长江南京段入江支流底质多营养级微生物多样性研究
2020-12-03曹军陈鑫琪张文龙
曹军,陈鑫琪,张文龙
(1.江苏省环境监测中心,江苏 南京 210019;2.江苏省南京环境监测中心,江苏 南京 210013;3.河海大学,江苏 南京 210098)
河流底质的微生物是河流生态系统物质循环及水体净化的驱动力,且微生物对水环境变化较为敏感,常被作为指示生物用于监测和反映水质情况[1],例如富营养化导致的生态环境条件恶化可造成微生物多样性下降[2]。而水环境的污染也可造成底质微生物群落多样性降低和导致特定功能菌群产生相应变化[3-4]。现通过解析长江南京段入江支流底质的不同营养级微生物群落组成、α及β多样性,探讨平水期和丰水期不同支流的底质微生物群落的结构组成差异,以期为长江南京段入江支流的生态保护和环境监测提供科学依据。
1 研究方法
1.1 研究区概况
长江南京段自安徽省东部流入,横贯南京市,下接镇扬河段。岸线全长近200 km,平均水深>15 m,江面宽阔[5]。北岸上起浦口区驷马山河口,迄六合大河口,长约93 km,有驷马山河、石碛河、高旺河、七里河、京新引水河、石头河、岳子河、划子口河、滁河六合段等入江支流。南岸上起江宁区,迄栖霞大道河口,长约98 km,有牧龙河、江宁河、工农河、板桥河、秦淮新河雨花段、外秦淮河、金川河、北十里长沟东支、九乡河、七乡河、三江河、大道河等支流[6]。
1.2 采样时间
结合流域内降水特征及水文节律特点[7-8],于2019年平水期(4月)及丰水期(9月)采集底质样品。
1.3 采样点位
采样点位分别为:X1B、X2B、X7B、X11B、X13B、X25B、X3A、X6A、X8A、X10A、X12A、X15A—X17A、X19A、X20A,X22A、X24A、X26A—X28A,见图1。
图1 采样点位
1.4 样品采集
采集的底质样品用无菌的50 mL聚丙烯管收集,所有的样品被储存在干冰盒里,并随后转移到-80℃的冰箱里保存用于DNA提取。
1.5 DNA提取、扩增及测序
使用土壤DNA快速提取试剂盒(Fast Soil DNA Kit)快速提取沉积物样品中的DNA。通过1% 琼脂凝胶进行电泳检测提取的DNA是否质量合格。将提取的合格的DNA模板送至上海凌恩生物科技有限公司进行聚合酶链式反应(PCR)扩增,PCR仪为ABI GeneAmp 9700型,扩增的具体条件见表1。扩增子荧光定量均一化后在Illumina HiSeq 4000平台上进行配对测序。

表1 PCR扩增引物及反应条件
1.6 生物信息分析流程
测序得到的原始数据存在一定比例的干扰数据,为了提升分析序列的质量,分析前要先对测序产生的原始序列进行过滤处理,得到优化序列。
(1)序列比对和操作分类单元(OTU)分析:用Flash软件 (v1.2.7) 对>75%扩增子长度的Illumina双端原始读数进行条形码和引物裁剪,并进行组合。然后使用QIIME v.1.9.1将测得的序列按照序列相似性为97%的阈值进行OTU聚类;
(2)物种分类:使用PyNAST比对算法与 Silva v128 参考数据库比较获得细菌分类数据,与Silva v132参考数据库比较获得原生和后生动物分类数据;
(3)微生物群落多样性分析:使用PAST3软件,分析每个分类单元的相对丰度分布,从而计算样品中的微生物多样性指数。
