利用分子标记鉴定长粒粳稻品种粒形相关基因的基因型
2020-12-07周雷李二敬徐华山刘凯李培德游艾青
周雷 李二敬,2 徐华山 刘凯 李培德 游艾青,3*
(1 湖北省农业科学院粮食作物研究所/粮食作物种质创新与遗传改良湖北省重点实验室,武汉430064;2 武汉大学研究生院,武汉430072;3 长江大学/主要粮食作物产业化湖北省协同创新中心,湖北 荆州 434025;第一作者:13971364609@163.com;*通讯作者:aq_you@163.com)
水稻是我国重要的粮食作物。我国的水稻品种属于亚洲栽培稻,有粳稻和籼稻两个亚种,粒长是区分籼粳两个亚种的主要特征之一。籼稻谷粒一般较长,而粳稻谷粒相反。我国粳稻长宽比一般为1.5~2.0;长宽比大于2.5、整精米粒长≥6.5 mm 者是长粒粳稻[1]。近年来,随着人们生活条件的改善,优质米受到越来越多人的追捧,对稻米的需求已由原先单一的饱腹功能转变为集美味、营养于一体的复合型需求。受此影响,我国粳稻品种在选育过程中有长粒化的趋势[2],并出现了一批长粒粳稻知名品种和品牌,如东北的稻花香2 号[3]。
湖北省实施水稻“籼改粳”工程以来,粳稻发展取得了阶段性成效[3],选育了鄂粳403、鄂香2 号等一批优质粳稻新品种。其中,鄂香2 号长宽比达到3.2,是湖北省近年来培育并通过审定的第1 个长粒粳品种。为解析长粒粳的粒长性状的遗传基础,收集了全国不同区域的7 个长粒粳品种,利用分子标记检测方法,对10 个已克隆的水稻粒型基因进行了检测和分析。开展本研究不仅能对现有长粒粳品种粒型基因型有一个全面的认识,还可为今后利用分子标记辅助培育长粒粳新品种奠定重要基础。
1 材料与方法
1.1 实验材料及DNA 提取
供试水稻材料为7 份优质长粒粳材料,包括鄂香2 号、稻花香2 号、CLJ-AH(安徽省农科院)、CLJ-YZ(扬州大学)、CLJ-HZ(中国水稻研究所)、CLJ-HB(湖北省农科院)、CLJ-JX(浙江省嘉兴市农科院),对照为短粒粳稻品种日本晴。
所有水稻材料2018 年正季种植于湖北省农业科学院南湖水稻试验基地,取幼嫩叶片进行基因组DNA的提取。
1.2 水稻籽粒粒形性状相关数据测定
