橡胶树HbCPA基因的克隆、表达及生物信息学分析
2020-12-04杨洪胡燕玲岳镒繁邓治代龙军李德军
杨洪 胡燕玲,2 岳镒繁,2 邓治 代龙军 李德军
(1.中国热带农业科学院橡胶研究所 农业农村部橡胶树生物学与遗传资源利用重点实验室 省部共建国家重点实验室培育基地-海南省热带作物栽培生理学重点实验室,海口 571101;2.南京农业大学农学院,南京 210095)
橡胶是四大工业原料之一,根据来源不同分为天然橡胶和合成橡胶。天然橡胶在弹性、耐磨性和延展性等特性方面具有合成橡胶不可替代的优势,在一些重要工业领域(如航空、航天和重型汽车等制造业)广泛应用。我国是天然橡胶第一大消费国和进口国,自2017年我国天然橡胶年消费量和进口量均在5.0×106t以上,而我国天然橡胶年产量持续在8.0×105t上下波动,供需缺口悬殊,需要通过进口来填补国内天然橡胶资源短缺。巴西橡胶树(Hevea brasiliensis,简称橡胶树)是典型的热带雨林乔木,是天然橡胶的主要来源[1]。天然橡胶产业是典型的环境约束型产业,受自然因素约束我国适宜种植橡胶树的地区主要集中在海南、广东、云南三地。截至2018年末,我国天然橡胶种植面积约为1.15×106hm2。目前,我国天然橡胶产业面临植胶区域有限且种植趋于饱和及天然橡胶价格持续低迷等问题,已很难通过扩大种植面积来增加产量。我国地处热带北缘,橡胶树在生长周期中不可避免地会遭受低温寒害、台风和季节性干旱等气候影响。不良的气候环境严重影响橡胶树单位面积产量,缩短橡胶树经济寿命。因此,提高橡胶树单位面积产量及其对逆境胁迫的耐受能力是缓解天然橡胶供需平衡压力的最佳途径。
多胺(Polyamine,PAs)是一类小分子脂肪族含氮多聚物,几乎存在于所有的生物体。在植物中多胺主要以腐胺(Putrescine,Put)、亚精胺(Spermidine,Spd)和精胺(Spermine,Spm)的形式存在,其中Put是多胺生物合成的中心产物[2-4]。Put的合成涉及两条途径,一条为通过鸟氨酸脱羧酶(Ornithine decarboxylases,ODC)催化鸟氨酸脱羧形成腐胺的简单反应,称为鸟氨酸途径;另一条以精氨酸为起始物质,因此称为精氨酸途径。精氨酸经精氨酸脱羧酶(Arginine decarboxylase,ADC)脱羧形成鲱精胺(Agmatine,Agm),鲱精胺在鲱精胺亚氨水解酶(Agmatine iminohydrolase/Agmatine deiminase,AIH)的催化下水解生成N-氨甲基腐胺(N-carbamoylputrescine,NCP),最后在N-氨甲酰基腐胺酰胺水解酶(N-carbamoylputreseine amidohydrolase,CPA)的催化水解下生成腐胺[5-7]。植物是唯一能够通过精氨酸途径合成Put的真核生物,该途径是某些植物(如拟南芥、小立碗藓)腐胺合成的唯一途径[8]。
在农业生产中,常通过外源施加PAs来提高作物产量和增强作物耐逆性。例如,外源施加Put能够提高小麦的耐热性[9],减轻苹果愈伤组织盐胁迫损伤[10]。利用转基因技术提高PAs合成相关基因的表达能使内源PAs在生物体内积累,从而达到作物抗逆、增产的目的[11-15]。Put是橡胶树胶乳中的主要PAs,割胶能够促进其生物合成[16]。目前,关于橡胶树PAs合成相关基因的研究仅限于SAMDC、ADC、AIH等少数几个基因的克隆及功能分析[17-19]。为进一步丰富和了解橡胶树PAs合成途径相关基因的生物学功能及表达特性,本文从橡胶树品系热研7-33-97中克隆了首个CPA基因并对其进行生物信息学及表达特性分析。
1 材料与方法
1.1 材料
