外源氮添加对2种豆科牧草功能性状的影响
2020-12-09张中信岳奇奇钱申吴静陶冶
张中信 岳奇奇 钱申 吴静 陶冶

摘要:以2种常见豆科牧草紫花苜蓿(Medicago sativa)和紫云英(Astragalus sinicus)为材料,设置对照、低氮和高氮(0 kg/hm2、50 kg/hm2、100 kg/hm2)3个梯度的外源氮添加处理,对比研究了2种豆科植物苗期形态、生物量分配、含水量及叶片光合色素含量等功能性状对外源氮添加的响应及性状间的权衡特征。结果表明:随施氮量增加,紫花苜蓿形态性状及含水量根冠比,紫云英光合色素特征及叶宽、株高、叶厚均有显著变化(P<0.05),但变化趋势大多不相同。2种豆科牧草形态性状总体表现为低促高抑效应,而叶绿素b和类胡萝卜素含量则表现为施氮后明显降低。相比之下,氮添加对2种豆科牧草生物量根冠比、含水量、叶绿素a及总叶绿素含量均无显著影响(P>0.05)。主成分分析结果表明,紫花苜蓿和紫云英前2个主成分所反映的功能性状协同性在种间差异较大。可见,2种豆科牧草生长和光合色素特征对外源氮添加具有明显的差异化响应,并表现出不同的协同性。
关键词:氮;紫花苜蓿;紫云英;功能性状;协同性
中图分类号:Q948文献标识码:A文章编号:1000-4440(2020)05-1197-08
Abstract:In this study, two common leguminous forage plants, Medicago sativa and Astragalus sinicus, were used as materials, and three nitrogen addition treatments, i.e., control (0 kg/hm2), low nitrogen level (50 kg/hm2) and high nitrogen level (100 kg/hm2), were set up. Moreover, the responses of functional traits including morphology, biomass allocation, water content and photosynthetic pigments of two leguminous forage seedlings to exogenous nitrogen addition, and the trade-off characteristics among functional traits were comparably analyzed. The results indicated that with the increase of nitrogen application rate, the morphological traits and root-shoot ratio of water content of M. sativa, the photosynthetic pigment characteristics, leaf width, plant height and leaf thickness of A. sinicus were significantly changed (P<0.05), but the change trends were different in most cases. The morphological traits of two leguminous forage plants presented a low-promotion and high-inhibition effect, while the