烃源岩中不溶有机质:源于生物体的骨壁壳有机大分子
2020-12-02秦建中潘安阳申宝剑
秦建中,潘安阳,申宝剑
(1.页岩油气富集机理与有效开发国家重点实验室,江苏 无锡 214126;2.国家能源页岩油研发中心,江苏 无锡 214126;3.中国石化油气成藏重点实验室,江苏 无锡 214126;4.中国石化 石油勘探开发研究院 无锡石油地质研究所,江苏 无锡 214126)
沉积有机质的原始来源是生物体及其分泌物、排泄物、代谢产物和生物残体[1]。这些物质稳定性差异很大,其中核酸、糖类等有效碳水化合物及活性蛋白质等稳定性较差,只有生物脂类物质(细胞的“油滴”、“脂肪粒”及细胞膜等类脂的“生物标志物”)和生物骨壁壳碎屑相对稳定,容易被保存下来。生物脂类物质是可溶有机质或地下石油最重要的来源[2],在优质烃源岩中经温度、压力、时间、催化等成岩作用而形成石油[3]。生物骨壁壳碎屑可以分为有机和无机两部分,生物骨壁壳有机碎屑就是不溶有机质,实质上是由纤维素、几丁质等无效碳水化合物或硬蛋白等非活性蛋白构成的有机大分子,也是本文讨论的对象。生物骨壁壳有机大分子与干酪根相当,均为不溶有机质,但又不完全一样,表现在沉积早期至低成熟阶段,干酪根不但包括了生物骨壁壳有机大分子,而且还包裹了(或与某些杂原子低能键相连接的)脂类小分子,是一种复杂和不均匀的缩聚物,分子结构难以用分子式表述,至今尚未弄清楚[4]。这里试图从生物分子结构和物质平衡的角度研究石油、天然气和不溶有机质,与干酪根生烃理论并不矛盾。
1 生物体骨壁壳有机大分子的构成与稳定性
对生物体起支持或保护作用的有机骨壁壳等生物高分子主要是由①纤维质+果胶质、木质素、几丁质、肽聚糖等无效碳水化合物和②硬蛋白等非活性蛋白质构成。它们的化学结构相对稳定且不易分解,不溶于水和一般有机溶剂。在还原、强还原尤其是在水体分层及底层水体呈现H2S的厌氧环境下形成的优质烃源岩中,生物体骨壁壳有机大分子尽管经历了长时间地质年代沉积成岩的演化,但其主要成分及部分结构相对容易保存下来,是不溶有机质的主要成分。
1.1 纤维素、半纤维素、木质素、几丁质、肽聚糖、果胶等无效碳水化合物构成的骨壁壳有机大分子不易分解,较为稳定
碳水化合物(糖类化合物)是自然界最丰富的有机物,根据碳水化合物的稳定性或能否被生物体消化吸收,可分为无效碳水化合物和有效碳水化合物。将生物体不能消化的碳水化合物或不能被消化成小分子进而被吸收的糖类统称为无效碳水化合物,主要包括构成生物骨壁壳的纤维素、半纤维素、木质素、几丁质、肽聚糖和果胶等多糖类或聚糖类,也包括各种结构糖蛋白类。