2 结果与分析
2.1 底质细菌群落组成
21条主要入江支流42个底质样品的优化序列数为61 696,可归属于62个门,选取平均相对丰度前10位的菌门作为优势门。平水期入江支流底质样品中的优势门包括:变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidetes)等(图2a)。其中,变形菌门(Proteobacteria)平均相对丰度最高,占51.56%,绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidetes)各占9.99%、9.19%。丰水期底质样品中的优势门包括:变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)等(图2b)。其中变形菌门(Proteobacteria)平均相对丰度最高,占51.98%,酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、硝化螺旋菌门(Nitrospirae)各占8.67%、8.33%、5.01%。丰水期与平水期相比,底质的细菌优势门在种类上基本一致;数量上,酸杆菌门(Acidobacteria)和硝化螺旋菌门(Nitrospirae)的相对丰度分别上升1.29%和2.23%,拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)和厚壁菌门(Phylum Firmicutes)各下降3.08%、1.66%和1.15%。
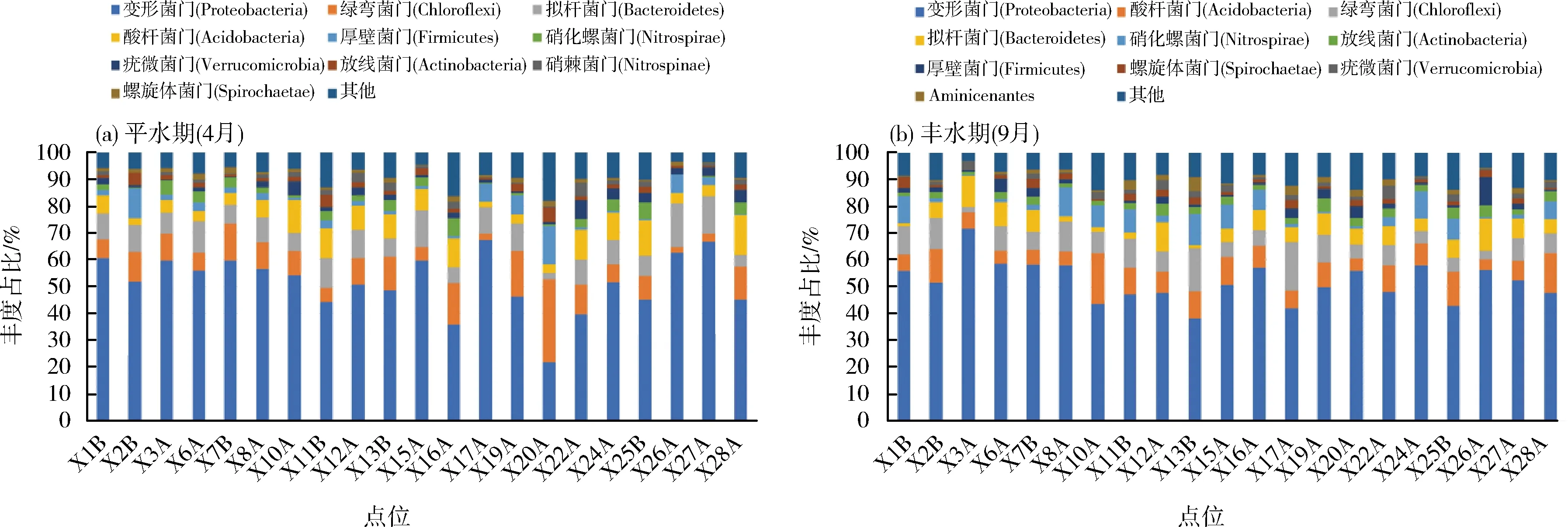
图2 入江支流底质样品在门水平上的细菌群落组成
样品优化序列可归属于285个纲,选取平均相对丰度前20的菌纲作为优势纲。平水期入江支流底质样品的优势纲包括:β-变形菌纲(Betaproteobacteria)、γ-变形菌纲(Gammaproteobacteria)、δ-变形菌纲(Deltaproteobacteria)、厌氧绳菌纲(Anaerolineae)、α-变形菌纲(Alphaproteobacteria)等(图3a)。其中,β-变形菌纲(Betaproteobacteria)的平均丰度最高,达到19.08%;γ-变形菌纲(Gammaproteobacteria)次之,平均丰度占14.35%;δ-变形菌纲(Deltaproteobacteria)、厌氧绳菌纲(Anaerolineae)、α-变形菌纲(Alphaproteobacteria)各占12.13%、5.81%、4.52%,梭菌纲(Clostridia)、鞘脂杆菌纲(Sphingobacteria)、硝化螺旋菌纲(Nitrospirae)各占3.01%、2.99%、2.78%。丰水期底质样品的优势纲在组成上与平水期一致(图3b)。而在数量上存在明显的水期差异:平水期相对丰度最高的β-变形菌纲(Betaproteobacteria)的优势地位在丰水期被δ-变形菌纲(Deltaproteobacteria)所取代,β-变形菌纲(Betaproteobacteria)相对丰度下降3.09%;厌氧绳菌纲(Anaerolineae)相对丰度下降2.50%;硝化螺旋菌纲(Nitrospirae)相对丰度上升2.23%。