chlorophyll b and carotenoid contents decreased obviously after nitrogen input. In contrast, nitrogen addition showed no significant effect on root-shoot ratio of biomass, water content, chlorophyll a and total chlorophyll contents of two leguminous forage plants (P>0.05). The results of principal component analysis showed that the synergism of functional traits reflected by the first two principal components differed greatly among the two species. Consequently, the growth and photosynthetic pigment characteristics of the two leguminous forage plants exhibited differential responses to exogenous nitrogen addition, and also showed different covariant patterns.
Key words:nitrogen;Medicago sativa;Astragalus sinicus;functional traits;synergism
氮素是植物生長的必需元素,也是植物生长的主要限制性元素之一[1-4]。随着工农业发展、人口增长及全球环境变化,东亚地区氮沉降量逐渐增高,已成为继欧、美之后的第3大氮沉降集中区[2]。大气氮沉降可以给土壤带来一定量的外源性氮输入,对植物生长有一定的促进作用,且适量的氮添加也能部分缓解干旱对植物生长的影响[4-6],对农田作物生长有一定益处。但当氮沉降达到或超过一定量时,氮在土壤及植物体内积累过多,植物生长会受到限制[2]。不仅如此,高浓度的氮沉降会给陆地及水生生态系统的生产力、稳定性以及生物多样性带来显著的负面影响[1,3-4,7-9]。因此,研究氮沉降等外源性氮输入对不同功能群植物的影响有助于深入揭示植物适应对策,并对厘清农田、森林等不同陆地生态系统对氮沉降的综合响应具有重要意义。
非固氮植物是研究氮沉降效应的主体材料。大部分非固氮植物的生长本身受到氮素的限制,因此对氮添加的响应极为敏感[10-11]。而具有与根瘤菌共生固氮能力的植物,如豆科植物,其共生体所固定的氮,足以满足植物自身生长需求[12]。尽管如此,在氮沉降量日益增大的背景下,这些豆科植物依然要面对外源氮输入带来的影响,这是不可忽视的现实问题。有研究者发现,一定量(0~40 kg/hm2)的氮添加能够促进3年生紫花苜蓿(Medicago sativa)的生长并提高光合性能[13],但是不同量的外源性氮输入对豆科牧草苗期生长和生理的影响尚不明晰。
植物在与环境相互作用过程中,逐渐形成了很多外部形态和内在生理方面的适应对策[14-15]。功能性状是指对生态功能的形成产生影响,并对个体适合度和自身表现产生影响的性状,例如植株高度、叶片大小、生物量分配、光合能力等[14-17]。功能性状之间具有表型整合或表型协变关系,能客观地反映植物对自身各功能之间权衡及对环境变化的适应性[18]。氮添加能改变植物生长环境,而植物也能通过功能性状及其组合的变化调整环境变化(包括氮沉降)影响下的响应策略[2]。例如,氮添加使植物茎和叶生物量分配增加,进而显著降低根冠比,且在营养生长与繁殖上也会出现生物量权衡分配[10]。因此,采用多功能性状整合的方法有利于揭示豆科植物对氮增加的响应和适应策略。