(1)纤维素大分子是由很多D-葡萄糖以β-1,4糖苷键组成的多糖,分子式为(C6H10O5)n,其化学组成含碳44.44%、氢6.17%、氧49.39%[5-6]。天然纤维素是极性高分子,分子之间存在氢键,在常温下呈现较稳定的化学结构,最新研究表明其是柔性分子[7]。温度在150 ℃以下时,纤维素不发生显著变化,超过该温度时即会由于脱水而不断焦化,发生分解的温度为325~375 ℃[8]。纤维素是自然界中分布最广且最丰富的有机物,地球上每年通过光合作用合成约2 000×108t纤维素,占植物界碳含量的50%以上[9]。纤维素是植物(维管束植物、地衣植物)细胞壁中的主要成分,以高度结晶的有序结构微纤丝状态存在,构成了植物细胞壁的骨架结构,与半纤维素和果胶质等构成完整的细胞壁[10-11]。一部分藻类(如绿藻、红藻、灰藻、黄—绿藻等)的细胞壁[12-13]、一些微生物(如醋酸菌)[14-15]以及被囊动物(亦称尾索动物,如海鞘)的被囊也能产生纤维素[16-17]。
半纤维素是由几种不同单糖组成的聚糖高分子物质[18-19],分解温度为225~350 ℃[8]。在植物细胞壁中半纤维素与纤维素之间通过氢键和范德瓦耳斯力结合共生,半纤维素附着在微纤丝的表面,并把相邻的微纤丝相互连接,构成了坚硬的细胞网络结构[20-21]。半纤维素在植物体木质组织(针叶木、阔叶木和禾本科等)中含量丰富,在非木材植物中(如麦秆和稻类秸秆)更是占比30%以上[22]。半纤维素化学成分因植物种属的不同而有很大差异,可分为木葡聚糖、木聚糖、β-(1→3,1→4)-葡聚糖、甘露聚糖和葡甘聚糖这几类[23]。其中木葡聚糖存在于从苔藓植物开始的每一种陆生植物中,但在轮藻中还未发现[24-26];木聚糖广泛存在于被子植物以及诸如松柏类的裸子植物中[23];甘露聚糖则是轮藻中的主要半纤维素[25-26],似乎在早期的陆生植物中就非常丰富了,尤其是苔藓植物[27]和石松门植物[28];β-(1→3,1→4)-葡聚糖存在于整个禾本目植物中,但很多在种子植物类群中并不存在[23]。
木质素从严格意义上来说并非碳水化合物,而是复杂酚类聚合物,自然界中它与碳水化合物(纤维素、半纤维素等)交联在一起,通常以木质素—碳水化合物复合体的形式存在[29]。不同的植物木质素的元素组成往往会存在一些差别[30],但总体来说木质素的化学组成主要含碳、氢、氧3种元素,分别约占60%,6%和30%,此外还含有少量的氮元素[31]。木质素也是构成高等植物细胞壁的主要成分之一。在自然界普遍存在的植物中,一般大分子物质纤维素(30%~50%)、半纤维素(23%~32%)和木质素(15%~25%)为主要成分[32],其中木质素在针叶木形成层中、阔叶木形成层中和单子叶禾木科植物茎中分别占23%~35%,20%~25%和15%~25%[19]。
