黑曲霉和酵母菌联合固态发酵凉茶渣提高酶活力的工艺研究
2020-12-05袁明贵徐志宏彭新宇余丹妮周廷斤唐兴刚
袁明贵 徐志宏 彭新宇 余丹妮 田 雅 周廷斤 唐兴刚 向 蓉*
(1.广东省农业科学院动物卫生研究所 广东省畜禽疫病防治研究重点实验室农业农村部兽用药物与诊断技术广东科学观测实验站广东省中兽药工程技术研究中心,广东广州510640;2.广东省前沿动物保健有限公司,广东广州510640)
中国主流的饮用性凉茶是由鸡蛋花、金银花、菊花、凉粉草、夏枯草、甘草和布渣叶共7种植物药物煎制而成。由于凉茶的提取过程更注重风味成分的保留,造成凉茶渣仍具有与原药材相似的营养成分[1],是一种未被充分利用的生物资源。然而对于凉茶渣的资源化应用研究比较缺乏,仅在直接作为饲料添加剂[1]或生物质能源[2]方面有少量的报道。
凉茶渣中仍含有较高的粗蛋白质、粗脂肪和粗灰分等营养成分,可用作饲料添加剂[1]。但细胞壁中的纤维素、木聚糖和果胶阻碍了动物对营养的消化利用。内切葡聚糖酶是纤维素酶的重要成分之一,可以从纤维素分子内部水解糖苷键[3-4],木聚糖酶[5-7]和果胶酶[8-9]可以分别高效降解抗营养因子木聚糖和果胶,生成单糖或寡糖,从而促进植物细胞壁的降解,三者在饲料和食品工业中都有广泛的应用[10-12]。由于木聚糖酶和果胶酶的存在能够更好地促进纤维素酶降解底物[13],因此发酵产物中不进行分离的纤维素酶、木聚糖酶和果胶酶更能促进菌株转化利用植物细胞[14]。黑曲霉是一种常见的无毒性曲霉属真菌[15],可以产生多达21种生物酶类[16-17]。酵母菌是工农业最常用菌种之一,和黑曲霉共同发酵时具有良好的协同作用[18-20]。本文拟通过单因素实验和正交实验,探索利用黑曲霉和产朊假丝酵母联合固态发酵凉茶渣生产内切纤维素酶、木聚糖酶和果胶酶的最佳工艺,为凉茶渣在饲料中的资源化利用提供参考。
1 材料与方法
1.1 材料与仪器
黑曲霉(GIM3.562)、产朊假丝酵母(GIM2.148),购于广东省微生物菌种保藏中心;凉茶渣由国内某一大型食品饮料公司提供。
5804R高速冷冻离心机(Eppendorf公司),XS205DU电子天平[梅特勒-托利多仪器(上海)公司],LDZX-50KB 立式高压灭菌器(上海申安医疗器械厂),Elx50酶标仪(BioTek Instruments 公司),BME生物显微镜(上海莱卡公司)。
1.2 培养基
马铃薯葡萄糖琼脂培养基(PDA 培养基):马铃薯200 g去皮,切成块,煮沸30 min,纱布过滤,加入葡萄糖20 g、琼脂20 g,补水至1 000 ml,然后121 ℃湿热灭菌20 min,趁热制备菌种培养基。