本研究以2种豆科常见牧草紫花苜蓿和紫云英(Astragalus sinicus)为研究对象,设置3种氮施加梯度(最高为100 kg/hm2),采用大棚盆栽培养的方法,测定植物苗期形态指标、生物量、含水量及光合色素含量,采用单因素方差分析、相关性分析和主成分分析法,对比研究2种豆科植物多种功能性状对氮施加的响应以及功能性状间的相关性和协调性,以综合了解氮沉降等外源性氮输入对豆科植物生长影响,为氮沉降量逐渐增加情况下的植物生长权衡策略解析及生态系统管理提供科学依据。
1材料与方法
1.1试验材料与试验设计
紫花苜蓿和紫云英种子采自安庆市宜秀区牧草种植地,盆栽试验在安庆师范大学多功能温室中开展。选择农田施用量最大的尿素为氮源[13],制成水溶劑后使用。设置3个用量梯度,分别为高用量(100 kg/hm2)、低用量(50 kg/hm2)和不施加氮素的对照(CK,0 kg/hm2)。每个处理5个重复,共30盆。种子破除休眠后,选取颗粒饱满的种子,均匀播入直径20 cm的花盆,覆土浇水。待幼苗生长10 d后定苗(每盆12株),并开始氮施加处理,每2 d用喷壶给每盆植物施加100 ml氮素溶液,对照施加等量水。试验于2018年5月11日开始,35 d后取样测定相关指标。
1.2植株生长特征的测定
每盆随机选取6株植物,先测定植株高度,再将地上部和地下根系分开称鲜质量(电子天平精度为0.000 1 g)。随后在每株上选取3枚健康叶片,用直尺、电子游标卡尺等工具测量叶长、叶宽、叶厚,计算叶长宽比。将每盆6株植物地上部分和地下部分分别混合,放入75 ℃烘箱中烘24 h,再称其干质量。用公式计算地上部含水量和地下部含水量,含水量=[(鲜质量-干质量)/干质量]×100%。再以此计算含水量的根冠比[(根含水量/地上含水量)×100%]和生物量的根冠比[(根干质量/地上部干质量)×100%]。
1.3光合色素含量的测定
每盆选取6株植株,每株选择3枚健康叶片,共18枚叶片作为1个重复。叶片剪成条状,用电子天平(精度为0.000 1 g)称取0.2 g样品,采用95%乙醇法萃取光合色素。用刻度试管定容至20 ml,在黑暗条件下静置24 h,期间摇晃2次。利用分光光度法测定上清液在波长为645 nm、663 nm及470 nm处的吸光值,根据Lichtenthaler方法计算叶绿素a、叶绿素b、类胡萝卜素含量及总叶绿素含量[19],并计算叶绿素a/b值。
1.4数据处理
采用单因素方差分析对比叶长、叶宽、叶厚、叶长宽比、叶绿素a含量、叶绿素b含量、类胡萝卜素含量、叶绿素a含量与叶绿素b之比、地上部含水量、地下部含水量、含水量根冠比及生物量根冠比在不同氮添加量处理间的差异性,利用Levenes test进行方差分析,使用Duncans法进行多重比较。用Pearson相关性分析检验氮施加影响下2种豆科植物功能性状之间的相关性。用主成分分析(PCA)解析不同功能性状的协同变化关系[18,20]。单因素方差分析、Pearson相关性分析以及主成分分析采用SPSS 20.0(SPSS Inc. USA)软件完成,常规数据分析采用Excel 2016完成。
2结果与分析
2.12种豆科牧草形态性状对不同用量氮的响应
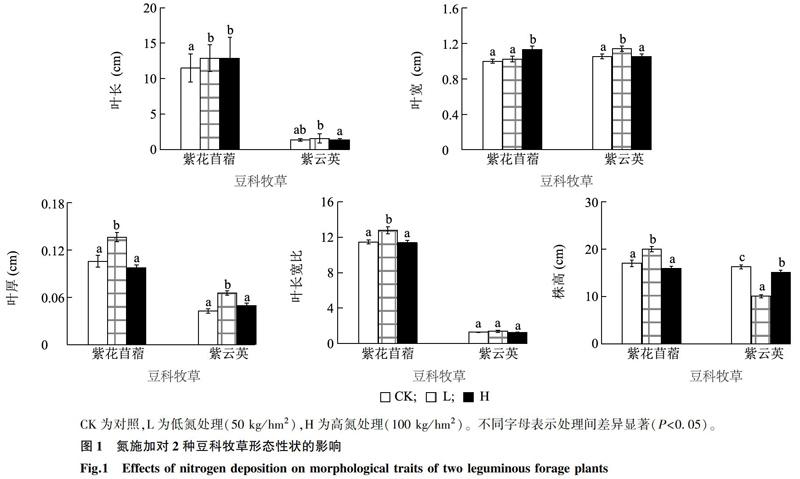
