浙江九龙山国家级自然保护区长序榆群落的结构特征及种间联结性分析
2020-12-04杜有新王军峰杨晓君王志高
杜有新,王军峰,梁 炜,杨晓君,王志高,刘 伟,①
(1. 华东药用植物园科研管理中心,浙江 丽水 323000; 2. 丽水市白云山生态林场,浙江 丽水 323000;3. 浙江九龙山国家级自然保护区管理局,浙江 丽水 323300; 4. 浙江省林业科学研究院,浙江 杭州 310023)
物种多样性和种群结构是植物群落的基本属性,影响着群落的发展方向和演化趋势[1]。一般而言,随着植被群落的正向演替,群落的种类组成和结构逐渐趋于完善和稳定,种间关系也逐步趋向正相关关系,以达到种类间的稳定共存状态[2]。对植物群落的种类组成和种间结构进行研究,可以了解群落中的种类数量及种间的空间分布关系[3-4],有利于濒危植物的保护和利用[5]。
长序榆(UlmuselongataL. K. Fu et C. S. Ding)隶属于榆科(Ulmaceae)榆属(UlmusLinn.),为落叶乔木,属国家Ⅱ级重点保护野生植物,产于福建、江西、浙江和安徽等地,零星分布于海拔500~900 m的半阳坡阔叶林及毛竹〔Phyllostachysedulis(Carriere) J. Houzeau〕林中[6]。浙江是长序榆的主要分布区,也是其模式标本的产地,在浙江九龙山国家级自然保护区[7]分布有长序榆的最大种群。目前,对长序榆的研究主要集中在种群天然更新状况[8]、种群保护[7,9]及群落生态学[10]等方面,但对长序榆群落结构及种间联结性的研究尚不深入。
鉴于此,作者以浙江九龙山国家级自然保护区的长序榆群落为研究对象,分析该群落的种类组成和群落结构以及主要树种的种间关联特征,探讨长序榆与其他树种的相互关系,并从种间关系角度探讨群落环境及其对长序榆种群发展的影响,为长序榆种群的有效保护管理提供基础研究资料。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于浙江省遂昌县九龙山国家级自然保护区,地理坐标为东经118°49′38″~118°55′03″、北纬28°19′10″~28°24′43″。