固态凉茶渣培养基:将新鲜凉茶渣晒干粉碎,取粉末2 g,加入硫酸铵0.08 g、葡萄糖0.04 g、磷酸二氢钾0.01 g、磷酸氢二钾0.008 g、水8 g,121 ℃湿热灭菌,灭菌后pH值为5.0。
1.3 实验方法
在80%湿度,30 ℃的恒温恒湿培养箱中,利用PDA 平板培养基将黑曲霉培养120 h,产朊假丝酵母培养24 h,两种菌种都连续活化两次。制备黑曲霉孢子和产朊假丝酵母菌种悬液,利用血球板计数法,测定各菌种浓度,混合菌种悬液,使每种菌种浓度都为2×109CFU/ml,将菌种按照10%接种量接种至固态凉茶渣培养基中,再根据各部分实验设计进行培养。
1.3.1 单因素实验
分别考察发酵时间(48~144 h)、温度(28~37 ℃及室温)、含水量(65%~85%)、浸泡液pH值(5.0~9.0)、氮源种类(尿素、氯化铵、硫酸铵、豆粕和不补充氮源)和碳源种类(蔗糖、葡萄糖、糖蜜、α-乳糖和不补充碳源)共6 个因素对内切葡聚糖酶、果胶酶活力和木聚糖酶活力的影响。文中硫酸铵含量、葡萄糖含量、含水量以及接种量均按凉茶渣的干重计算。
1.3.2 正交实验
根据单因素实验结果,以硫酸铵为氮源,葡萄糖为碳源,对含水量(A,60%、70%、80%)、发酵温度(B,31、34、37 ℃)、浸泡液pH 值(C,7.0、8.0、9.0)、发酵时间(D,72、120、168 h)四个因素进行三水平正交实验优化(表1)。
1.3.3 酶活力测试
发酵结束后,将发酵培养物加入15 ml生理盐水,震荡后静置过夜;经过5 000 r/min 离心10 min,采用3,5-二硝基水杨酸(3,5-dinitrosalicylic acid, DNS)显色法检测上清液中内切葡聚糖酶、木聚糖酶和果胶酶活力[13,21-22],沉淀烘干后质量即为发酵样品干重。
1.3.3.1 酶活力测试方法
取底物溶液0.4 ml,发酵上清液0.1 ml,混匀后,40 ℃水浴进行酶促反应10 min;然后加入DNS 0.5 ml,再次混匀后,煮沸10 min,以蒸馏水代替底物溶液为对照,测定特定波长吸光度,计算酶活力。