不同氮用量处理下,2种豆科植物形态特征具有不同程度的响应趋势(图1)。随氮施加量的增加,紫花苜蓿的叶长和叶宽也显著增大(P<0.05),即H(高氮组)>L(低氮组)>CK(对照组);而株高、叶厚及叶长宽比则呈先增后降趋势,表明出现了高用量抑制的现象。对紫云英而言,其叶长、叶宽及叶厚也呈现先增后降的趋势,但其叶长宽比在不同施氮用量下无显著差异。此外,紫云英株高在氮施加条件下显著降低,其中低用量下株高降低最明显,表明氮施加后紫云英以横向生长为主。
2.22种豆科植物含水量与生物量分配对不同用量氮的响应
随施氮量的增加,紫花苜蓿和紫云英的地上部含水量、地下部含水量及生物量根冠比均无显著差异(图2),但紫花苜蓿含水量根冠比在高氮下显著低于对照组(P<0.05),表明施氮没有改变2种豆科植物生物量分配,但抑制了紫花苜蓿根的水分吸收和利用。
2.32种豆科植物光合色素特征对不同用量氮的响应
随着施氮量增加,紫花苜蓿叶绿素a、类胡萝卜素含量和总叶绿素含量变化均不显著(图3);其叶绿素b含量表现为CK>H>L,即施氮显著降低了叶绿素b含量;对照组紫花苜蓿叶绿素a/b值显著低于低氮处理组(P<0.05)。对紫云英而言,其叶绿素a和总叶绿素含量在处理间均无显著差异,但其叶绿素b、类胡萝卜素、叶绿素a/b值均表现为施氮后显著降低。
2.4不同施氮量下2种豆科牧草功能性状的相关性
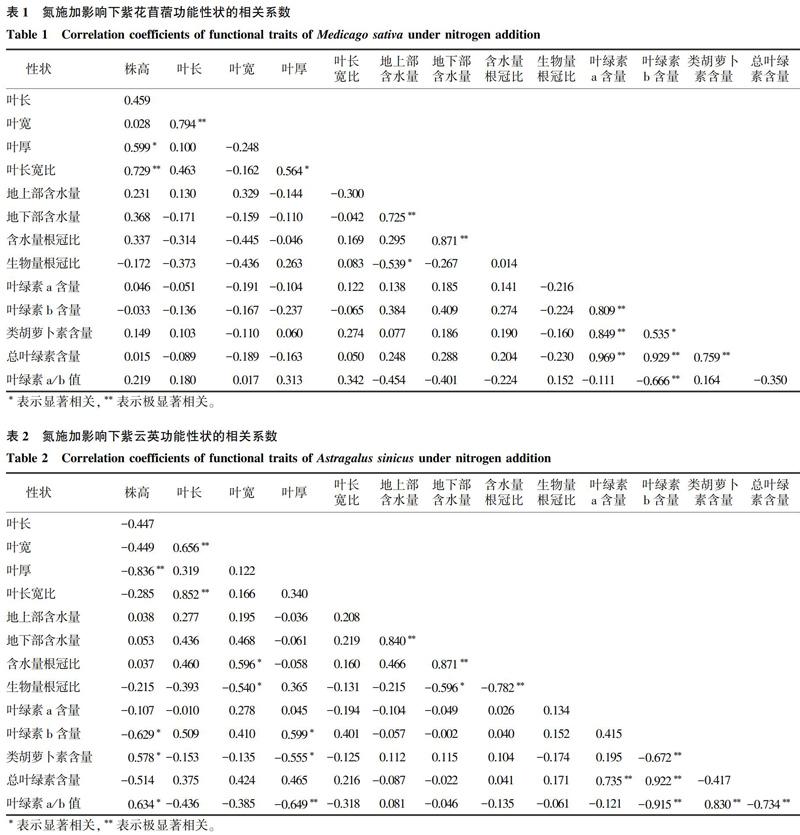
相关性分析结果表明,紫花苜蓿叶绿素a/b值与叶绿素b含量呈极显著负相关关系,总叶绿素含量与叶绿素a、叶绿素b、类胡萝卜素含量呈极显著正相关关系,类胡萝卜素含量与叶绿素a和叶绿素b含量分别呈极显著和显著正相关关系(表1)。生物量根冠比与地上部含水量呈显著负相关关系,含水量根冠比与地下部含水量之间及地下部含水量与地上部含水量之间均极显著正相关关系。叶长宽比与株高和叶厚分别呈极显著和显著正相关关系,叶宽与叶长也呈极显著正相关关系。
紫云英叶绿素a/b值与株高、类胡萝卜素含量分别呈显著和极显著正相关关系,与叶厚、叶绿素b含量以及总叶绿素含量呈极显著负相关关系(表2)。叶绿素b含量与叶厚呈显著正相关关系,总叶绿素含量与叶绿素a、叶绿素b含量呈极显著正相关关系,类胡萝卜素含量与叶绿素b含量呈极显著负相关关系。生物量根冠比与叶宽、地下部含水量及含水量根冠比呈显著或极显著负相关关系。含水量根冠比与叶宽和地下含水量分别呈显著和极显著正相关关系。叶厚与株高呈极显著负相关关系,叶长与叶宽呈极显著正相关关系。
2.5不同施氮量下2种豆科牧草功能性状的权衡特征
由表3可知,在紫花苜蓿PCA第1主成分中,地上部含水量、地下部含水量、叶绿素a含量、叶绿素b含量、类胡萝卜素含量和总叶绿素含量的变化具有协同性(特征值的绝对值>0.5)。在第2主成分中,株高、叶厚、叶长宽比和叶绿素a/b值有一定的协同性,叶长、叶宽、生物量根冠比和含水量根冠比与其他性状并没有明显的协同变化趋势。
对紫云英功能性状的PCA分析结果表明,第1主成分中株高、叶长、叶宽、叶厚和叶长宽比、叶绿素b含量、类胡萝卜素含量、总叶绿素含量和叶绿素a/b值表现出较强的协变趋势,但株高、类胡萝卜素含量及叶绿素a/b值与其他性状的协变方向相反。在第2主成分中,叶宽、地上部含水量、地下部含水量、生物量根冠比、含水量根冠比具有协同变化特征,但生物量根冠比与其他性状的协变方向相反。此外,紫云英叶绿素a含量与其他性状均无明显的协变性。
由此可见,2种豆科牧草形态、生物量分配、含水量及叶片光合色素等功能性状对氮施加的协变趋势有较大差异,体现了明显不同的权衡特征。
3讨论