成熟期选取每个品种单株的主穗,考查植株粒型。将种子烘干,每个单株随机选择50 粒饱满的种子用考种仪器测量各品种谷粒的粒长、粒宽、长宽比及千粒重。获得的数据用Excel 2010 进行统计分析。
1.3 水稻粒形基因检测
参考前人报道,分别合成能够扩增GS3、LGY3/OsLG3b、qGL3、GL7/GW7、SLG7、TGW6、GS9 等 7 个粒长基因和 GS5、GW5、GW8 等 3 个粒宽基因共 10 个基因的引物序列(表1),引物均由武汉昆泰锐生物公司合成。
PCR 反应体系为 15 μL,含 1.0 μL 模板 DNA(50 ng/μL),0.2 μL Tag 酶 (5 u/μL),1.50 μL 10 ×Buffer(Mg2+plus),引物各 0.70 μL(66 ng/μL),0.8 μL dNTPs(2.5 mM/μL),补 ddH2O 至总体积为 15.00 μL。PCR 扩增程序为:95℃预变性 5 min;95℃变性 30 s,退火 30 s,延伸72℃,共35 个循环;最后72℃下延伸5 min,扩增产物在4℃下保存。
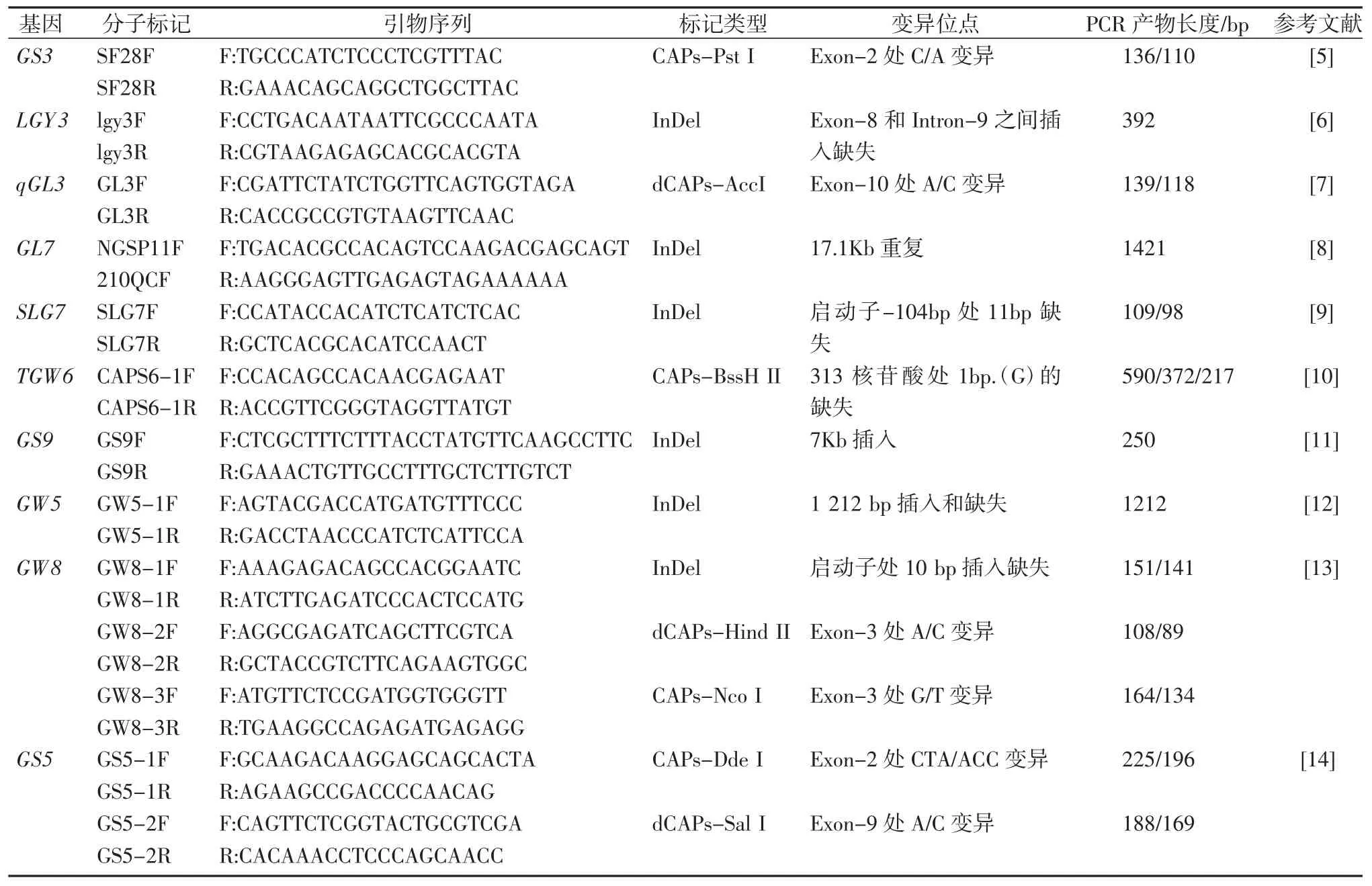
表1 水稻粒型相关功能分子标记
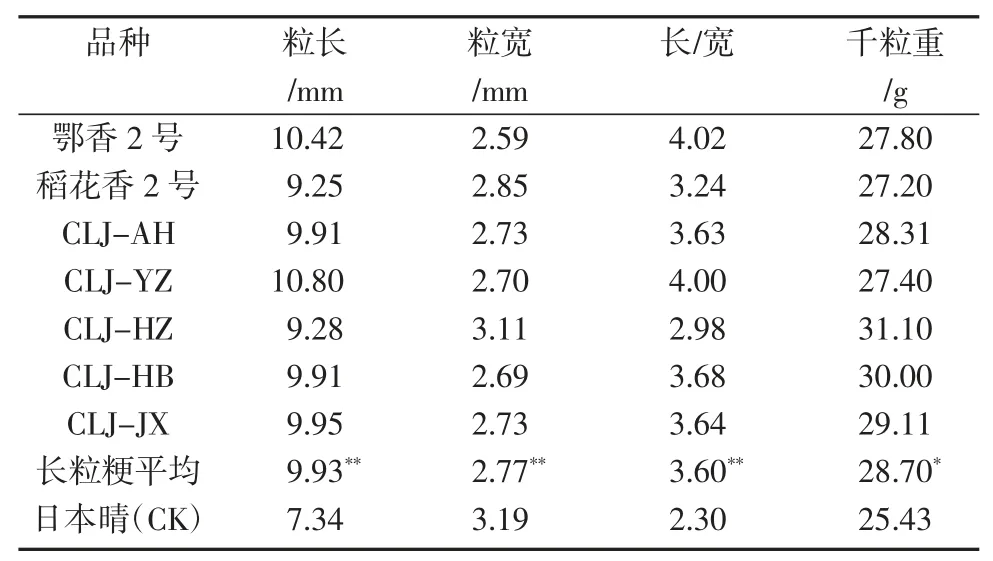
表2 7 个长粒粳品种谷粒粒形性状测量结果
酶切体系:取 PCR 扩增产物 5 μL 与 PCR 管中,加入 1 μL 相应的反应缓冲液,0.3 μL 限制性内切酶,用ddH2O 补足至 10 μL,37℃水浴 3 h,60℃停止反应。
电泳检测:分别用PAGE 胶或琼脂糖凝胶电泳进行目的条带检测。
2 结果与分析
2.1 7 个长粒粳品种粒形性状
从表2 可见,7 个长粒粳品种粒长在9.28~10.80 mm 之间,粒宽在 2.69~3.11 mm 之间,长宽比在 2.98~4.02 之间,千粒重在 27.20~31.1 g 之间。t 检验结果表明,这7 个品种的粒长、长宽比均大于日本晴,差异极显著(P<0.01);千粒重大于日本晴,差异显著(P<0.01);粒宽小于日本晴,差异极显著(P<0.01)。