β-变形菌纲(Betaproteobacteria)包含了固氮细菌,为沉积物提供了更强的固氮能力,是富营养化沉积物的主导微生物,适合生活在较高富营养化的水环境中[9]。硝化螺旋菌纲(Nitrospirae)具有氨氧化和硝化作用,促进氮循环[10-11];厌氧绳菌纲(Anaerolineae)属于兼性厌氧细菌(anaerobic bacteria),也具有反硝化功能[12]。因此结合本研究中不同水期各优势菌纲的数量变化可见,丰水期入江支流底质环境的硝化作用可能得到加强,而固氮和反硝化作用有所减弱;且相比平水期的底质环境溶氧较高,水体富营养化程度较低。王毛兰[13]和魏静等[14]的相关研究分别表明丰水期水体的富营养化水平和氨氮浓度相较平水期和枯水期较低,与文章分析结果一致。
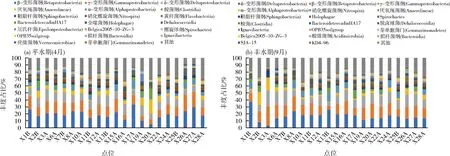
图3 入江支流底质样品在纲水平上的细菌群落组成
2.2 细菌群落多样性
Shannon指数综合考虑群落的丰富度和均匀度,其数值越大,表明群落结构的复杂程度和稳定性越高,生物多样性越高[15-16]。而生物多样性是河流生态系统平衡和河流系统健康的基础[17],常被用作监测和预测一个水生生态系统健康与否的标准[18]。HU等[19]曾比较多种生物指数和多样性指数对明珠湖水质评价的结果,提出采用Shannon指数能较准确对该湖的水质进行评价。现采用Shannon指数表征入江支流底质微生物群落的复杂程度和稳定性,以及河流生态系统健康的程度。见表2。

表2 入江支流底质样品的细菌、原生动物和后生动物的Shannon指数
由表2可见,平水期细菌Shannon指数数值为5.803~7.608,丰水期Shannon指数数值为5.767~7.488,均值分别为6.828和6.899。Shannon指数明显较低的点位有X3A(北十里长沟东支,丰水期)、X20A(石头河,平水期)、X27A(板桥河,平水期)、X17A(七里河,平水期)等。同时基于Bray-Curtis距离的非度量多维尺度分析(NMDS,见图4)表明,平水期和丰水期的样品群落组成呈现比较明显的差异;平水期X2B(金川河)、X20A(石头河)与丰水期X13B(高旺河)的底质样品细菌群落的结构组成与同水期的其他点位差异较大。NMDS结合置换多元方差分析(ADONIS,p= 0.047 < 0.05)表明入江支流底质细菌群落组成在平水期与丰水期具有差异。底质样品的理化性质检测分析结果显示硝酸盐氮、总磷(TP)和总有机碳(TOC)在平水期和丰水期存在显著差异(独立样本t检验,p< 0.05),而平水期金川河、石头河的底质TOC含量远高于其他点位,表明入江支流底质环境在不同水期存在差异。

图4 不同水期各采样点底质样品的非度量多维尺度分析(NMDS)
2.3 底质原生动物和后生动物群落组成