用0.2 mol/l的乙酸-乙酸钠缓冲溶液(pH=5.0),配制6.25 mg/ml羧甲基纤维素钠溶液为底物,以葡萄糖为标准物质,测定530 nm 的吸光度,制定标准曲线并计算内切葡聚糖酶活力[13];用0.2 mol/l 的磷酸氢二钠-0.1 mol/l柠檬酸缓冲溶液(pH=5.0),配制0.5%的果胶溶液为底物,以D-半乳糖醛酸为标准物质,测定540 nm的吸光度,制定标准曲线并计算果胶酶活力[21];用0.1 mol/l 的磷酸氢二钠-0.1 mol/l 柠檬酸缓冲溶液(pH=5.0),配制0.5%的木聚糖溶液(pH=5.0)为底物,以木糖为标准物质,测定550 nm的吸光度,制定标准曲线并计算木聚糖酶活力[22]。
1.3.3.2 酶活力计算
规定40 ℃时,每分钟产生1 μmol 产物的酶量为1个酶活力单位(U)。
式中:C0——对照的还原糖浓度(μmol/ml);
C1——酶促反应后还原糖浓度(μmol/ml);
V1——参与反应的酶溶液体积(ml);
V2——酶溶液总体积(ml);
t——酶促反应时间(min);
m——发酵样品干重(g)。
1.4 数据处理
采用SPSS 21 软件对单因素实验中的同种酶活力进行方差分析;并利用该软件进行正交实验设计及方差分析,以P<0.05作为差异显著标准。所有实验都进行3次平行实验。
2 结果与分析
2.1 单因素实验
2.1.1 发酵时长对酶活力的影响
向凉茶渣补充4%硫酸铵,2%葡萄糖,温度为31 ℃,含水量为80%,考察发酵时长在48~144 h内变化对内切葡聚糖酶、果胶酶和木聚糖酶活力的影响。实验结果(见图1)显示,三种酶活力均随着发酵时间的延长而增加;超过72 h后内切葡聚糖酶和果胶酶的活力变化不显著;木聚糖酶活力在96 h和120 h时无显著差异。

图1 发酵时长对酶活力的影响
时间对发酵效果具有显著影响。如果发酵时间太短,菌体生长没有达到稳定期,使发酵不完全;发酵时间过长,营养供应不足、菌种老化、产生有害的次级代谢产物,甚至造成菌体自溶[23]。经过化学诱变选育的黑曲霉X1U4-1 在发酵玉米芯和麸皮(3:7)混合培养基时,纤维素酶活力84 h 时达到最大值,木聚糖酶活力68 h时达到最大值[16]。在利用黑曲霉GJ-2和酒精酵母J-1发酵橙子皮和麦麸生产果胶酶时,当菌种同时接入发酵60 h 后,果胶酶活力不再显著增加[19]。综合考虑规模化处理效率,将后续的单因素实验的时间设定为72 h。
2.1.2 温度对酶活力的影响
向凉茶渣补充4%硫酸铵,2%葡萄糖,含水量为80%,发酵72 h,考察温度在28~37 ℃范围内以及室温条件下,对三种酶活力的影响。从实验结果(见图2)可以发现,温度对木聚糖酶活力无显著影响;而在34 ℃时的内切葡聚糖酶活力显著高于28 ℃时的活力;温度在28~37 ℃范围内变化时,对果胶酶的活力无显著影响。