2.2 基因型分析
2.2.1 GS3 基因型分析
GS3 是第1 个被克隆调控粒型的基因,是主要调控水稻粒长和粒重的主效基因,当其在第2 个外显子上的一个C-A 突变引起N 端调控器官大小的结构域发生功能缺失突变后,导致水稻籽粒变长。FAN 等[5]据此位点设计的引物SF28F/SF28R 对水稻材料基因组DNA 扩增,可以得到一条136 bp 的目的片段。短粒型等位基因品种的这个片段在PstI 酶切后,产生110 bp和26 bp 两条片段;长粒型等位基因由于C-A 突变,产生的136 bp 不能被PstI 识别和酶切,还是136 bp 的片段。检测结果显示,只有对照品种日本晴能被PstI 识别和酶切,扩增出110 bp 片段,含有GS3 基因;长粒粳品种都不能被PstI 识别和酶切,都含有长粒的gs3 等位基因(图 1)。
2.2.2 LGY3 基因型分析
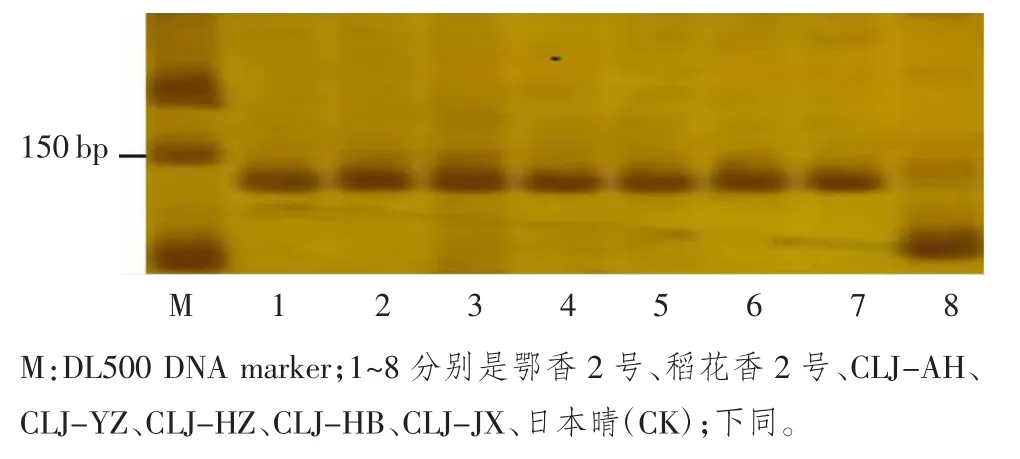
图1 7 个长粒粳品种GS3 基因型分析(PstI 酶切后)
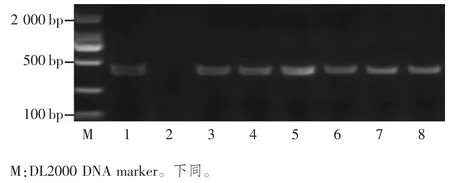
图2 7 个长粒粳品种LGY3 基因型分析

图3 7 个长粒粳品种qGL3 基因型分析(AccI 酶切后)
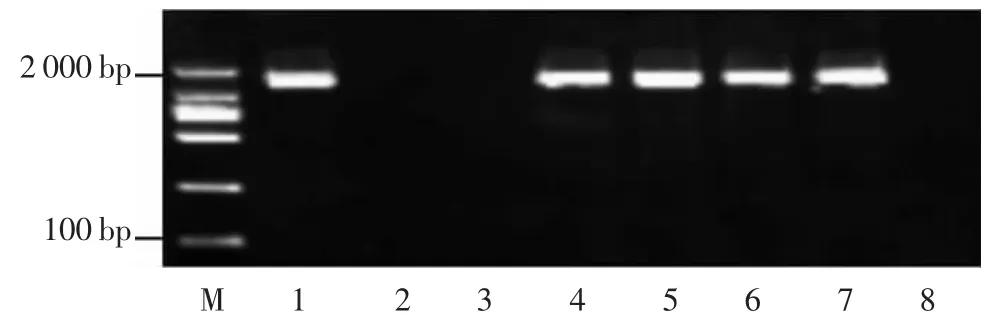
图4 7 个长粒粳品种GL7 基因型分析
LGY3/ OsLG3b 基因编码1 个含有MADS 域的转录因子OsMADS1,是G 蛋白βγ 二聚体下游的关键效应子[6,15]。LGY3 基因在第 8 个内含子和第 9 个外显子之间存在1 个缺失(TCCTTGGTGAAGGTA)插入(ATGTATATATACT)突变产生的功能缺失,使水稻的粒长增加。根据突变位点设计引物lgy3F/lgy3R 对所选水稻材料进行扩增,如果没有发生突变,则可以扩增出392 bp的序列,若突变成为长粒基因,则没有条带出现。实验结果表明,只有稻花香2 号含有lgy3 长粒等位基因,其它品种含有LGY3 短粒等位基因(图2)。
2.2.3 qGL3/GL3.1/qGL3-1 基因型分析