增加土壤养分会改变植物多方面的适应对策,其中形态学适应是一种最基本的机制[21]。氮添加使土壤可利用性氮增加,在一定程度上会促进植物生长。本研究结果显示,低氮处理组紫花苜蓿的株高显著高于其他2组,这与李双娥等[22]的研究结果一致,即适量的氮促进了海南红豆(Ormosia pinnata)与马占相思(Acacia mangium)地上部分的累积,过量氮反而抑制植物地上部分的生长[4,10]。然而紫云英的株高為对照组>高氮处理组>低氮处理组,可能是由于紫云英植株高度对氮的响应更加敏感,且出现了横向(匍匐)生长。紫花苜蓿叶长与叶宽表现为高氮处理组>低氮处理组>对照组,表明氮对叶片的生长有明显的促进作用[23]。而紫云英叶长与叶宽随着氮用量的增加先增后降,这与大豆叶片面积响应氮添加的结果相一致[24]。紫花苜蓿的叶厚与叶长宽比和紫云英的叶厚随着氮用量的增加先增后降,是典型的低促高抑现象。
植株体内不同器官的有机物分配往往会根据外界环境的变化而改变,以提高自身对环境的适应能力[10]。根系是豆科植物生长的重要结构和物质基础。本研究中紫云英的根冠比顺序为对照组>低氮处理组>高氮处理组,即施氮降低了植物根冠比,这与木荷(Schima superba)等木本植物响应氮添加的结果一致[25];同时,该现象也与荒漠草本植物对氮的响应相符,即植物在氮施加后,地上部分的生长比地下根系快,进而导致根冠比降低[26]。这可能是因为对照组植物受到了氮的限制,进而分配给地下根系更多生物量以吸收营养,保证植物正常营养供应,且这种限制会在氮施加后逐渐解除[10]。紫花苜蓿的根冠比顺序为低氮处理组>对照组>高氮处理组,即根冠比先增后降,其中高量氮添加下根冠比下降趋势更明显,这也符合氮添加后植物根冠比下降的普遍规律[4,10]。但一些研究结果表明,在较低的施氮水平下,施氮增加了紫花苜蓿的地下生物量[27],说明植物对体内生物量分配比例的调控也受到氮施用量的影响,低用量促进而高用量抑制。两种植物含水量的变化并不显著,表明氮用量对植物含水量的影响有限。
植物叶片是光合作用的主要器官,光合色素含量是衡量光合作用强弱的重要指标,环境因子的变化会影响植物光合色素含量[28-34]。本研究中对照组紫云英类胡萝卜素含量大于施氮组,表明氮对紫云英类胡萝卜素的合成有抑制作用。叶绿素a是光合作用中心色素分子,具有把光能转换为电能,进行电子传递并最终转化成化学能的功能,而叶绿素b具有吸收光能并传递光能的能力[32]。本研究中,紫云英和紫花苜蓿的叶绿素b含量大小顺序为对照组>高氮处理组>低氮处理组,而叶绿素a变化不显著,这可能是由于低氮促进叶绿素b的合成,而高氮抑制其合成。叶绿素a在遮光条件下会加速向叶绿素b转化,使叶绿素a/b值降低,有利于提高植物对短波蓝紫光(弱光)的利用率。而叶绿素a/b值升高则表明,植物在强光下光合速率会提高,能明显抵抗光抑制[32,35]。在本研究中,紫花苜蓿叶绿素a/b值低氮处理组显著大于对照组,而紫云英为对照组大于施氮处理组,表明施氮后紫花苜蓿对强光的利用率提高,而紫云英则对弱光的利用率较高。
紫花苜蓿和紫云英株高、叶宽、含水量根冠比、类胡萝卜素含量、叶绿素a/b值对不同氮用量的响应表现出较为明显的差异性,这可能是由种间差异性导致的,即不同物种对氮添加的响应不同。厘清这种多功能性状的差异性,就需要进行性状整合分析。研究结果表明,紫花苜蓿和紫云英植株功能性状之间具有一定相关性,也表现出一定的协变特征,尤其在叶片光合色素方面。表型协变是植物性状之间互相作用而协同变化的现象[36],它可以影响功能性状间的变化而适应环境的变化,从而形成功能性状间的最佳组合[18,37],是物种进化的重要标志。紫花苜蓿的含水量特征(地上含水量、地下含水量)与光合性状(叶绿素a、叶绿素b、类胡萝卜素、总叶绿素含量)之间表现出较强的整合格局,反映了紫花苜蓿的含水量和光合色素特征之间具有适应性的协同作用,是氮施加影响过程中植物体发育性状间相互权衡的结果。紫云英的形态性状(株高、叶长、叶宽、叶厚和叶长宽比)和光合生理性状(叶绿素b含量、类胡萝卜素含量、总叶绿素含量和叶绿素a/b值)之间也有较强的协变趋势,其中株高、叶绿素a/b值和类胡萝卜素含量与其他性狀具有反向协变性,说明紫花苜蓿地上部分功能性状具有明显的整体性;紫云英生物量根冠比与其他性状的协变趋势相反,这表明紫云英在氮施加下会通过形态结构和生理特征响应,优先调配资源给地上部分,形成地上部分和地下部分的不一致性,也即产生了地上部分和地下部分的权衡。