本实验是在广州市的8月下旬进行,室温较高,适合黑曲霉与酵母菌的生长,因此凉茶渣室温发酵时,三种酶活力也较强。尽管28 ℃发酵时,果胶酶活力较高,但是31 ℃时内切葡聚糖酶活力和木聚糖酶活力相对较高,考虑到南方气温偏高,并且黑曲霉在发酵过程中存在放热现象,因此将后续单因素实验的温度设定为31 ℃。在利用黑曲霉和产朊假丝酵母发酵甜高粱酒糟和麦麸生产蛋白饲料,35 ℃时纤维素酶活力最大[24],与本部分的内切葡聚糖酶活力最大值对应温度基本一致。

图2 温度对酶活力的影响
2.1.3 含水量对酶活力的影响
含水量是指当体系仅含有凉茶渣和水时,水的百分含量。在本部分实验中,向凉茶渣补充4%硫酸铵、2%葡萄糖,含水量在65%~85%范围内变化,31 ℃发酵72 h,考察含水量对三种酶活力的影响。实验结果(见图3)显示,当含水量从65%增加到85%过程中,内切葡聚糖酶活力和果胶酶活力基本上逐渐降低,其中80%和85%时的内切葡聚糖酶活力显著低于65%的内切葡聚糖酶活力;80%和85%时的果胶酶活力显著低于65%~75%的果胶酶活力;含水量对木聚糖酶活力没有显著影响。
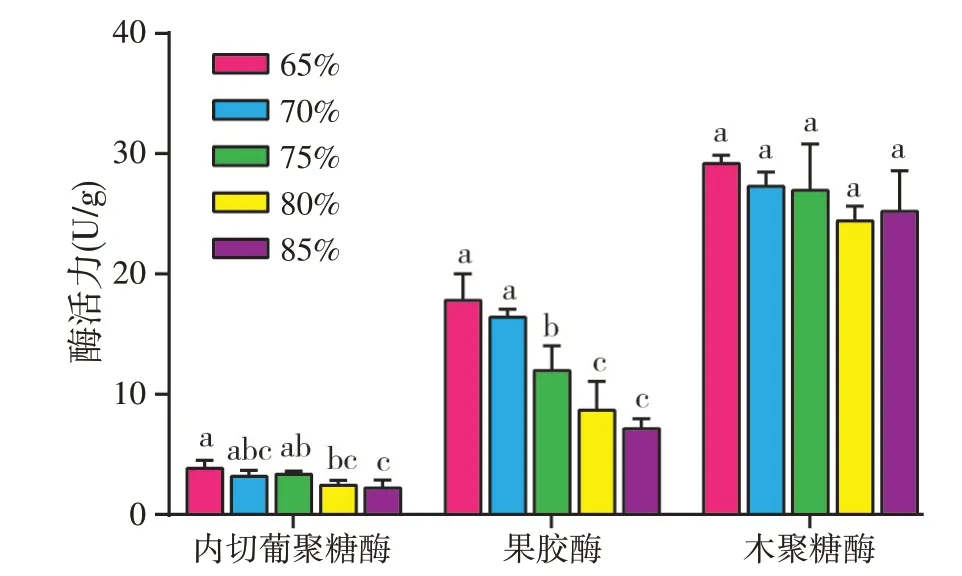
图3 含水量对酶活力的影响
基质的含水量影响着胞外酶的分泌和转移,影响氧的传递以及菌体生长。如果起始含水量太低,使菌种正常生长受到限制;含水量过高,基质密度过大,透气性和传热性较差,也不利于菌种生长。一般来说真菌发酵时需要相对更低的含水量,如利用黑曲霉和产朊假丝酵母发酵三七渣生产蛋白饲料时,最优含水量为70%[25];在利用黑曲霉HN-1 发酵豌豆荚生产纤维素酶时,发现含水量超过65%以后,纤维素酶活力显著降低[17]。本实验也发现当含水量在65%~85%之间时,三种酶活力基本上随着含水量的增加而降低。由于新鲜凉茶渣的含水量为79.45%[1],接近80%,而本研究的目的是为凉茶渣的资源化利用提供参考,因此将后续的单因素实验含水量均设定为80%。
2.1.4 浸泡液pH值对酶活力的影响
首先向计算量60%的蒸馏水中加入与其他实验PO43-等物质的量的磷酸,利用3.0 mol/l的氢氧化钾逐渐上调pH 值,形成pH 值分别为5.0、6.0、7.0、8.0、9.0的低浓度磷酸盐缓冲体系,然后加入干凉茶渣进行浸泡,再补充4%硫酸铵和2%葡萄糖,将每份样品含水量均调节为80%后,31 ℃发酵72 h,考察浸泡液pH值对三种酶活力的影响。结果发现,pH 值对内切葡聚糖酶活力影响不显著;在pH 值为9.0 时,木聚糖酶和果胶酶活力显著高于pH值为7.0时的酶活力(见图4)。