本文所用材料均为橡胶树品系热研7-33-97。健康树树皮、叶片、茎尖、雌花、雄花、胶乳等组织样品及死皮橡胶树胶乳、树皮样品均采自中国热带农业科学院试验农场;不同发育阶段叶片样品采自国家橡胶树种质资源圃(儋州);过氧化氢(H2O2)、乙烯利(Ethephon,ET)和茉莉酸甲酯(Methyl jasmonate,MeJA)处理的胶乳样品均来自于已达到开割标准的未开割橡胶树;低温、干旱、高盐胁迫处理的叶片样品取自移栽6个月且长势基本一致的橡胶树组培苗,处理方法参照杨洪等[19]方法。
1.2 方法
1.2.1 总RNA提取及cDNA第一链合成 用北京百泰克生物技术有限公司生产的通用植物总RNA提取试剂盒,按其使用说明书提取样品总RNA,分光光度法检测RNA浓度,琼脂糖凝胶电泳进行完整性检测。参照PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒说明书进行样品cDNA合成。
1.2.2 HbCPA全长序列克隆 进行橡胶树转录组测序结果分析时我们获得了一条与植物CPA基因序列高度同源的unigene,通过NCBI的blast比对确定其为CPA基因。由于该unigene缺少阅读框3'末端序列,因此采用SMARTerTMRACE cDNA Amplification Kit(Clontech,USA)试剂盒进行3'-RACE扩增。3'-RACE所用引物为5'-CACATTTTATGGGAACTCATTC-3'和5'-CAAATCCAAGAGACATAGTTGG-3'。按照试剂盒说明书用3'-RACE CDS Primer A对1 μg各组织混合RNA进行反转录,获得用于3'-RACE的模板cDNA。以获得的3'-RACE cDNA为模板用PrimeSTAR Max Premix(TaKaRa,China)进行3'端序列扩增。将获得的3'端序列与转录组中获得的unigene序列进行拼接,根据拼接序列并设计全长cDNA扩增引物,进行PCR和测序验证。
1.2.3 HbCPA生物信息学及进化分析 用NCBI的ORF finder预测开放阅读框;用NCBI保守结构域数据库网站(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测橡胶树CPA蛋白保守结构域;用SMART(http://smart.embl-heidelberg.de/)分析蛋白保守结构域;用Expasy网站的Prot-Param程序(http://web.expasy.org/protparam/)分析蛋白质理化性质;用NCBI中的BLAST和DNASTAR软件进行同源性分析;用SignalP 4.1Server 软件(http://www.cbs.dtu.dk/services/SignalP/)进行信号肽预测分析;用TMHMM 2.0程序(http://www.cbs.dtu.dk/services/TMHMM/)预测跨膜结构域;利用在线工具(https://wolfpsort.hgc.jp)分析目的序列的亚细胞定位;用ClustalX和MEGA6.0进行多序列比对和Neighbor-Joining进化树的构建,用多序列比对显色工具ESPript对多序列比对显示美化[20]。
1.2.4 实时荧光定量PCR 根据CPA序列设计特异性荧光定量引物(F:5'-GACGTCCAGATTTGTACAAGG-3'和R:5'-CCACAAAAAATTCATGGTTGGG-3')。以橡胶树18S rRNA基因为内参基因(F:5'-GCTCGAAGACGATCAGATACC-3'和R:5'-TTCAGCCTTGCGACCATAC-3')。以不同样品反转录cDNA稀释5倍后为模板,反应体系为20 μL,包含2 μL模板,10 μL 2×SYBR Premix、10 μmol/L上游引物和下游引物各1 μL、灭菌水补足至20 μL。qPCR在Bio-Rad CFX96 qPCR仪进行,程序设置为:95℃预变性5 min;95℃ 10 s,58℃ 30 s,72℃ 20 s,40个循环后进行熔解曲线分析,以确定引物的特异性。基因相对表达量采用公式2-ΔΔCt计算。