(2)几丁质又称甲壳素、壳多糖等,分子式(C8H13O5N)n。几丁质广泛存在于自然界,是地球上仅次于纤维素的第二大纤维源[33],为甲壳类动物的外壳、软体动物的器官和部分真菌类(酵母菌等)细胞壁的主要成分,也存在于一些绿藻中[34]。
(3)肽聚糖又称黏肽、黏质复合物、胞壁质,为细菌细胞壁的主要成分[35]。不同种类细菌细胞壁所含肽聚糖占细胞干重的比例有所差异,革兰氏阳性细菌和革兰氏阴性细菌细胞壁所含的聚糖分别占干重的50%~80%和5%~20%,只有少数细菌(如嗜盐菌)不含肽聚糖[36]。
葡聚糖为一种由D-葡萄吡喃糖单体组成的高分子量多糖,存在于某些微生物分泌的黏液中,结构因微生物种类和生长条件的不同而有差别,另外在一些植物和真菌中也存在葡聚糖[37]。胞外聚合物(Extracellular Polymeric Substances,EPS)是在一定环境条件下主要由细菌等微生物分泌于体外的高分子聚合物,包括荚膜、黏液层以及其他表面物质[38],其中组成以多糖和蛋白质为主(占总量的70%~80%),还有少量的核酸、磷脂、褐藻酸、腐质酸等物质[39-40]。黏多糖是多糖分子与蛋白质结合而成的胞外生物高分子,是细胞间结缔组织的主要成分。
(4)果胶又称原果胶、果胶质,普遍存在于高等植物中,是构成植物细胞间层质的主要物质[41]。
藻胶是由藻类细胞壁中的多糖组成,包括卡拉胶、琼脂、褐藻胶及其衍生物。其中卡拉胶又称石花菜胶、鹿角菜胶等,是来自红藻类的细胞壁多糖,不同世代的红藻所含有的卡拉胶主要类型不同[42]。琼脂,又称琼胶,分子式(C12H18O9)n,也主要为红藻类植物细胞壁的组成成分,由琼脂糖(形成凝胶的组分)和琼脂胶(非凝胶部分)两部分组成[43]。褐藻胶是广泛存在于褐藻细胞壁中的一类多糖物质,一些细菌(如假单胞菌属、固氮菌属等)也可以产生胞外褐藻胶[44-45]。藻类的原生质体也能够分泌出一层果胶质的被膜,当细菌等生物死亡后,残体也能够被类胶质体黏接包裹,藻类死亡后则被黏接起来,甚至形成连片状的藻席。
1.2 硬蛋白等非活性蛋白多为三螺旋结构,很难被分解开,相对稳定
蛋白质是由基本组成单位氨基酸构成的有机大分子,是构成细胞的基本有机物。蛋白质的分类或种类有很多,这里根据生物体中蛋白质的活性或稳定性和功能特征分为非活性蛋白质和活性蛋白质。非活性蛋白质是一大类对生物体起保护或支持作用的结构蛋白质或硬蛋白或胶原蛋白等,很难被分解。常见的非活性蛋白有硬蛋白,它是一群不溶于水、盐、有机溶媒、稀酸和碱的简单蛋白质的总称,包括胶原蛋白、角蛋白和弹性蛋白等。