事实上,凉茶渣自身pH值为6.1,且具有一定pH值缓冲能力。当用pH 值为5.0、6.0、7.0、8.0、9.0 的浸泡液浸泡凉茶渣,pH 值实测值分别为5.6、5.7、5.8、6.1 和7.2。尽管7.0 是浸泡液pH 值考察范围的中间值,但是,可能不是菌种最适宜生长环境或产酶环境,导致果胶酶活力和木聚糖酶活力显著降低。考虑到规模化处理时,尽量简化操作步骤,在后续的单因素实验中依然向凉茶渣中补充0.5%磷酸二氢钾和0.4%磷酸氢二钾,使起始pH值为5.0。

图4 浸泡液pH值对酶活力的影响
2.1.5 氮源种类对酶活力的影响
以2%葡萄糖为碳源,含水量为80%,向凉茶渣补充4%尿素、氯化铵、硫酸铵、豆粕四种氮源以及不补充氮源,31 ℃发酵72 h,考察氮源种类对三种酶活力的影响(见图5)。结果显示,与不补充氮源相比,尿素显著提高了内切葡聚糖酶活力,硫酸铵和氯化铵显著地提高了内切葡聚糖酶活力;氯化铵显著降低了果胶酶活力;尿素、氯化铵和硫酸铵显著提高木聚糖酶活力。实验结果表明,无机氮源比豆粕能够更好地促进凉茶渣发酵生产内切葡聚糖酶和木聚糖酶。在利用黑曲霉SD-03发酵玉米芯生产木聚糖酶时,发现多种无机氮源的发酵效果普遍优于有机氮源[26]。绝大多数微生物都需要氮源合成蛋白质、核酸等物质,当氮源不足时,不利于微生物生长增殖。由于凉茶渣中氮源比较缺乏,因此不加任何氮源时,内切葡聚糖酶和木聚糖酶活力相对较低。综合考虑四种氮源和不补充氮源对三种酶活力的影响效果,在后续实验中选择向凉茶渣中补充4%硫酸铵作为氮源。

图5 氮源种类对酶活力的影响
2.1.6 碳源对酶活力的影响
含水量为80%,以硫酸铵为氮源,向凉茶渣补充4%蔗糖、葡萄糖、糖蜜、α-乳糖以及不补充碳源,31 ℃发酵72 h,考察碳源种类对三种酶活力的影响(见图6)。从实验结果可以看出四种碳源和不补充碳源对内切葡聚糖酶活力无显著影响;与不补充碳源和α-乳糖相比,蔗糖、葡萄糖和糖蜜显著降低了果胶酶活力和木聚糖酶活力;不补充碳源和α-乳糖对果胶酶和木聚糖酶活力影响无显著差异。由于黑曲霉发酵时产生的生物酶种类丰富,可以降解淀粉、纤维素等多糖生成单糖或寡糖,供自身生长和增殖需要,因此,利用黑曲霉发酵中药渣时,可以不补充碳源[27-29]。本实验发现了不补充碳源时,黑曲霉和产朊假丝酵母发酵凉茶渣对纤维素酶活力和木聚糖酶活力无显著影响,而且果胶酶活力优于补充蔗糖、葡萄糖和糖蜜时的酶活力。
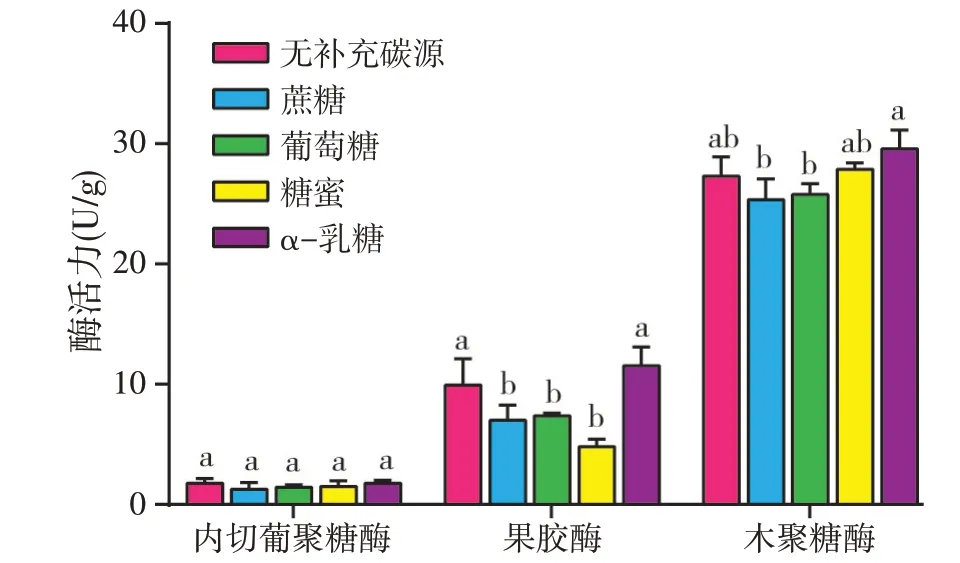