2 结果
2.1 HbCPA的克隆及序列分析
通过对橡胶树转录组测序结果分析,获得了一条长度为1 068 bp的unigene。NCBI在线blast比对显示该unigene与高度植物CPA基因序列相似性均在85%以上,推测其为橡胶树CPA基因。由于该unigene缺少阅读框3'末端序列,因此进行了3'-RACE扩增。经过测序、拼接及RT-PCR验证,获得具有完整阅读框的橡胶树CPA基因全长序列,并将其命名为HbCPA。HbCPA序列全长1 342 bp,开放阅读框906 bp,编码301个氨基酸(图1)。
HbCPA预测理论分子量约为33.48 kD,理论等电点为6.01。不稳定系数为33.67,为稳定蛋白。信号肽预测结果表明,HbCPA不含信号肽,为胞内蛋白。疏水性分析表明,该蛋白亲水性氨基酸残基为62个,占20.60%;疏水性氨基酸残基为113个,占37.54%,平均疏水系数为-0.269,为亲水性蛋白。氨基酸组成中带负电荷的氨基酸(Asp+Glu)共有40个,占13.29%;带正电荷的氨基酸(Arg+Lys)共有35个,占11.63%。跨膜结构域预测结果表明,HbCPA不含跨膜结构域,为非膜蛋白。亚细胞定位预测结果表明,HbCPA定位于细胞质。
2.2 HbCPA同源比对及二级结构特征
氨基酸序列比对结果表明CPAs在不同植物中高度相似,本研究所选物种的CPAs与HbCPA的一致性均大于87%。HbCPA与木薯MeCPA氨基酸序列一致性最高,达98.01%;其次是克莱门氏小柑橘CcCPA和大戟科的麻风树JcCPA,一致性均为92.36%。二级结构预测结果表明无规则卷曲为HbCPA二级结构的主要构成元件,占39.87%;其次是α-螺旋和β-片层结构,占比分别为29.90%和23.36%;β-转角结构最少,仅占6.98%(图2)。
为了分析HbCPA与其他植物CPA蛋白的亲缘关系,选取与HbCPA同源的15个不同植物CPAs进行系统进化分析。结果(图3)表明,橡胶树HbCPA先与同属大戟科的木薯MeCPA、蓖麻RcCPA和麻风树JcCPA聚为一枝,然后再与胡杨PeCPA和毛白杨PtCPA聚为一枝,表明它们有较近
2.3 HbCPA系统进化分析
的亲缘关系。而蒺藜苜蓿MtCPA、大豆GmCPA和水稻OsCPA聚为较远的一枝,暗示它们与HbCPA的亲缘关系较远。

图1 HbCPA cDNA序列及其编码的氨基酸序列
2.4 HbCPA组织表达特性及对橡胶树死皮的响应
实时荧光定量PCR结果(图4-A)显示,HbCPA在橡胶树的不同组织(树皮、叶片、茎尖、雌花和雄花)均有较高的表达量,但其表达存在组织差异。表达量从高到低依次最高为树皮、叶片、雌花、雄花、茎尖。叶片不同发育时期HbCPA表达结果(图4-B)表明,淡绿期HbCPA的表达量远高于其他4个时期,变色期和稳定期表达量相当,古铜期和衰老期表达量相当,但变色期和稳定期的表达量高于古铜期和衰老期。比较健康和死皮橡胶树中表达差异发现(图4-C),HbCPA在死皮橡胶树树皮中表达明显上升,而在胶乳中HbCPA的表达基本一致。
2.5 HbCPA在逆境胁迫及激素处理的表达模式
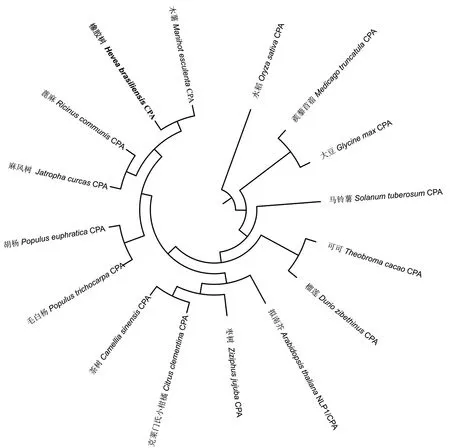
图3 HbCPA与其它物种CPAs蛋白系统进化分析

图4 HbCPA在橡胶树不同组织(A)、叶片不同发育时期(B)及健康/死皮橡胶树胶乳和树皮(C)表达模式