(1)胶原蛋白是动物结缔组织(骨、软骨、皮肤、腱、韧等)中的主要成分,约占胶原纤维固体物的85%。胶原蛋白不同于一般的蛋白质的双螺旋结构,而是稳定的三螺旋结构[46],普遍存在于动物体中,也是哺乳动物体内含量最多、分布最广的功能性蛋白[47-48]。其中骨胶原蛋白也称“骨胶原”(UC-Ⅱ),是由三条多肽链构成的螺旋分子结构,是胶原蛋白的主要成分,并构成了骨骼和软骨(占90%)[47]。
(2)角蛋白是保护或加强机械强度的不溶性蛋白质,在所有溶剂中都不发生溶解,也不能被酶分解。存在于毛发、指甲、羽毛、角等中[49]。
(3)此外,还有动物体中起支持与润滑作用的弹性蛋白。
2 不同类型生物体有机骨壁壳及碎屑
对生物体起支持或保护作用且相对坚固稳定的生物有机高分子物质构成了不同类型生物体的有机骨壁壳及其碎屑,其精细结构多数只有在扫描电镜、纳米CT、FIB、透射电镜等超显微技术下才能被识别。包括浮游藻类、疑源类、底栖藻类、真菌类和细菌类等的细胞壁壳及其碎屑;浮游动物结缔组织的壳骨皮毛丝腱韧及其碎屑;水生陆生高等植物的细胞壁和细胞骨架及其碎屑。
2.1 浮游藻类及疑源类细胞壁的主体成分是纤维素+果胶
浮游藻类细胞壁的主要成分是纤维素+果胶[50]。如浮游绿藻细胞壁(图1a)[51]的外层是果胶质,内层是纤维质,有些目/属的细胞壁中还有几丁质(刚毛藻属、鞘藻属和毛鞘藻属)和胼胝质(松藻目)。甲藻类(图1b)[52]除少数裸型种类外,都具有主要是纤维素组成的厚细胞壁(也称为壳),由许多具有花纹的甲片相连而成。黄藻类细胞壁化学成分主要是果胶,有些种的细胞壁内沉积有二氧化硅,只有无隔藻属和黄丝藻属的细胞壁是由纤维素组成。硅藻类细胞壁(图1c)成分含有果胶质和硅质,不含纤维素。不同于其他藻类,蓝藻细胞壁(图1d)化学组成以肽聚糖为主。隐藻细胞不具纤维素细胞壁,细胞外有一层周质体,柔软或坚固。
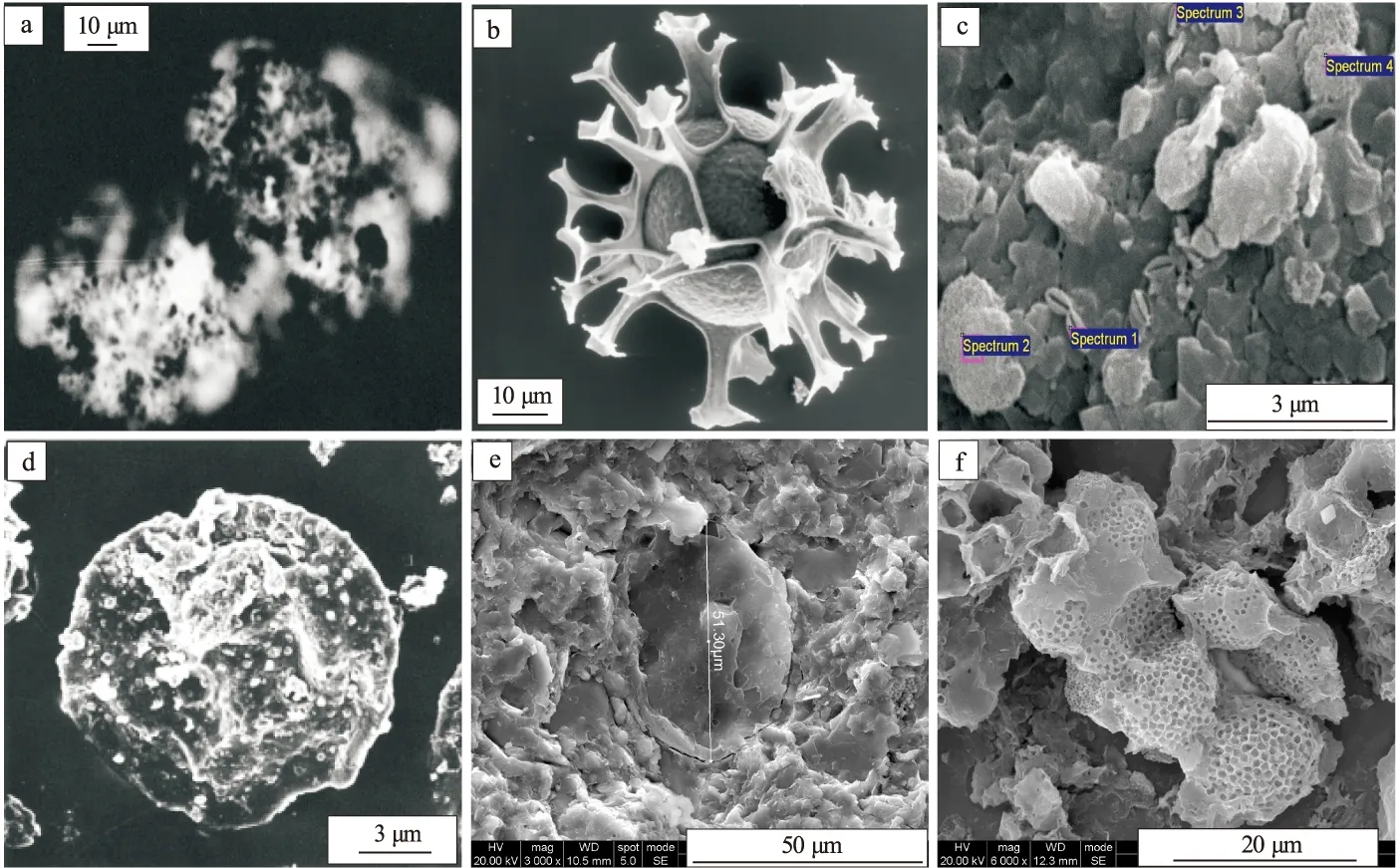
图1 浮游藻类和疑源类纤维质细胞壁及其碎屑扫描电镜照片
疑源类(图1e,f)是一类多源的、具有不同亲缘关系的微体化石类群,可能包括海生真核浮游植物、绿藻和单细胞原生生物的有机质壁囊胞,以及一些非海相的形态类型(如真菌孢型、蓝藻)和非藻类亲缘的生物形态(如高等生物的卵、外骨骼)[53]。疑源类是具有机细胞壁的,由单一或多层有机成分的壁包封的中央腔组成。疑源类的细胞壁主要成分也是纤维素,也有果胶质、几丁质及肽聚糖等无效碳水化合物。
这些源于海洋(或海侵湖泊)浮游藻类和疑源类的细胞壁壳及其碎屑结构相对稳定,在稳定水体(强还原环境)沉积过程中,常常可以与油滴类脂等脂类物质一起保存下来,但是其量相对保存下来的脂类物质来说所占比例很低,一般不超过15%[54]。这些细胞壁碎屑+油滴类脂等脂类也可以在细菌及其胞外聚合物的共同作用下,形成具有荧光的组分,但是细胞壁壳即使在未成熟—低成熟阶段也不具荧光,在高成熟—过成熟阶段通过超显微技术可识别其原始形态。实际上,正是由于这些浮游藻类和疑源类的纤维质细胞壁保存在高成熟—过成熟阶段优质烃源岩中才能被识别,而此阶段的油滴类脂等脂类物质已经裂解和缩聚为甲烷等气体和固体沥青。