该区域属亚热带湿润季风气候,年均温约17 ℃;最冷月为1月,极端最低温-17 ℃;最热月为7月,极端最高温37 ℃;年均相对湿度83%,夏季高温多雨、冬季寒冷潮湿;年均降水量约1 855 mm[10]。土壤为沙质黄棕壤。
该区域地带性植被类型属于亚热带常绿阔叶林,长序榆主要生长在海拔500~900 m石砾密布的沟谷中。该保护区内长序榆分布面积在1 000 m2以下,主要有岩坪村(东经118°53′28″、北纬28°22′34″,海拔669 m,面积约200 m2)和秀才坑(东经118°50′43″、北纬28°22′27″,海拔715 m,面积约800 m2)2个分布地点。由于岩坪村沟谷陡峭且规模较小,因而,将规模较大的秀才坑设为调查主体,该长序榆种群位于沟谷东坡,坡度20°,林分郁闭度0.85。
1.2 研究方法
1.2.1 样地设置及调查和测量方法 以长序榆集中分布的秀才坑为调查区域,在2019年6月对长序榆实际分布范围进行踏查。在该种的集中分布区内设置1块面积20 m×30 m的样地,然后将该样地划分为6个面积10 m×10 m的乔木样方;另在该种的零星分布地段增设4个面积10 m×10 m的乔木样方[6];此外,分别在每个样方的左上角设置1个面积4 m×5 m的灌木样方,在每个灌木样方的左上角设置1个面积2 m×2 m的草本样方。
对乔木样方内胸径大于等于2 cm(或株高大于等于3 m)的木本植物进行每木检测,并记录种类、胸径、株高和地理坐标等信息;记录每个灌木样方内灌木植物(包括株高小于3 m的乔木幼树)的种类、地径、株高和冠幅等信息;记录每个草本样方内草本植物的种类、株(丛)数、株高、冠幅和盖度等信息。用钢卷尺(精度0.1 cm)测量乔木胸径;用测高杆(精度0.1 m)测量木本植物株高,用钢卷尺测量草本植物株高;用钢卷尺测量植物的东西向和南北向的冠径,二者的乘积即为冠幅,以冠幅占样方面积的百分比计为盖度;采用游标卡尺(精度0.1 mm)测量灌木的地径。所有指标均重复测量2次,结果取平均值。