橡胶树胶乳中HbCPA的表达还受乙烯利(ET)、茉莉酸甲酯(MeJA)等处理和低温、干旱、高盐和H2O2等逆境条件的影响。H2O2处理后HbCPA表达先下降再上升,处理24 h表达量最低,至48 h表达量略有上升但也低于处理前水平(图5-A)。ET处理后胶乳中HbCPA表达呈现波动变化,处理后各时间点表达量均低于处理前水平。ET处理4 h表达量急剧下降至8 h表达量达到最低水平,24 h开始表达量开始上调,72 h开始下降至24 h的表达水平(图5-B)。MeJA处理4 h HbCPA表达急剧下降,处理8 h后逐渐升高直至处理48 h,然后又开始下降至处理72 h其表达水平与24 h基本一致(图5-C)。低温处理抑制HbCPA的表达,并且随着处理时间延长抑制作用越明显(图5-D)。胶乳中HbCPA的表达还受干旱和高盐胁迫调控,且表达趋势基本一致(图5-E,F)。干旱或高盐处理24 h,HbCPA表达达到最高水平,分别为处理前的3.3倍和4.4倍;处理48 h表达水平明显下降,但仍略高于处理前水平(干旱)或与处理前一致(高盐)。

图5 不同处理条件下HbCPA的表达模式
3 讨论
CPA是一种腈水解酶,其功能活性依赖于Glu-Lys-Cys催化三联体结构[21]。作为精氨酸途径合成腐胺过程的终反应酶,深入了解CPA基因的结构、功能对理解生物体内PAs代谢平衡具有重要意义。本文首次从橡胶树中克隆了一个CPA基因HbCPA,该基因ORF长906 bp,编码301个氨基酸,含有其功能活性所需的Glu-Lys-Cys催化三联体结构。生物信息学分析表明,HbCPA为亲水性蛋白,不含跨膜结构域和信号肽,是一个胞质酶。序列分析表明HbCPA与其他植物的CPAs一致性很高,说明CPAs在不同植物中具有相似的结构和功能。进化分析表明HbCPA与同属大戟科的木薯MeCPA、蓖麻RcCPA和麻风树JcCPA亲缘关系最近,与植物学传统分类一致。
天然橡胶是在树皮特化细胞——乳管中合成。组织表达谱分析表明,HbCPA在树皮中的表达量最高,同时该基因在死皮树树皮中明显上调表达。死皮是橡胶树割面部分或全部不排胶的现象,是橡胶树产胶、排胶障碍的极端形式。HbCPA的表达分析表明,该基因可能与橡胶树橡胶生物合成和代谢相关,可能是橡胶树产胶排胶的负向调控因子。橡胶树叶片发育分为古铜期、变色期、淡绿期、稳定期和衰老期五个时期。质体的发生、发育是橡胶树叶片由淡绿期转化到稳定期的重要原因[22]。橡胶树叶片不同发育时期的表达分析表明,HbCPA在叶片淡绿期大量表达,推测其可能与叶片由淡绿期转化到稳定期密切相关,可能与质体的发生、发育有关。
乙烯作为延长排胶时间提高产胶量而被广泛应用的植物激素,而多胺和乙烯的合成竞争同一底物S-腺苷蛋氨酸(SAM)[23]。MeJA是橡胶树乳管分化和发育的重要调节因子,可以诱导橡胶树乳管分化,调节橡胶生物合成[24-26]。本研究发现ET和MeJA处理均抑制了HbCPA的表达,但随着处理时间的延长HbCPA表达整体呈上升趋势,暗示HbCPA可能在调控橡胶树产排胶信号转导中发挥重要作用。CPAs还与植物逆境响应密切相关。水稻含有4个CPAs基因,分别为OsCPA1-4。干旱胁迫下OsCPA3表达下调而OsCPA1、OsCPA2、OsCPA4转录水平未出现变化[27]。拟南芥CPA在水胁迫处理下出现滞后上调表达[28];同样的,在本研究中发现HbCPA表达也受干旱、低温、高盐和H2O2等逆境胁迫影响,表明HbCPA可能参与橡胶树逆境胁迫应答。
4 结论
从橡胶树中克隆获得HbCPA,其开放阅读框906 bp,编码301个氨基酸,预测其为亲水性蛋白。HbCPA表达无组织特异性,在检测的组织中均有表达。HbCPA可能参与橡胶树叶片由淡绿期向稳定期转化过程,参与低温、干旱、高盐、氧化胁迫等非生物逆境应答等过程,并可能负向调控橡胶树死皮发生。