选取平均相对丰度较高的原生动物及后生动物作为优势种群,基于Bray-Curtis距离算法对样本距离矩阵进行聚类分析,构建样本层级聚类树与群落结构堆积图组合形式展现,见图5(a)(b)。原生动物中,不等鞭毛类(Stramenopiles)、囊泡虫类(Alveolata)和绿色植物(Chloroplastida)为主要优势类群,相对丰度分别为42.31%,27.96%和16.86%。后生动物中,节肢动物(Arthropoda)、轮虫(Rotifera)和线虫(Nematoda)为主要的优势类群,相对丰度分别占27.19%,17.56%和14.16%。聚类分析表明,平水期X2B(金川河)、X16A(划子口河)、X28A(秦淮新河)和丰水期X2B(金川河)、X8A(三江河)、X11B(驷马山河)与同水期其他点位的相似性很低,相对独立。说明这些河流底质的原生及后生动物群落构成与其他河流差异较大。

图5 入江支流底质样品的原生动物及后生动物在的群落组成
2.4 原生动物和后生动物多样性
底质样品中原生动物和后生动物的Shannon指数结果见表2。21个采样点中14个采样点的样品原生动物多样性在丰水期高于平水期,Shannon指数数值为0.640~5.308,平水期和丰水期的均值分别为4.046和4.178;平水期和丰水期中分别有X2B(金川河)和X6A(九乡河)、X17A(七里河)的原生动物多样性显著低于其他点位,金川河采样点尤其显著。另一方面,12个采样点的后生动物多样性丰水期高于平水期;Shannon指数数值为0.369~2.984,平水期和丰水期的均值分别为2.013和1.906;平水期和丰水期中分别有X17A(七里河)、X11B(驷马山河)和X25B(牧龙河)的后生动物多样性明显低于其他点位。
原生及后生动物的多样性往往与污染物浓度呈负相关[20],即水环境的污染程度越高,原生动物和后生动物的多样性越低。由表2可见,整体上平水期的原生及后生动物的多样性较低,表明平水期入江支流的底质生态系统稳定性相对较差、水体污染程度整体上可能稍严重于丰水期;而金川河、七里河、牧龙河等河流的水污染可能尤为严重。金川河作为南京市重要入江支流且包含省控断面,其流域主要覆盖老城区,人口密集[21],长期存在水质不达标、水环境恶化等问题。虽然近年来相关部门对入江支流进行了水环境整治,但以上分析表明金川河、七里河、石头河等入江支流底质的多营养级微生物多样性较低,水生态系统可能不够稳定。
3 结论
(1)长江南京段入江河流底质多营养级微生物优势种与国内其他区域河流组成类似。细菌的优势菌门为变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)等,优势菌纲为β-变形菌纲(Betaproteobacteria)、α-变形菌纲(Alphaproteobacteria)、δ-变形菌纲(Deltaproteobacteria)等。原生动物和后生动物中优势类群分别为不等鞭毛类(Heterknta)、囊泡虫类(Alveolata)和节肢动物(Arthropoda)、轮虫(Rotifera)等。
(2)细菌优势纲中硝化螺旋菌纲(Nitrospirae)、厌氧绳菌纲(Anaerolineae)等在相对丰度上的水期差异显示丰水期入江支流底质环境相比平水期有较高的硝化作用、溶氧较高而反硝化作用减弱,进一步表明丰水期入江支流的富营养化水平和氨氮浓度比平水期较低。
(3)细菌、原生动物、后生动物多营养级的群落结构组成和多样性分析结果表明,入江支流不同水期的微生物组成存在显著差异,丰水期的多营养级微生物多样性整体上高于平水期。