1.2.2 种类多样性和群落结构分析 根据调查数据分别计算乔木层、灌木层和草本层的种类数(R)和重要值(IV)[11]。根据各种类的重要值计算Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(J)[12-13]。
用大小级结构替代年龄结构的方法[3],根据株高(H)把林冠层〔包括乔木层和灌木层中株高大于等于2 m的所有乔木和灌木树种〕划分为第1层(H≥12 m)、第2层(12 m>H≥10 m)、第3层(10 m>H≥8 m)、第4层(8 m>H≥6 m)、第5层(6 m>H≥4 m)和第6层(4 m>H≥2 m),用株高级替代龄级分析长序榆种群的年龄结构和群落垂直结构[14]。用大小级指数(SDI)定量估计种群更新的连续性[15]。将胸径小于2 cm或株高小于3 m的乔木个体视为更新苗,分别统计样方内各乔木树种的更新苗数量。
1.2.3 种间总体相关性和联结性检验 基于样地调查数据,选取重要值大于等于1%的25种主要树种,将野外调查数据转化为二元数据矩阵,建立2×2联列表,并统计各个种对间的a、b、c和d值[5,16];据此计算共同出现百分率和联结系数,并进行χ2检验。
采用方差比率(VR)法检验该群落内种间的总体关联程度。在独立性假设条件下,VR期望值为1,若VR>1,表示种间呈正关联;若VR<1,则表示种间呈负关联[2]。考虑到种间的正负关联可相互抵消,故采用统计量W检验VR值偏离1的显著程度:若种间不显著关联,则W值在3.940~18.307之间的概率为90%[16-17]。采用Yates校正公式计算χ2统计量[18],检验各种对关联的显著性:若χ2≥3.841,则种间为显著性关联;若χ2<3.841,则种间为不显著性关联[2]。
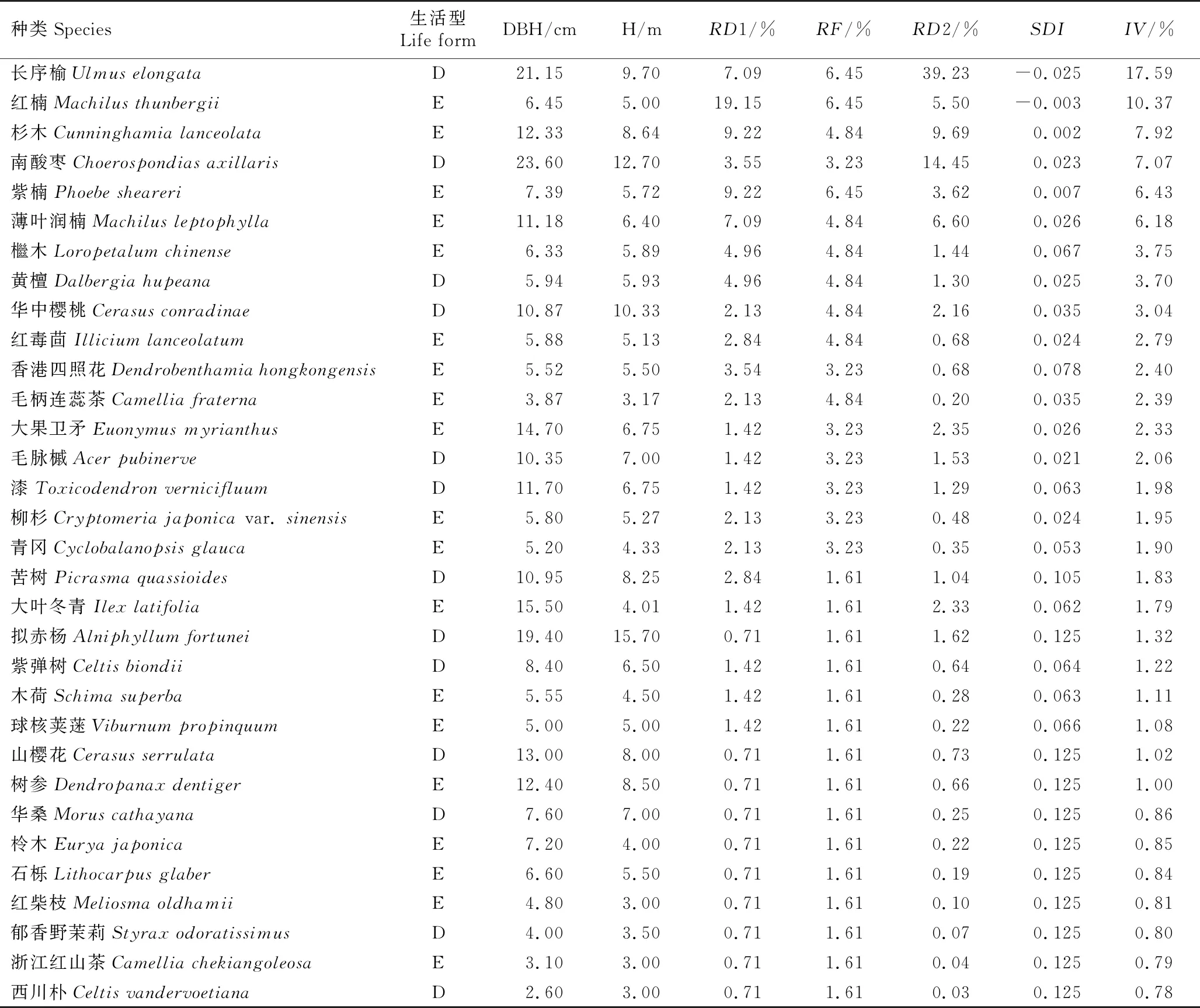
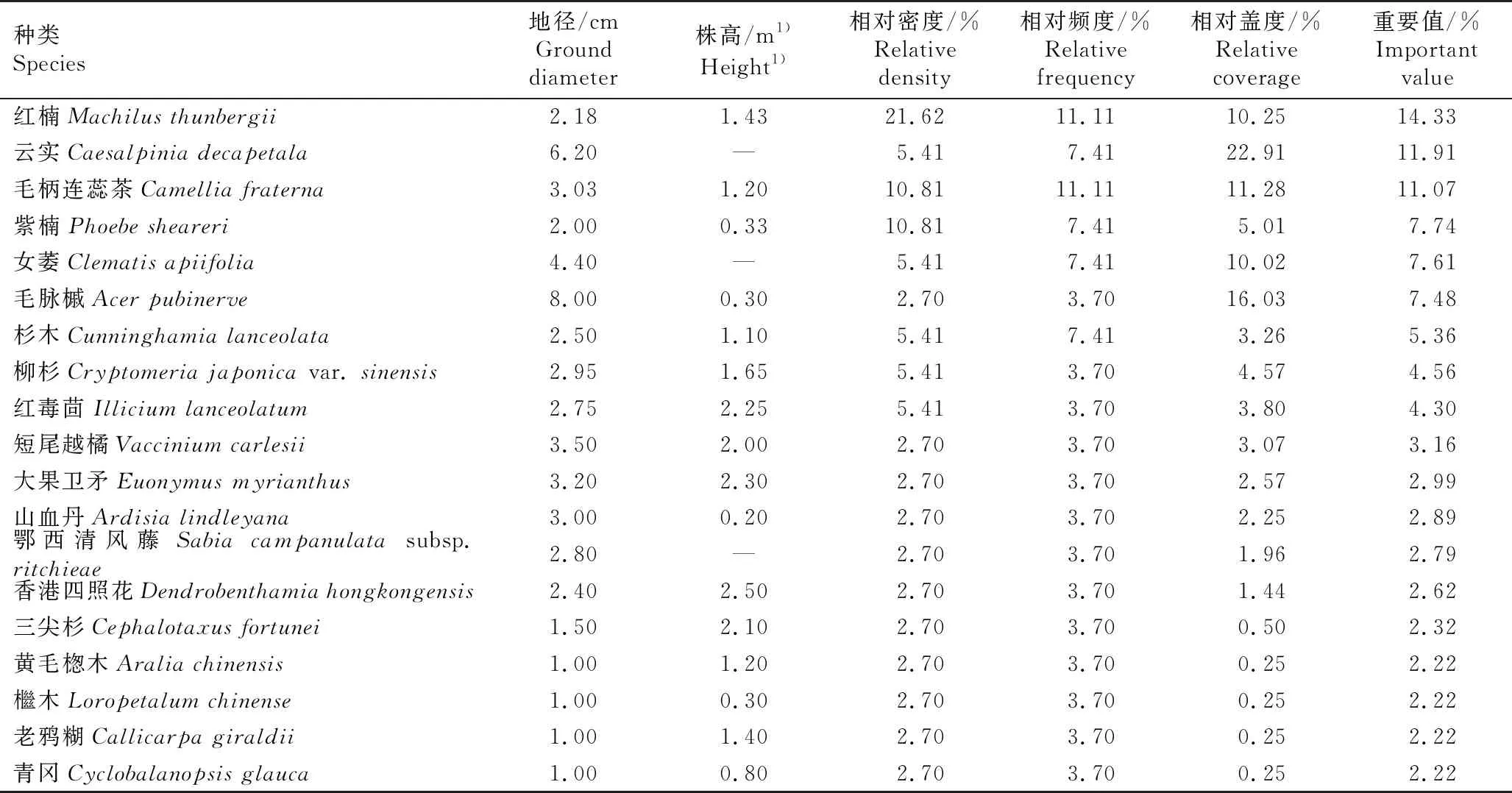
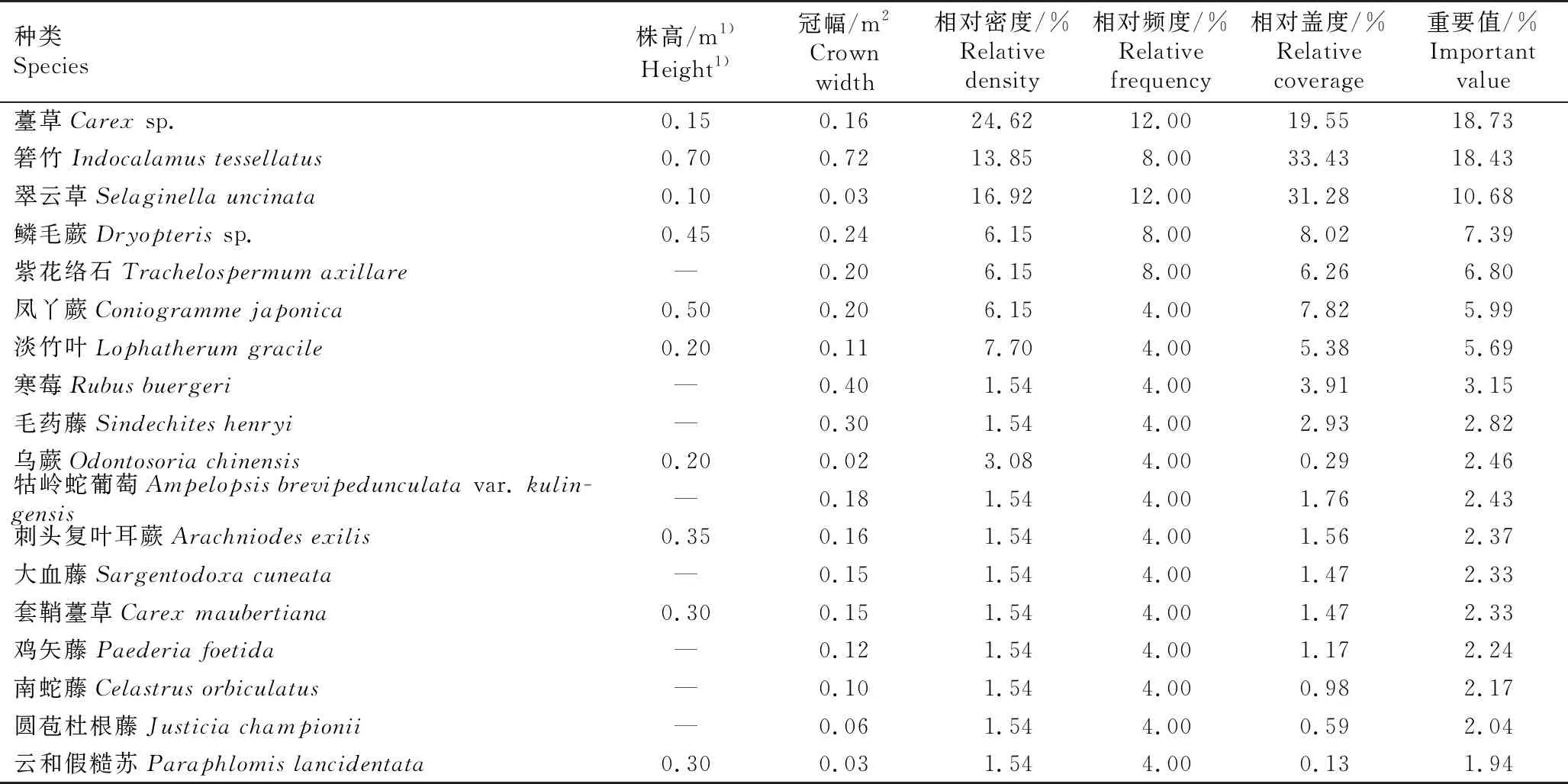
