic diagram of the combination infrared and humidity control devices[41]
6 结论与展望
细胞壁水分的研究在多个尺度上具有不同的特点。竹材是一种力学性能优异的梯度材料,因此竹材中水分与力学性能的关系被重点关注。竹材纤维饱和点在竹材的加工及利用过程中扮演着重要的角色,准确认识竹材纤维饱和点有利于竹产业的发展。然而目前竹材纤维饱和点的范围上没有明确定论,需进一步进行验证与完善。为了将更多的竹材付诸实际应用,必须要克服一些挑战。例如,为了实现竹材的稳定利用,必须了解环境水分对材料材性的影响,了解竹材的变形开裂问题。此外,为实现竹材的产业化应用,有必要对竹材加工时,竹材中水分的作用进行研究。在竹材人造板的加工中,水分也是非常重要的因素,其影响了人造板的质量[14]。
在宏观尺度,对材料水分的研究主要利用物理力学法进行测试,其中吸湿解吸等温线是非常重要的吸湿解吸过程的表征手段,不但能表示材料宏观层面的水分含量,也可以通过模型模拟将宏观水分含量与微观水分类型进行联系。同时,表征技术和模拟方法提高了我们对水分和材料相互作用的理解,使我们能够更精确地认识水分对细胞壁的影响,水分子的吸附类型,吸附位置等信息。
由于生物质材料的各向异性和测试技术的局限性,微观层面水分与细胞壁大分子的相互作用一直是研究的热点。竹材水分吸附机理的研究,大多数研究停留在吸湿等温线的测定上,分子层面的竹材吸湿性的相关研究较少。迄今为止,在分子水平,对细胞壁中水分的研究主要是利用核磁和红外等设备对调湿后的材料进行测试。然而,考虑到移动及测试过程中环境因素的变化,应尽量使用联用装置,减少样品测试期间水分的变化,保证测试的准确性。
