铅胁迫对向日葵幼苗矿质元素吸收及蛋白二级结构的影响
2020-12-04牟美睿王聿双刘海学杨仁杰
牟美睿,王聿双,刘海学,杨仁杰
(1.天津农学院 农学与资源环境学院,天津 300384;2.天津农学院 工程技术学院,天津 300384)
向日葵富含蛋白质及矿质营养元素,是世界重要油料作物之一[1],具有较强的低温、干旱耐受性[2]及自我修复能力[3]。在中国西北地区随着铅锌矿产的开采,周边环境中重金属污染也成为亟待解决的问题[4]。重金属铅作为作物生长过程中的一种非必须元素[5],不仅会影响作物的生长发育[6],还会引起生理生化[7]和分子水平[8]的变化,对作物产生毒害作用[9],最终通过食物链进入人体危害健康。
矿质元素与蛋白质广泛参与植物体的各项生命活动[10],也是决定作物品质的重要指标[11]。除受遗传基因的调控,生长环境条件也影响作物吸收矿质元素[12]和蛋白质结构[13],其中重金属对作物的毒害作用在生理生化方面表现为增加氨基酸浓度[14]对蛋白质产生毒害作用[15],引起膜脂过氧化导致小分子物质外泄[16],在分子水平上能直接与蛋白质结构中的巯基结合,导致分子构象改变[17]、酶变性失活[18]及必需元素缺乏[19-20]。
目前,重金属对作物的影响已有研究,但对于矿质元素吸收及蛋白结构变化的综合研究尚未见报道,因此,本试验以3个向日葵品种幼苗(‘2603’‘HA89’和‘HA300’)为材料,运用光焰原子吸收光谱法(FASS)与傅里叶红外光谱技术(FTIR),对铅胁迫后吸收矿质元素的变化和蛋白二级结构的稳定性进行研究,分析向日葵吸收矿质元素及蛋白二级结构响应铅胁迫的相关性,为进一步研究向日葵抗重金属胁迫生理变化及蛋白分子结构变化提供参考,同时可为后续探明向日葵抗重金属胁迫机理提供理论依据。
1 材料与方法
1.1 试验材料
供试向日葵(HelianthusannuusL.)种子为稳定性良好、具有较强耐受性及适应性的自交系品种‘2603’‘HA89’和‘HA300’。
1.2 材料处理
挑选饱满均匀的籽粒于纯水中催芽8 h,播种在20 cm×20 cm的发芽盒中,每盒40粒种子,采用Hoagland营养液水培,31 d后采取样品,期间生长温度为(25±1)℃,相对湿度70%,光照强度100%,每24 h补充醋酸铅溶液40 mL,光照 8 h。本试验设5组醋酸铅处理,质量浓度分别为0 (CK)、200 mg·L-1、400 mg·L-1、600 mg·L-1、800 mg·L-1,重复3次。
1.3 测定方法