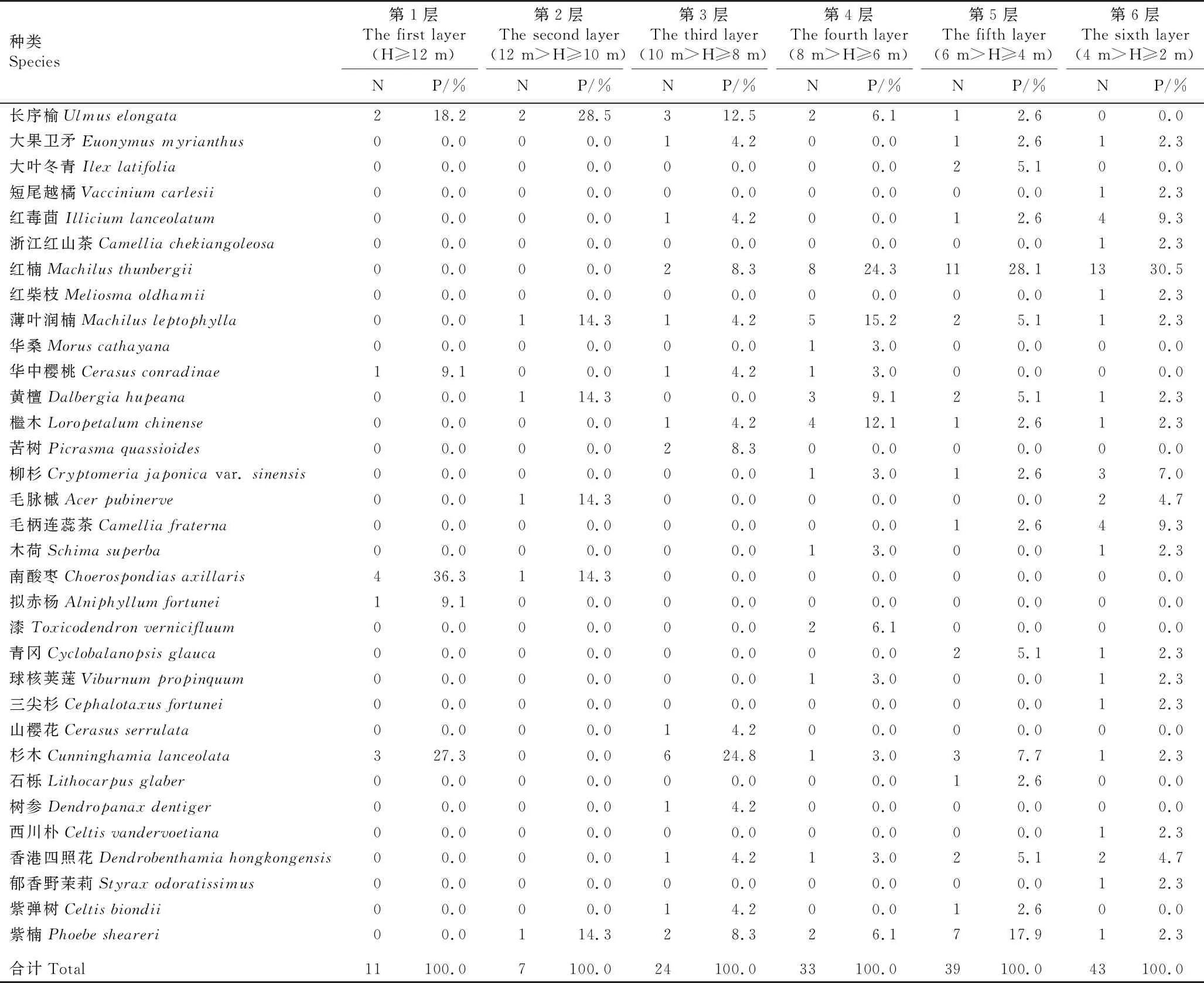
综合共同出现百分率(PC)和联结系数(AC)进一步定量分析种间联结程度[2],重点分析长序榆与其他树种的关联特征。PC的阈值为[0,1],PC值越趋近1,表明种间关联越紧密,其中,PC≥0.6为紧密关联,0.2≤PC<0.6为一般关联,0.0 采用EXCEL 2003和SPSS 17.0软件对数据进行处理和分析。 调查结果显示:浙江九龙山国家级自然保护区长序榆群落共有维管植物37科53属58种,其中乔木、灌木、草本和藤本植物分别有30、8、10和10种。根据重要值从大到小进行排序,该长序榆群落乔木层、灌木层和草本层的种类组成和重要值等基本特征分别见表1、表2和表3。 表1 浙江九龙山国家级自然保护区长序榆群落乔木层种类组成及其重要值1) 表2 浙江九龙山国家级自然保护区长序榆群落灌木层种类组成及其重要值 表3 浙江九龙山国家级自然保护区长序榆群落草本层种类组成及其重要值 由表1可见:该群落的乔木层植物有19科26属32种,其中重要值在5.00%以上的有6种,分别为长序榆、红楠(MachilusthunbergiiSieb. et Zucc.)、杉木〔Cunninghamialanceolata(Lamb.) Hook.〕、南酸枣〔Choerospondiasaxillaris(Roxb.) B. L. Burtt et A. W. Hill〕、紫楠〔Phoebesheareri(Hemsl.) Gamble〕和薄叶润楠(MachilusleptophyllaHand.-Mazz.),重要值分别为17.59%、10.37%、7.92%、7.07%、6.43%和6.18%,长序榆优势明显。乔木层植物的Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(J)分别为3.03、0.93和0.87。 由表2可见:该群落的灌木层植物有17科18属19种,包括3种藤本植物和10种乔木树种的幼树,其中重要值在5.00%以上的有7种。