qGL3/GL3.1/qGL3-1 是位于水稻第3 染色体上控制粒长的基因,2012 年由我国3 个实验室同时报道,编码1 个含有2 个Kelch 功能域的蛋白磷酸酶OsPPKL1,在水稻粒长调控中发挥负调节子的作用[7]。在此基因的第10 个外显子处,有1 个 A/C 变异位点,与水稻的粒长相关。基因型A 的平均粒长超过基因型为C的平均粒长。裔传灯等人根据这个突变位点为qGL3开发了1 个分子标记,若基因型为A,产生的139 bp 的目的片段缺乏AccI 识别酶切位点,不能被AccI 酶识别和酶切;基因型为C 可被AccI 酶识别和酶切,产生1个118 bp 的大片段和21 bp 的小基因条带。PAGE 凝胶电泳检测结果显示,全部品种都能被AccI 识别和酶切,判断全都为C 基因型,属于不含qGL3/GL3.1/qGL3-1 长粒基因的品种(图3)。
2.2.4 GL7 基因型分析
GL7 是位于水稻第7 染色体上控制粒长的基因,编码一种与拟南芥长叶蛋白同源的蛋白,调节细胞的纵向伸长,GL7 位点的1 个17.1 kb 片段的串联复制导致GL7 的上调和附近负调节因子的下调,导致籽粒长度的增加和外观品质的改善。WANG 等[8]据此突变位点设计引物,若所检测水稻材料的基因存在17.1 kb 片段的串联重复片段,则进行PCR 时可以扩增出1 条1 421 bp 大小的片段,若没有,则进行PCR 之后没有扩增产物出现。检测结果显示,鄂香2 号、CLJ-YZ、CLJHZ、CLJ-HB、CLJ-JX 都能扩增出 1 421 bp 的片段,判断其含有GL7 长粒基因型,稻花香2 号、CLJ-AH、日本晴没有条带,判断其不含有GL7 长粒基因(图4)。
2.2.5 SLG7 基因型分析
SLG7 是水稻第7 染色体上控制粒长的基因,纵向增加细胞长度而横向减小细胞宽度,使籽粒细长[9]。通过比对研究发现,长粒等位基因启动子-104 bp 处11 bp 的缺失与粒型紧密连锁,据此位点开发InDel 分子标记M-SLG7-F/M-SLG7-R。利用该标记对水稻材料进行扩增,用PAGE 电泳时可以区分出11 bp 的插入-缺失,含有11 bp 的材料可以扩增出大小为109 bp 的DNA 片段,不含11 bp 的扩增出98 bp 的片段。