向日葵幼苗矿质元素含量测定采用火焰原子吸收分光光度计法,参照张薇等[21]所用的硝酸-高氯酸法并加以改进。称取1.0 g研磨后的向日葵样品置于消解管中,加入25 mL硝酸-高氯酸(体积比4∶1)置于电炉上加热消解赶酸,直到样品近干时取下冷却,后用体积分数为2%的盐酸定容到50 mL容量瓶内,同时做纯水空白。蛋白二级结构测定采用傅里叶红外光谱法测定,将铅胁迫后的3个品种向日葵幼苗,置于烘箱中 (60 ℃, 10 h),烘干后与溴化钾按质量比1∶200混合研磨至粉末,压片进行红外光谱采集。采用FTIR200傅里叶红外光谱仪衰减全反射法FTIR-ATR室温下扫描,扫描范围为4 000~400 cm-1,分辨率为2 cm-1,扫描累加次数为32次。
1.4 数据处理
使用Microsoft Office Excel 2016与Origin Pro 9.0软件对金属元素的数据进行处理与分析。使用Omnic 8.1软件对蛋白二级结构酰胺Ⅰ带(1 700~1 600 cm-1)进行自解卷积且残差(R2)均在0.97以上。
2 结果与分析
2.1 向日葵幼苗吸收Mg、Cu、Ca、Zn、Fe、K对铅胁迫的响应
对向日葵3种幼苗体内的矿质元素含量进行测定(图1),结果显示品种‘2603’对Ca、Mg、Zn、Fe的吸收在低浓度胁迫时显著增加,当铅胁迫浓度大于400 mg·L-1后吸收量随着浓度的增加逐渐减少,而Cu、K元素则在胁迫下吸收变为抑制,品种‘HA89’除Zn元素含量在600 mg·L-1铅胁迫下较对照组显著增加(P<0.05),其余元素均随着铅胁迫浓度的增加吸收量在0.05水平上呈显著性减小,而品种‘HA300’经铅胁迫后,促进对Ca元素的吸收,抑制对K元素的吸收,对Mg元素的吸收表现为低浓度促进高浓度抑制,对Cu元素的吸收表现为低浓度促进高浓度抑制,抑制Zn元素的吸收,对Fe元素的吸收除400 mg·L-1的胁迫均起到促进作用。由此可见,铅胁迫对向日葵幼苗体内吸收矿质元素的影响存在品种间及元素间差异,品种‘2603’表现为对大部分矿质元素低浓度促进,‘HA89’表现为对全部矿质元素抑制吸收,而品种‘HA300’则没有明显规律。
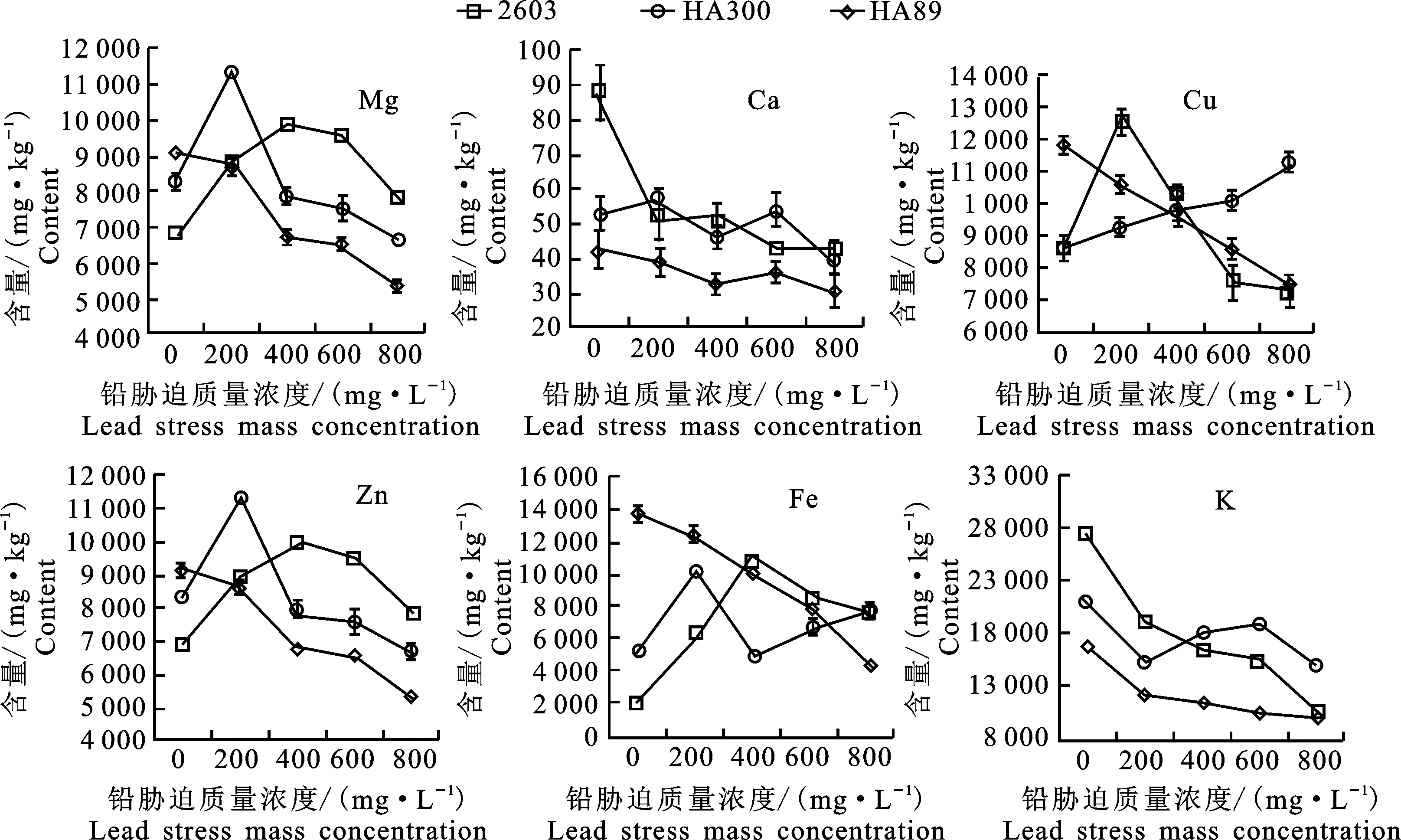
图1 不同质量浓度处理下的3种向日葵幼苗吸收矿质元素含量Fig.1 Absorption of mineral elements contents of three sunflower seedlings under treatments of different mass concentrations
2.2 向日葵幼苗蛋白二级结构对铅胁迫的响应