4结论
不同量氮施加对紫花苜蓿形态性状、含水量根冠比、叶绿素b含量、叶绿素a/b值和紫云英的形态性状及光合色素特征均具有显著影响。两物种间部分功能性状(如叶厚、叶绿素a含量、叶绿素b含量、地上地下含水量等)对氮添加的响应具有相同趋势,但大部分功能性状存在种间差异性。不仅如此,2种物种功能性状对氮素施加的协同适应性也存在种间差异。研究证实2种不同豆科牧草对外源氮添加具有差异化的响应特征和性状协变趋势,其机理需进一步研究。
参考文献:
[1]BOBBINK R, HICKS K, GALLOWAY J, et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis[J]. Ecological Applications, 2010, 20(1):30-59.
[2]毛晋花,邢亚娟,马宏宇,等. 氮沉降对植物生长的影响研究进展[J]. 中国农学通报, 2017, 33(29):42-48.
[3]刘学军,张福锁. 环境养分及其在生态系统养分资源管理中的作用: 以大气氮沉降为例[J]. 干旱区研究, 2009, 26(3):306-311.
[4]常运华,刘学军,李凯辉,等. 大气氮沉降研究进展[J]. 干旱区研究, 2012, 29(6):972-979.
[5]张维,赵文勤,谢双全,等. 模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响[J]. 中国生态农业学报, 2018, 26(1):106-115.
[6]肖迪,王晓洁,张凯,等. 模拟氮沉降对五角枫幼苗生长的影响[J]. 北京林业大学学报, 2015, 37(10):50-57.
[7]CLARK C M, TILMAN D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands[J]. Nature, 2008, 451(7179):712-715.
[8]张燕,崔学民,樊明寿. 大气氮沉降及其对草地生物多样性的影响[J]. 草业科学, 2007, 24(7):12-17.
[9]苏洁琼,李新荣,回嵘,等. 氮沉降对荒漠化草原草本植物物种多样性和群落组成的影响[J]. 西北植物学报, 2012, 32(4):795-801.
[10]毛晋花,邢亚娟,闫国永,等. 陆生植物生物量分配对模拟氮沉降响应的 Meta 分析[J]. 生态学报,2018,38(9):3183-3194.
[11]万宏伟,杨阳,白世勤,等. 羊草草原群落6种植物叶片功能特性对氮素添加的响应[J]. 植物生态学报, 2008, 32(3):611-621.
[12]赵叶舟,王浩铭,汪自强. 豆科植物和根瘤菌在生态环境中的地位和作用[J]. 农业环境与发展, 2013, 30(4):7-12.
[13]李雪芬. 氮添加对紫花苜蓿生长特性及光合生理特性的影响[J]. 北方园艺, 2015(2):158-164.
[14]刘晓娟,马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45:325-339.
[15]何念鹏,刘聪聪,张佳慧,等. 植物性状研究的机遇与挑战: 从器官到群落[J]. 生态学报, 2018, 38(19): 6787-6796.