检测结果表明,鄂香 2 号、CLJ-YZ、CLJ-HZ、CLJ-HB、CLJ-JX扩增出98 bp 的片段,表明其含有SLG7 长粒基因,稻花香2 号、CLJ-AH、日本晴扩增出109 bp 的片段,则表明它们不含SLG7 长粒等位基因(图5)。
2.2.6 TGW6 基因型分析

图5 7 个长粒粳品种SLG7 基因型分析
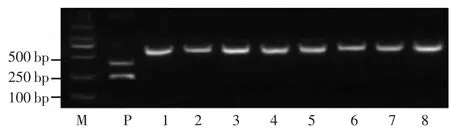
图6 TGW6 基因型分析-BssHⅡ酶切
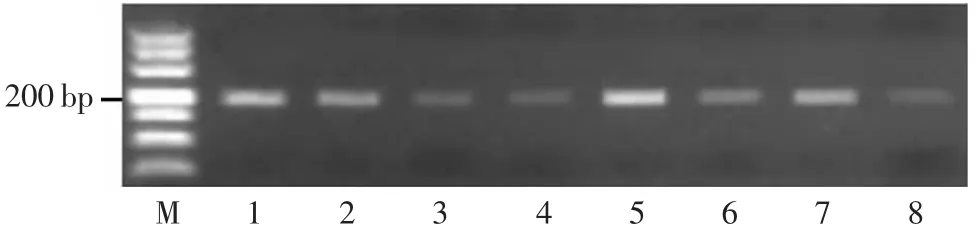
图7 7 个长粒粳品种GS9 基因型分析
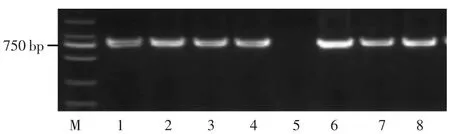
图8 7 个长粒粳品种GW5 基因型分析
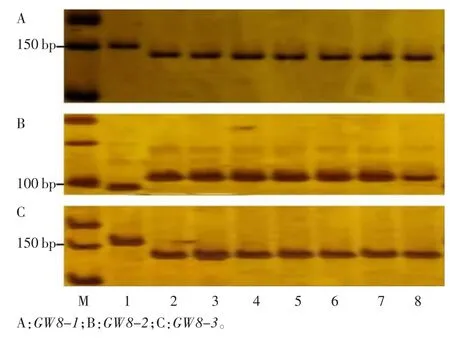
图9 7 个长粒粳品种GW8 基因型分析
TGW6 是位于6 号染色体上控制水稻粒长和千粒重的基因,编码IAA-葡萄糖水解酶,只有1 个外显子而没有内含子。王军等[10]设计了1 个CAPS-BssHII 标记,利用此标记对水稻基因组进行扩增,可以得到1 个590 bp 的目的片段,长粒等位基因型可被BssHII 识别和酶切,产生1 个372 bp 和217 bp 2 个条带;短粒等位基因PCR 产物不能被BssHII 限制内切酶识别和酶切,还是590 bp 条带。结果所示,阳性对照II-32B 有2个带条,7 个长粒粳品种和日本晴不能被BssHII 识别和酶切只有1 个条带,判断其不含TGW6 长粒等位基因(图6)。
2.2.7 GS9 基因型分析
GS9 是水稻9 号染色体上控制粒长的基因,编码一种没有已知保守功能域的蛋白质,是调控水稻粒型和外观品质的转录激活因子。ZHAO 等[11]根据多态性位点第2 外显子7 kb 的插入开发了InDel 分子标记。GS9短粒等位基因型材料无7 kb 片段的插入,PCR 可得到250 bp 左右的片段;gs9 长粒等位基因材料有7 kb 插入片段,不会产生250 bp 左右的片段。对产物进行琼脂糖电泳,检测结果表明产物都在250 bp 左右,说明7个长粒粳品种和日本晴都不含有7 kb 插入,判断其都不含有GS9 长粒基因(图7)。