2.2 褐藻等底栖藻类的细胞壁及其碎屑主体成分是纤维素+藻胶;真菌类细胞壁及其碎屑主体成分是几丁质或纤维素;细菌类细胞壁的主体成分是肽聚糖或杂多糖
(1)底栖藻类中褐藻细胞壁分为两层,内层是纤维素,外层是藻胶,同时在细胞壁内还含有褐藻糖胶(一种无效碳水化合物)。底栖藻类中红藻细胞壁也分两层,内层为纤维素,外壁是由琼胶和卡拉胶等果胶组成,因种类而异[50]。此外,底栖宏观藻——褐藻等“茎枝”丝炭化可以形成海相丝质体或半丝质体;藻类细胞分泌物形成的分泌体也可能属一种海相惰质体。褐藻等底栖藻类的纤维质+藻胶细胞壁也可以凝胶化形成镜状体[55-56]。宏观藻类细胞壁或丝壁及其碎屑尤其是在细菌及其胞外聚合物的共同作用下,可以形成藻席(图2a,b,c)。例如,优质烃源岩中常见的底栖藻类细胞壁或丝壁及其碎屑主要有①底栖红藻的丝质体残片和丝壁碎屑;②藻席和底栖藻孢子等[57]。
(2)真菌细胞壁和菌丝体主要成分为几丁质,也有纤维素、葡聚糖、甘露聚糖等多糖[58]。高等真菌、低等真菌和酵母菌分别以几丁质、纤维素和葡聚糖为主。霉菌细胞壁分为3层,外层、中层和内层分别是无定形的β葡聚糖、糖蛋白和几丁质微纤维。子囊菌亚门真菌菌丝体(亦称子囊菌,丝状子囊菌细胞壁由几丁质构成)与藻类(以绿藻或蓝藻为多)共生形成地衣。海洋(或海侵湖泊)真菌地衣类的几丁质或纤维质丝壁碎屑,与动物的几丁质外壳碎屑和底栖藻类的纤维质丝壁碎屑一样,结构相对稳定(图2d)。此外,真菌遗体形成的真菌体可以是一种海相惰质组[59]。
(3)细菌类细胞壁的成分多为肽聚糖(图2e)。细菌类分泌物形成的胞外聚合物(图2f)结构相对较稳定,在厌氧条件下,这些细菌类肽聚糖的壁壳可以单独保存下来,也可以与其他相对稳定的有机生屑一起保存下来。古细菌细胞壁不含肽聚糖,不同古细菌细胞壁成分不同,包括蛋白质、杂多糖(如甲烷八叠球菌和盐球菌)、假肽聚糖(如产甲烷菌),目前有关烃源岩中观察到古细菌的报道较少,但甘油二烷基甘油四醚类生物标志物的发现指示古细菌确实存在。
因此,褐藻等底栖藻类的细胞壁(或丝壁)及其碎屑主体成分是纤维素+藻胶;真菌及地衣类的细胞壁(或丝壁或菌丝)及其碎屑主体成分是几丁质或纤维素;细菌类(真细菌)细胞壁的主体成分是肽聚糖,古细菌类细胞壁的主体成分类似于肽聚糖或是杂多糖。这些源于海洋(或海侵湖泊)低等植物的细胞壁(或丝壁或菌丝)及其碎屑在优质烃源岩中常见。此外,这些低等植物的细胞壁也可以凝胶化形成镜状体,菌丝或丝壁可成为菌丝体。