根据铅胁迫下3个品种向日葵幼苗蛋白二级结构的相对含量可知(图2),0~800 mg·L-1铅胁迫后向日葵3个品种的幼苗蛋白二级结构均以β-折叠和β-转角结构为主,且以β-折叠结构的相对含量最多。与CK相比,品种‘2603’幼苗在铅胁迫后其蛋白二级结构中有序含量减少,这可能是在铅胁迫后,破坏了蛋白二级结构中的氢键,分子间展开程度增加,使蛋白二级结构发生了变化,最终增加了无序结构的相对含量,使得其蛋白二级结构趋于松散。较CK而言,品种‘HA300’和‘HA89’在铅胁迫后幼苗蛋白二级结构中β-折叠结构的相对含量均上升,而β-转角结构的相对含量均下降。但二者的有序结构相对含量在200 mg·L-1、800 mg·L-1铅胁迫后下降,且以800 mg·L-1铅胁迫后的‘HA300’幼苗变化最明显,减少了2.23%,表明此时这两个品种幼苗蛋白二级结构较不稳定,不利于幼苗生长;而400 mg·L-1、600 mg·L-1铅胁迫后幼苗蛋白质分子间聚集程度增加,有序结构相对含量增加,且品种‘HA300’在400 mg·L-1铅胁迫后增加最多,为2.23%;‘HA89’则在600 mg·L-1铅胁迫后变化最明显,增加了4.37%,表明此时这两个品种幼苗能较好地抵抗铅毒害,使其蛋白二级结构不受影响。
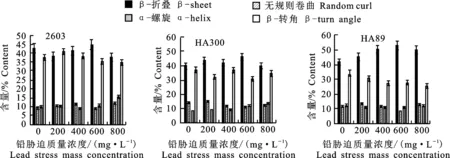
图2 铅胁迫下3个品种向日葵根蛋白二级结构含量Fig.2 Secondary structure content of sunflower root protein in three varieties under lead stress
2.3 向日葵幼苗吸收矿质元素与蛋白二级结构相关性
对向日葵矿质元素含量与蛋白二级结构中不同结构含量进行Pearson相关分析(表1),在不同浓度铅胁迫下,有序结构α-螺旋、β-折叠与大部分矿质元素呈负相关关系,其中α-螺旋在‘HA89’品种中与Ca元素相关系数为-0.9,与Fe元素相关系数为-0.93,呈显著负相关(P<0. 05),而蛋白二级结构中无序结构无规则卷曲与β-转角与除Ca、Zn元素外其余矿质元素呈不显著正相关关系,其中β-转角与Ca、Zn元素相关系数分别为0.97、0.88。由此可见,在铅胁迫下矿质元素增加的同时向日葵蛋白二级结构中的有序结构降低,蛋白质稳定性下降。

表1 3个向日葵品种在不同浓度胁迫下吸收矿质元素与蛋白结构相关系数Table 1 Correlation coefficient between mineral elements and protein structure of three sunflower varieties under stress of different concentrations
注Note:*P<0.05;**P<0.01.