2.2.8 GW5/qSW5 基因型分析
GW5 是位于5 号染色体上控制水稻粒宽的基因,窄粒水稻与宽粒水稻相比,发现宽粒品种核苷酸有段1 212 bp 的缺失。王鸣森[12]根据这个1 212 bp 差异的缺失设计了1 个InDel 标记。用该引物对检测水稻材料扩增,宽粒等位基因品种PCR 之后没有PCR 产物;窄粒等位基因扩增产物为775 bp 的片段。琼脂糖电泳检测结果表明,只有CLJ-HZ 含有1 212 bp 核苷酸的缺失,说明CLJ-HZ 含有GW5 宽粒基因。其它长粒粳品种和日本晴都不存在1 212 bp 核苷酸的缺失,说明都不含GW5 宽粒基因(图8)。
2.2.9 GW8 基因型分析
GW8 是位于8 号染色体上控制水稻粒宽的基因,裔传灯等[13]根据该基因启动子区10 bp 的InDel 和第3外显子A/C 和T/G 的2 个错义位点分别开发了功能标记(GW8-1)、dCAPs(GW8-2)和 CAPs(GW8-3),并依据这3 个变异位点将该基因分成8 种单倍型,Hap1、Hap2、Hap3、Hap7 的粒长相对于 Hap4、Hap5、Hap6 的较长。对于GW8-1 引物,若所检测水稻材料存在10 bp序列插入片段,则进行PCR 之后会产生151 bp 的片段,若不存在10 bp 序列插入片段则会产生141 bp 的片段。检测结果表明,鄂香2 号的3 个检测位点分别属于10 bp+、A、T 三种等位基因,属于粒宽的Hap3 单倍型;稻花香 2 号、CLJ-AH、CLJ-YZ、CLJ-HZ、CLJ-HB、CLJ-JX、日本晴的3 个检测位点分别属于10bp-、C、G三种等位基因,属于粒宽变小的Hap6 单倍型(图8)。
2.2.10 GS5 基因型分析