灌木层优势种为红楠、云实〔Caesalpiniadecapetala(Roth) Alston〕和毛柄连蕊茶(CamelliafraternaHance),重要值分别为14.33%、11.91%和11.07%;紫楠、毛脉槭(AcerpubinerveRehd.)和杉木3种乔木幼树在灌木层中也有明显优势,重要值分别为7.74%、7.48%和5.36%;藤本植物女萎(ClematisapiifoliaDC.) 的重要值为7.61%,在灌木层中也有一定的优势。灌木层植物的H、D和J值分别为2.73、0.92和0.93。 由表3可见:该群落的草本层植物有14科17属18种,包括7种藤本植物,其中重要值在5.00%以上的有7种。薹草(Carexsp.)、箬竹〔Indocalamustessellatus(Munro) Keng f.〕和翠云草〔Selaginellauncinate(Desv.) Spring〕在草本层中占有绝对优势,重要值分别为18.73%、18.43%和10.68%;鳞毛蕨(Dryopterissp.)、紫花络石(TrachelospermumaxillareHook. f.)、凤丫蕨〔Coniogrammejaponica(Thunb.) Diels〕和淡竹叶(LophatherumgracileBrongn.)在草本层中也有明显优势,重要值分别为7.39%、6.80%、5.99%和5.69%。草本层植物的H、D和J值分别为2.55、0.90和0.88。 2.2.1 群落垂直分布特征 浙江九龙山国家级自然保护区长序榆群落林冠层不同层次的种类组成见表4。结果显示:林冠层越低种类数越多。第1层5种11株,包括南酸枣4株、杉木3株、长序榆2株,其他2种各1株;第2层6种7株,包括长序榆2株,其他4种各1株;第3层14种24株,包括杉木6株、长序榆3株,其他14种分别仅1或2株;第4层14种33株,包括红楠8株、薄叶润楠5株、檵木〔Loropetalumchinense(R. Br.) Oliver〕4株、黄檀(DalbergiahupeanaHance)3株,其他10种分别仅1或2株;第5层16种39株,第6层21种43株,其中,红楠优势较为明显,第6层未发现长序榆植株。 