[16]LAVOREL S, GRIGULISK, LAMARQUE P, et al. Using plant functional traits to understand the landscape distribution of multiple ecosystem services[J]. Journal of Ecology, 2011, 99:135-147.
[17]LIENIN P, KLEYER M. Plant trait responses to the environment and effects on ecosystem properties[J]. Basic and Applied Ecology, 2012, 13(4):301-311.
[18]邱东,张静,吴楠,等. 安徽大龙山国家森林公园东亚小金发藓配子体与孢子体个体性状及其相互关系[J]. 植物研究, 2019, 39(6):835-845.
[19]LICHTENTHALER H K, WELLBURN A R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents[J] . Biochemical Society Transactions, 1983, 11:591-592.
[20]CONCHITA A, CARLOS M H. Patterns made of patterns: Variation and Covariation of leaf nutrient concentration and between populations of Prunus mahaleb[J]. New Phytologist, 2001, 150:629-640.
[21]PATTERSON T B, GUY R D, DANG Q L. Whole-plant nitrogen- and water-relations traits, and their associated trade-offs, in adjacent muskeg and upland boreal spruce species[J]. Oecologia, 1997, 110(2):160-168.
[22]李双娥,李义勇,方熊,等. 不同氮添加和添加方式对南亚热带4个主要树种幼苗的生长的影响[J]. 植物生态学报, 2019, 39(10):950-961.
[23]武新红,彭克勤,梁宗锁,等. 氮磷对菘蓝营养生长期根、冠生长的影响[J]. 西北农业学报, 2008, 17(4):274-278.
[24]宋英博. 施氮对大豆叶片生理指标的影响[J]. 中国农学通报, 2010, 26(15):243-246.
[25]李明月,王健,王振兴,等. 模拟氮沉降条件下木荷幼苗光合特性、生物量与C、N、P分配格局[J]. 生态学报, 2013, 33(5):1569-1577.
[26]周晓兵,张元明,王莎莎,等. 3种荒漠植物幼苗生长和光合生理对氮增加的响应[J]. 中国沙漠, 2011, 31(1):82-89.
[27]ESECHIE H A, AL-BARHI B, AL-GHEITY S, et al. Root and shoot growth in salinity-stressed alfalfa in response to nitrogen source[J]. Journal of Plant Nutrition, 2007, 25(11):2559-2569.
[28]袁孟玲,岳堃,王红,等. 增强UV-B辐射对芒果成年树光合作用及其产量与常规品质的影响[J].南方农业学报,2018,49(5):930-937.
[29]高祺,李明,朴琳,等. 拔节期弱光和渍水胁迫对春玉米光合作用、根系生长及产量的影响[J].江苏农业学报,2018,34(6):1276-1286.
[30]郝正刚,王志恒,魏玉清,等.外源钙镉处理对甜高粱幼苗叶片光合作用的影响[J].江苏农业科学,2019,47(19):75-80.
[31]DE E, DONG D, ZENG X, et al. Direct effect of acid rain on leaf chlorophyll content of terrestrial plants in China[J]. Science of the Total Environment, 2017, 605/606:7-769.
[32]李合生. 现代植物生理学[M]. 3版. 北京: 高等教育出版社, 2012:100-140.
[33]李君,娄运生,马莉,等. 夜间增温和水分管理耦合对水稻叶片光合作用和荧光特性的影响[J].江苏农业学报, 2019, 35(3):506-513.
[34]王锐洁,刘筱,杨淑君,等. 氮沉降背景下遮陰对虎耳草生长和光合作用的影响[J].南方农业学报,2019, 50(2):330-337.
[35]占吉成,王利军,黄卫东. 弱光环境下葡萄叶片的生长及其在强光下的光合特性[J]. 中国农业大学学报, 2004, 7(3):75-78.
[36]KLINGENBER G, PETER C. Morphological integration and developmental modularity[J]. Annual Review of Evolution and Systematics, 2008, 39(1):115-132.
[37]施宇,温仲明,龚时慧. 黄土丘陵区植物叶片与细根功能性状关系及其变化[J]. 生态学报, 2011, 31(22):6805-6814.
(责任编辑:张震林)