图6 碳源种类对酶活力的影响
2.2 正交实验
尽管在发酵72 h的单因素实验中,发现不补充碳源时内切葡聚糖酶活力和木聚糖酶活力与补充其他碳源时无显著差异,且果胶酶活力显著高于添加蔗糖、葡萄糖和糖蜜时的活力,但是考虑到在正交实验中,随时间的延长,可能会对产酶具有较强的副作用,且考虑成本因素,未选择酶活力较高的α-乳糖,而选择添加2%的葡萄糖作为正交实验中碳源;并且在单因素实验中发现,当发酵时间越长,含水量越低时,酶活力越高,因此在正交实验中将发酵时间延长至168 h,含水量降低至60%,探索更宽的时间和含水量范围对三种酶活力的影响。同时,向凉茶渣中补充4%硫酸铵作为氮源,对含水量(A)、浸泡液pH 值(B)、温度(C)以及发酵时间(D)四个因素进行三水平正交实验优化,结果见表1。
对表1 中的三种酶活力分别进行多因素方差分析,结果见表2。从表2可以发现,当对内切葡聚糖酶活力进行正交分析时,发现各因素对发酵效果的影响顺序为时长>含水量>浸泡液pH值>温度,其中时长对内切葡聚糖酶活力具有显著影响;最优工艺为含水量为60%、浸泡液pH值为7.0、温度为37 ℃、发酵120 h,即A1B1C3D2。
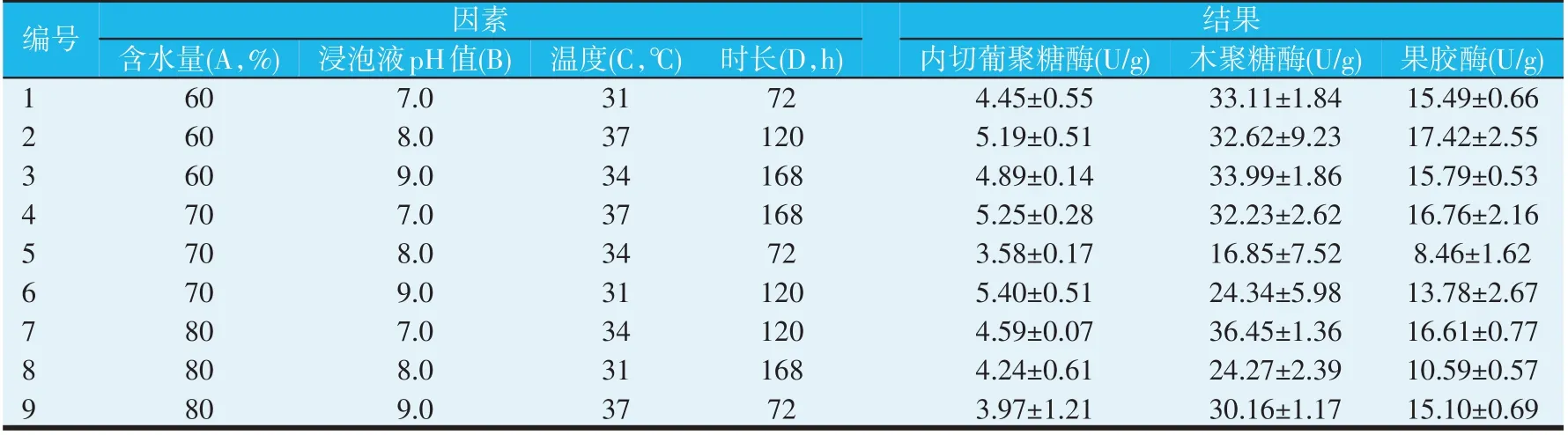
表1 凉茶渣固态发酵正交实验L9(34)

表2 正交实验的方差分析
当对木聚糖酶活力进行正交分析时,发现各因素对发酵效果的影响顺序为浸泡液pH 值>含水量>时长>温度,其中浸泡液pH值和含水量对木聚糖酶活力具有显著影响;最优工艺与产内切葡聚糖酶的最优工艺相同,也为含水量为60%、浸泡液pH值为7.0、发酵温度为37 ℃、发酵120 h,即A1B1C3D2。
当对果胶酶活力进行正交分析时,发现各因素对发酵效果的影响顺序为温度>时长>浸泡液pH 值>含水量,其中温度和时长对木聚糖酶活力具有显著影响;最优工艺为含水量为60%、浸泡液pH值为9.0、发酵温度为37 ℃、发酵120 h,即A1B3C3D2。
2.3 验证实验
由于浸泡液pH 值对果胶酶活力影响不显著(P=0.087),且pH 值在7.0 和9.0 水平时,果胶酶活力分别为10.171 U/g 和10.426 U/g,差异不显著,因此对果胶酶活力的正交分析结论进行验证时,采用与内切葡聚糖酶活力和木聚糖酶活力相同的工艺。经过验证,含水量为60%,浸泡液pH 值为7.0,37 ℃发酵120 h(即A1B1C3D2),发酵产物中内切葡聚糖酶、木聚糖酶、果胶酶均获得较高的活力,分别为(5.55±0.77)、(37.32±2.26)、(29.51±3.23) U/g,与正交实验结论相符。