表4 浙江九龙山国家级自然保护区长序榆群落林冠层不同层次的种类组成1) 2.2.2 群落更新状况 对群落中乔木种类的更新苗数量进行统计,结果显示:该样地中乔木树种更新苗共有10种23株,包括红楠8株,紫楠4株,杉木、柳杉(Cryptomeriajaponicavar.sinensisMiquel)和红毒茴(IlliciumlanceolatumA. C. Smith)各2株,毛脉槭(AcerpubinerveRehd.)、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、香港四照花〔Dendrobenthamiahongkongensis(Hemsl.) Hutch.〕、大果卫矛(EuonymusmyrianthusHemsl.)和三尖杉(CephalotaxusfortuneiHook.)各1株。未见长序榆的更新苗。 浙江九龙山国家级自然保护区长序榆群落主要树种间的共同出现百分率(PC)和联结系数(AC)的半矩阵图见图1。 : PC≥0.6; : 0.4≤PC<0.6; : 0.2≤PC<0.4; : 0.0 统计分析结果显示:该群落25个主要树种的方差比率(VR)为1.924 7,表明这些树种间呈正关联;统计量W值为7.698 9,介于3.940~18.307之间,表明这些树种间总体呈不显著的正关联;各种对间的χ2值均小于3.841,表明这些树种间无显著关联性。 由PC值(图1)可见:在300个种对中,PC≥0.6的种对有58个,表明这些种对紧密关联;0.2≤PC<0.6的种对有174个,表明这些种对一般关联;0.0 由AC值(图1)可见:在300个种对中,有188个种对呈正关联、112个种对呈负关联,正负关联种对的比例为1.68∶1.00。