2.3 甲壳类等节肢动物的骨壳及其碎屑主体成分是几丁质;笔石类等半索动物的壳及其碎屑主体成分是骨胶原;原生动物的外壳及其碎屑除有硅质、钙质外,还有纤维质、几丁质、硫酸锶等
(1)甲壳类及三叶虫类等节肢动物的骨壳在优质烃源岩中通常用普通显微镜甚至肉眼就可以见到(图3a),多为几丁质组成。其中甲壳纲是节肢动物门中的一个重要的纲,甲壳纲中的桡足类、枝角类分别是海洋和淡水浮游动物群落中最重要的类群[60]。三叶虫属于节肢动物门中的三叶虫纲,是所有化石动物中最丰富的,仅出现在寒武纪—二叠纪期间。

图2 优质烃源岩中底栖藻类、真菌类和细菌类丝壁壳碎屑扫描电镜照片

图3 动物类有机骨壳碎屑化石Fig.3 Bone and shell detritus fossils of animals
(2)笔石类等半索动物的骨壳(图3b)视为骨胶原(硬蛋白),一般保存为化石的是其硬壳(或皮)(图3c)。笔石作为一类现已灭绝的主要繁盛于早古生代的海生群体动物,保存下来的仅是笔石虫体所分泌的骨骼,即笔石体,普遍发育的五峰组—龙马溪组富含笔石页岩地层是我国四川盆地重要的页岩气勘探目标层系。虽然关于笔石体的成分一直存在着争议,但通过能谱分析普遍证实笔石体主要由碳元素组成[61-62],而且通过透射电子显微镜、显微傅里叶红外光谱和高分辨飞行时间二次离子质谱仪分析表明,笔石体可能主要是由胶原蛋白类高分子聚合物组成[63-66]。
(3)原生动物的外壳除有硅质、钙质外,有些鞭毛虫、肉足虫、纤毛虫、等辐骨放射虫的内外骨骼还由纤维质、几丁质、硫酸锶等组成。此外,还有软体动物的外套膜、鳃丝及最初贝可能由角蛋白、几丁质构成,脊索动物类的被囊(甲壳)由纤维素组成[60]。
因此,上述甲壳类、三叶虫等节肢动物的几丁质骨壳、笔石类等半索动物的骨胶原壳和原生动物的纤维质壳等动物有机骨壳碎屑,均可经细菌凝胶化作用形成镜状体或海相镜状体,也可以直接演化成为碳膜,或称之为海相惰质组[67]。在放射虫、海绵等原生动物的硅质骨骼碎屑中和在有孔虫等钙质动物骨壳碎屑中,多数也含有几丁质或胶原蛋白或黏多糖等(与动物有机骨壳成分相同的)用来黏结生物分子结构的有机质。这部分有机质含量和生烃能力以前未进行研究和予以重视,在中国南方海相等硅质生屑或钙质生屑优质烃源岩中通过扫描电镜+能谱分析等已经得到证实,其含量及对有机质类型的评价不可忽视[68-70]。
2.4 水生陆生高等植物的细胞壁主要成分是纤维素+果胶或纤维素+木质素+半纤维素+果胶,细胞骨架的主要成分是蛋白质纤维