GS5 位于水稻5 号染色体上,是一个控制水稻粒宽、充实度和千粒重的数量性状基因,编码1 个丝氨酸羧肽酶;GS5 在第2 外显子的ACC/CTA 和第9 外显子A/C 的2 个变异位点对水稻籽粒的粒长、粒宽和长宽比性状存在显著差异。裔传灯等[14]根据这2 个位点分别设计2 个CAPS 标记GS5-1 和 GS5-2,依据这 2 个变异位点将该基因分成4 种单倍型,GS5-1 可以扩增出225 bp 的PCR 产物,经过DdeI 酶切后,能够被切成196 bp 的片段,基因型为CTA,不能被切的基因型为ACC,GS5-2 可以扩增出 188 bp 的 PCR 产物,经过 SalI酶切后,能够被切成169 bp 的片段,基因型为C,不能被切的基因型为A。以长粒的珍汕97B 作对照,结果显示,7 个长粒粳均出现225 bp 和188 bp 条带,对应变异位点为ACC 和A,和日本晴单倍型一致(图10)。
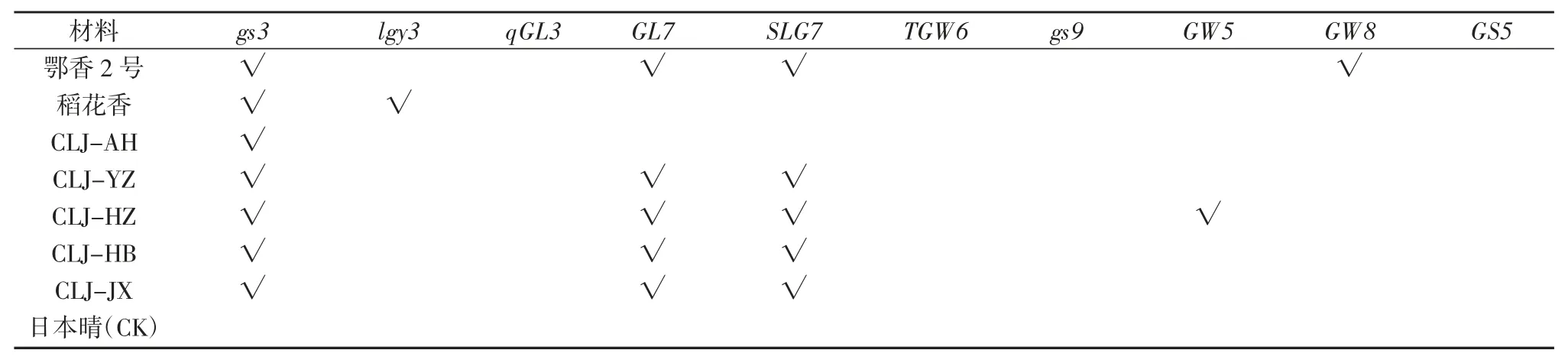
表3 7 个长粒粳品种粒型基因检测结果

图10 7 个长粒粳品种GS5 基因型分析
2.2.11 7 个长粒粳品种10 个粒形相关基因的基因型综合分析
检测结果表明,7 个长粒粳稻都含有gs3 长粒基因;鄂香 2 号、CLJ-YZ、CLJ-HZ、CLJ-HB、CLJ-JX 等 5个品种带有GL7、SLG7 长粒等位基因;稻花香2 号带有lgy3 长粒等位基因;鄂香2 号带有GW8 宽粒等位基因;CLJ-HZ 带有GW5 粒宽等位基因;被测水稻中均不含 qGL3、TGW6、gs9 长粒等位基因和 GS5 粒宽等位基因。
3 讨论
水稻的粒形性状与产量和品质密切相关,主要包括粒长、粒宽、粒厚、长宽比和长厚比等;粒长不仅决定稻米的外观品质和碾磨品质,而且对水稻的千粒重也有影响[1]。尽管粒长是数量性状,与环境条件有一定关系,但基因型仍是决定粒长的关键因素。一般来讲,粳稻的蒸煮食味品质要优于籼稻,而在粒型上,籼稻比粳稻要长,外观品质更佳,市场竞争力强。长粒大米受到消费者的喜爱,育种家们根据经验在田间选育出了长粒粳稻品种,但是其中的遗传和分子机制还不清楚。未来的粳稻育种需要着重改良粒型,利用已经克隆的粒长相关基因,开展分子辅助育种,并应用于育种实践,有助于水稻单产的提高和稻米品质的改良。