其中,AC≥0.60的种对有92个,表明这些种对紧密正关联;AC≤-0.60的种对有91个,表明这些种对紧密负关联;0.20≤AC<0.60的种对有31个,表明这些种对一般正关联;-0.20 物种多样性可反映自然生境中物种丰富度及分布均匀性状况[19]。随着植物群落演替的正向发展,资源也向最佳分配和充分利用方向发展[20],使群落物种多样性不断丰富,种间关系逐步由独立或排斥演替到共存的正相关关系[2]。在浙江九龙山国家级自然保护区内分布的长序榆群落处于常绿落叶阔叶混交林中,位于海拔500~900 m的沟谷中,群落层次较为丰富,乔木层种类的Shannon-Wiener多样性指数(H)达到3.03,明显高于刘鹏等[10]在2007年的研究结果(H=2.71)。该长序榆群落兼有常绿阔叶树和落叶阔叶树成分[21],从物种组成及优势树种看,属于常绿阔叶林到落叶阔叶林过渡阶段的次生阔叶林[22],其物种丰富度(58种)和Shannon-Wiener多样性指数均低于该保护区天然常绿阔叶林的物种丰富度(67种)和Shannon-Wiener多样性指数(H=3.29)[22]。种间联结性结果表明:该长序榆群落乔木层的主要种类总体表现为正关联,但种间关系未达显著关联水平,表明这些树种间总体呈不显著正关联关系,该长序榆群落仍处于不断演化的发展阶段。 χ2检验结果显示:该长序榆群落乔木层主要树种的种间关联不显著,说明这些树种间尚未达到稳定共存的格局[2]。这些树种间的共同出现百分率(PC)和联结系数(AC)的检验结果基本一致,如长序榆与大果卫矛、红毒茴、红楠、薄叶润楠和紫楠等10个树种的PC值均较高(PC≥0.6),种间呈紧密关联;长序榆与红楠和紫楠的AC值较高(AC≥0.60),种间呈紧密正关联,与大果卫矛、红毒茴、薄叶润楠、华中樱桃、黄檀、檵木和柳杉也呈正关联(0.20≤AC<0.60)。处于同一林层的树种呈正关联关系,如处于林冠上层的长序榆与华中樱桃和薄叶润楠等,可能是由于这些种类在资源利用和生境需求方面的相似性或互补性[15];而处于不同林层的树种在生态位上出现重叠,种间关系表现为正关联,如长序榆与次林层的红毒茴、红楠和檵木等,说明这些种类对光照和水分等环境因子具有协调利用性和资源利用的相似性[18]。