高等植物(陆生和水生)的细胞壁的主要成分是纤维素+果胶或纤维素+木质素+半纤维素+果胶,而细胞骨架的主要成分是蛋白质纤维(非活性蛋白)(图4)。这些骨壁碎屑结构稳定,在形成优质烃源岩的稳定水体(强还原环境)沉积环境中可以保存下来,但是水体沉积环境不利于高等植物的发育,尤其是陆源高等植物骨壁碎屑只有靠长距离的搬运(水体或风)才能到达。因此,优质烃源岩中的陆源高等植物骨壁碎屑含量一般较低。这些高等植物骨壁碎屑可以为丝质体(惰质体),也可以在细菌凝胶化作用下形成镜质体[67]。
3 生物骨壁壳有机碎屑的生烃气潜力
从上述骨壁壳有机碎屑纤维质、几丁质、骨胶原、肽聚糖和胶质等生物高分子的化学结构和稳定性来看,一是它们在沉积水体稳定的强还原环境条件下相对容易保存下来;二是经过早期成岩变化和成熟后,不具备生油能力或生油能力很低,高成熟—过成熟早期可具有一定的生烃气能力,与显微组分镜质体或基质镜质体的生烃能力相当,一般烃气的转化率低于15%。
3.1 源于高等植物的纤维素+果胶等无效碳水化合物并经过凝胶化的镜质组在高成熟—过成熟阶段具有一定的生烃气能力,最高总烃气转化率一般不超过15%
大量源于高等植物纤维素+果胶等无效碳水化合物并经过凝胶化的镜质组或以镜质组为主体的未成熟—低成熟煤系烃源岩评价和热压模拟实验结果已经证实,烃源岩中源于高等植物的纤维素+果胶等无效碳水化合物经过凝胶化的镜质组在高成熟—过成熟阶段最高总烃气转化率一般不超过15%,属典型的Ⅲ型有机质类型[71-73]。

图4 水生高等植物纤维质细胞骨壁碎屑化石Fig.4 Fibrous cell wall and bone detritus fossils of aquatic higher plants
例如,贵州凯里鱼洞煤矿上二叠统龙潭组(P2l)以均质镜质体(86.7%)为主的煤(表1,原始样品有机质类型为Ⅲ型,总有机碳含量63.02%,碳酸盐含量小于5%,VRo为0.86%)常规热压模拟实验结果显示:①以生烃气(主要为甲烷)为主,在过成熟阶段烃气(甲烷)产率约为120 kg/tc,主要是镜质组直接热降解,[121.34-(26.81~1.28)×45%]/121.34=90.53%,45%为根据理论推算和模拟实验结果得到的当古原油密度为1 g/cm3时甲烷的最终碳转化率产生的(表1);壳质组(5%)在成熟阶段生成的可溶有机质(沥青“A”)在过成熟阶段最高烃气产率也只有(26.81~1.28)×45%=11.49 (kg/tc),仅占总烃气量的不到10%;而惰质组(8%)产生的烃气甚微。②烃气主要产生在高成熟—过成熟阶段,约占总烃气量的81%以上,过成熟阶段约占总烃气量的38%以上,如果排除壳质组(5%)在高成熟阶段可溶有机质(沥青“A”)热裂解生成的烃气,均质镜质体主要生烃气阶段是在过成熟早期阶段(VRo约2%~4%)。其成烃气模式见图5。