到目前为止,水稻中共定位到100 多个与粒长相关的QTLs,大多数定位结果显示,控制粒长的主效QTLs 位于第3 染色体和第7 染色体[16]。对位于第3 染色体上的粒长主效基因,一般认为是GS3 和qGL3 基因。但丁丹等[17]研究认为,在这2 个基因都存在时,并未表现出基因累加效应,而只有其中1 个基因存在时,才表现出较强的表型效应。由于qGL3 基因分布较少,所以第3 染色体上控制粒长的基因主要是GS3,因此,更多研究也集中在GS3 基因上。张磊[18]利用秋B 携带的GS3 基因对博B 的粒长进行了改良,获得了长谷粒型的博B。张剑霞等[19]用MAS 技术将GS3 导入到珍汕97B 和II-32B 中,使珍汕97B 的粒长由原来的8.0 mm增加到9.8 mm,II-32B 粒长由原来8.0 mm 增加到9.7 mm,粒长改良效果明显。杨梯丰[20]利用携带GS3 基因的单片段代换系进行长粒稻育种,有效改良了华粳籼74 的外观品质。WANG 等[21]分别将 9311 的 GS3 导入到珍汕97B 中,不仅使珍汕97B 的粒长增加了0.9 mm,达到9.2 mm,千粒重也有所增加,改良后的珍汕97B 不仅外观品质更好,产量也有所提高。这些gs3 基因在籼稻粒形改良上的应用为粳稻粒形改良提供了借鉴。
在长粒粳稻研究中,孟帅等[22]利用CRISPR/Cas9技术敲除GS3 基因,增加了11 个粳稻品种的粒长,进而改善了花时,为杂交粳稻育种提供了一个新的思路。LIU 等[6]发现,lgy3 和gs3 等位基因的聚合提高了总籽粒产量(约7%),且改善了稻米品质,为同时选育高产优质水稻提供了一种新的策略。从本研究可以看出,gs3 基因在长粒粳稻传统经验育种中已得到了广泛应用,7 个长粒粳稻品种都含有gs3 基因,分析原因除了gs3 基因是主效基因外,还在于它的亲本中分布较广,热带粳稻和长粒籼稻均含有gs3 基因。本研究发现,7个优质长粒粳稻品种中有5 个含有GL7 和SLG7 基因,说明这2 个基因在长粒粳稻育种中有较好的应用前景。7 个长粒粳稻品种中未发现qGL3、TGW6、gs9 的长粒等位基因,可能与这些基因在水稻资源中分布频率较低有关,利用分子标记技术进行定向选择,有利于这些基因在育种中的利用。