3 讨论
单一菌种进行发酵时,酶系往往不完全,而且发酵产物对酶具有反馈抑制作用,因此,选择具有协同作用的菌种进行联合发酵是持续深入发酵的关键。谭显东等[28]利用正交实验筛选发酵三七渣生产蛋白的最佳菌种时,发现黑曲霉与产朊假丝酵母联合发酵效果显著优于其他菌种组合,通过方差分析,发现黑曲霉和酵母菌,以及二者的相互作用对蛋白质含量均有显著影响。岳丽等[30]发现,利用黑曲霉和产朊假丝酵母联合发酵甜高粱酒糟生产蛋白时,与单一菌种相比,蛋白质含量显著提高,纤维素含量显著降低。黑曲霉发酵时可以分泌如纤维素酶等多种酶类,降解大分子糖类化合物生成单糖,过多的单糖对酶产生抑制作用,而单糖是酵母菌的优质碳源,这样利用黑曲霉和酵母菌联合发酵,可以相互促进,产生协同作用。黑曲霉和产朊假丝酵母的协同作用是本研究的基础。
岳丽等[24]利用黑曲霉和产朊假丝酵母联合发酵甜高粱酒糟和麦麸生产蛋白饲料,经过响应面优化,31 ℃发酵6.25 d 后,纤维素酶活力达到1.33 U/g。本研究利用黑曲霉和产朊假丝酵母对凉茶渣进行固态发酵,在最优条件下内切葡聚糖酶活力为(5.55±0.77) U/g。二者的区别主要是由于前者以滤纸为底物,测试的是纤维素酶的整体活力,而本研究主要是以纤维素钠为底物,测试的是内切葡聚糖酶的活力;同时发酵菌种、基质和条件的不同,使二者差异较大。周杰民等[19]利用黑曲霉GJ-2 和酒精酵母J-1 发酵橙子皮和麦麸生产果胶酶,经过条件优化后,果胶酶活力达到188.01 U/ml,而本研究的果胶酶活力为(29.51±3.23) U/g;张水龙[31]利用黑曲霉CICC40616 发酵含有豆粕和木聚糖的培养基,经优化后木聚糖酶活力为82.43 U/ml,而本研究的木聚糖酶活力为(37.32±2.26) U/g。可能是这两个实验是以生产果胶酶活力或木聚糖酶为目的,选择了产目标酶的专一菌株,而本文以黑曲霉和产朊假丝酵母固态发酵凉茶渣,同时产生内切葡聚糖酶、木聚糖酶和果胶酶,以期三者能够协调高效地降解植物细胞壁。
4 结论
本文利用黑曲霉和产朊假丝酵母的协同作用,对凉茶渣进行固态发酵,通过单因素实验和正交实验,发现含水量为60%、浸泡液pH值为7.0、温度为37 ℃、发酵120 h,是生产内切葡聚糖酶、木聚糖酶、果胶酶的较合适工艺,在此工艺下,三种酶活力均较高,分别为(5.55±0.77)、(37.32±2.26)、(29.51±3.23) U/g。本研究为凉茶渣在饲料中的资源化利用提供了参考。