3 讨 论
在植物生长过程中,重金属会导致植物体内矿质元素失衡[22]与蛋白质含量变化[23],同时植物在长期对环境的适应过程中,也会产生相应的防御机制,主要分为根部跨膜吸收[24]、木质部运输[25]、金属硫蛋白螯合作用[26]及转运蛋白结合金属离子[27]。其中,植物根部与木质部通过细胞壁中纤维素、木质素等物质与金属离子结合方式,阻止其进入植物细胞产生毒害作用,但植物根部与木质部作为植物矿质营养吸收运输的重要器官[28],在阻止重金属离子进入细胞的同时也会阻止对矿质元素的吸收,同时由于其相似的化学性质或关联的代谢途径,植物中矿质元素与重金属离子也存在相互作用和影响[12],宋洁等[7]在研究低Pb、Cd积累马铃薯品种矿质营养元素相关性时也发现土壤中高Pb能抑制大豆对Ca、Cu、Fe、Mg和Zn的吸收。因此在一定程度上矿质元素吸收的增加意味着重金属元素的积累[28],因而植物对矿质元素的吸收可以反映重金属防御机制的有效性。在本研究中,向日葵品种‘2603’对矿质元素的吸收随铅浓度的增加而增加,这与胡铁柱等[29]的研究一致,说明该品种的细胞壁并未有效结合金属离子起到防御作用;品种‘HA89’对矿质元素的吸收随铅浓度的增加表现为抑制,即植物在阻止重金属离子进入细胞的同时也阻止了对矿质元素的运输;品种‘HA300’对矿质元素的吸收随铅胁迫浓度的增加无规律性变化,这与包曙光等[30]的研究一致,这可能是由于该品种对重金属胁迫的响应存在离子间差异。
而植物对重金属的防御机制中的螯合作用与转运蛋白则是通过金属硫蛋白(MT) 、HMAs转运蛋白等蛋白质与重金属离子结合贮存在液泡中排出体外,从而缓解重金属对植物的毒害作用,同时植物体内的蛋白质可以发挥不同的作用是由于其功能特性[31],蛋白质的功能性又取决于结构的稳定性,因此蛋白二级结构可有效反应植物对重金属胁迫的抵御能力[32],同时龚宁等[33]发现蛋白二级结构的稳定性是作物抵御重金属毒害的重要原因,而史洲铭等[34]也在研究中发现,无序结构含量多可能是造成其重金属含量偏低的因素,在本研究中,向日葵品种‘2603’蛋白二级结构中无序结构相对含量增加,蛋白质功能性遭到破坏,从而导致部分蛋白无法发挥作用,破坏防御机制;而品种‘HA89’蛋白质中有序结构相对含量增加,说明该品种中蛋白质结构稳定性增加,抵御重金属胁迫的能力较强。
在本研究对3个品种向日葵幼苗矿质元素吸收量与蛋白二级结构相对含量间的关系进行了相关性分析得出,向日葵幼苗中对矿质元素的吸收量与蛋白二级结构中有序结构的相对含量呈不显著负相关关系,同时,矿质元素的吸收量与蛋白二级结构中无序结构相对含量呈不显著正相关,这可解释为向日葵幼苗在响应重金属胁迫时抵御机制发挥的作用具有一致性,这与的机制研究结论相似[35],但该论断是否合理仍有待进一步研究。
4 结 论
本研究在不同浓度铅胁迫下对向日葵吸收矿质元素及蛋白二级结构变化进行了研究,结果表明:向日葵抵御重金属胁迫存在品种间差异,随着铅胁迫浓度的增加,品种‘2603’对矿质元素的吸收增加,蛋白二级结构的稳定性降低,品种‘HA89’对矿质元素的吸收受到抑制,蛋白二级结构趋于稳定,品种‘HA300’对矿质元素的吸收无明显规律,同时在对向日葵幼苗吸收矿质元素与蛋白二级结构响应铅胁迫的相关性研究中发现,3种向日葵幼苗在铅胁迫下对矿质元素的吸收与蛋白二级结构的稳定性均存在负相关但不 显著。