此外,长序榆与南酸枣和香港四照花紧密负关联(AC≤-0.60),说明长序榆与同林层的南酸枣和次林层较耐阴的常绿树种香港四照花具有排斥性,这可能是由于长序榆与南酸枣和香港四照花对生境资源的需求存在较强的竞争关系[23-24]。由于长序榆种群自然分布区较小,供试样本数量也有限,因而,通过本研究只能解释该长序榆群落现状和种间关系的表面现象,难以揭示群落内种间关系的内在生态过程[25]。 在浙江九龙山国家级自然保护区长序榆群落中,长序榆为群落的主要优势树种,其种群结构影响着群落性质及其发展变化,但现有的长序榆种群规模小且缺乏小龄级植株,株高小于8 m的仅3株,且缺失株高4 m以下的幼苗和幼树,说明该长序榆种群的天然更新能力较差[10]。长序榆天然更新苗缺乏的主要原因除了生殖生物学上的限制外[7],还与不利的生境条件有关:一是长序榆种群的生境为石砾较多的沟谷,土层薄,影响种子萌发和幼苗生长[10];二是该群落郁闭度较高,林下光照弱[26],而长序榆为阳性落叶树种[8],林下光照条件无法满足其幼树生长所需[27]。值得注意的是,虽然在该长序榆群落中未发现长序榆更新苗,但从大小级指数(SDI)看,整个群落中只有长序榆和红楠的SDI值为负值,说明长序榆种群年龄结构较合理,种群中尚有一定数量的中小龄级个体,群落中长序榆的优势地位短期内不会有明显变化。 长序榆为珍稀濒危植物,其地理分布区相对狭窄,且居群之间相互隔离,遗传基础狭窄[28],居群间基因流缺乏导致其种群存在近交衰退现象[29]。浙江九龙山国家级保护区内的长序榆种群不仅规模小,且随群落的不断发展,该长序榆种群规模可因群落环境的变化以及自身生殖特性的限制而逐渐缩小。为遏制长序榆种群衰退,保护其野生资源,需改善长序榆的天然更新环境,适度清除有负面影响的树种,促进长序榆个体的生长发育,维持其种群的可持续发展。1.3 数据处理和分析
2 结果和分析
2.1 长序榆群落种类组成及多样性水平分析
2.2 长序榆群落结构及更新状况
2.3 种间联结性分析

3 讨论和结论