表1 高等植物形成的煤岩(镜质组为主体)常规热压模拟实验综合数据Table 1 Conventional thermocompression simulation experi-ment data of coal (mainly vitrinite) formed by vascular plants
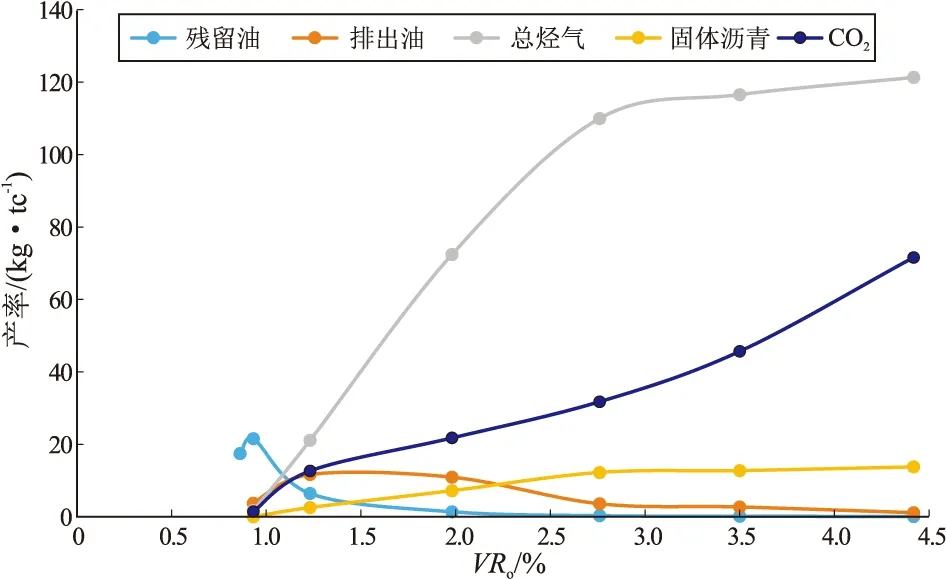
图5 海沼相均质镜质体为主体的煤(Ⅲ型)常规热压模拟生排烃模式Fig.5 Models of conventional thermocompression simulationexperiment of hydrocarbon generation and expulsion on coal(type Ⅲ) dominated by marine and lacustrine homocollinite
也就是说,由纤维质骨架(+果胶)经凝胶化形成的均质镜质体在成熟—过成熟阶段早期是生烃气的,最高烃气产率约120 kg/tc(表1),相当于Ⅲ型干酪根。实际上,基质镜质体、或由纤维质+藻胶等构成的细胞壁壳、或经细菌凝胶化形成的海相镜状体化学结构和成分与均质镜质体相似,只是可能掺杂的或细胞壁中或细菌中的“油”或“烃”含量略高,生烃气能力可能高于均质镜质体,最高烃气产率可达200 kg/tc或更高一些,相当于Ⅱ2型干酪根。但是,遭受氧化或炭化的惰质组或丝质体生烃气能力很差,部分在过成熟阶段生成少量烃气,最高烃气产率均小于100 kg/tc,相当于Ⅲ2型干酪根[70]。
因此,热压模拟实验证实海相优质烃源岩中保存下来的生物骨壁壳有机碎屑在成熟—高成熟阶段一般只具备生成一些凝析气或轻质油气的能力,在过成熟阶段早期仍可生成烃气(甲烷),主要生烃气期是在高成熟—过成熟阶段早期,最大生烃气能力一般不超过150 kg/tc,相当于Ⅲ型干酪根[70]。
3.2 海相优质烃源岩中常见的生物骨壁壳有机碎屑在成熟阶段不具备生油能力,在高成熟—过成熟阶段可以生成一些烃气,相当于Ⅲ型干酪根
(1)从上述纤维质、几丁质、骨胶原、肽聚糖和胶质等生物骨壁壳有机碎屑的化学结构和化学成分来看,除了结构相对稳定并容易保存下来之外,一是它们并非“石油”,即使成岩脱水热变化后其化学成分仍与石油成分不一致;二是它们的某些(芳)环上还带有甲基或乙基等,在高成熟—过成熟阶段在不同程度上能够生成一些天然气,但由于这些芳环上的甲基或乙基等(碳数多小于4)相对需要较高的能量才能热裂解下来;三是尽管它们并非“石油”,有的细胞壁有时含有少许脂类(蜡、脂肪等),在高成熟—过成熟阶段也能够热裂解生成一些天然气。因此,其生烃潜力相当于镜质体或基质镜质体,一般烃气的总转化率低于15%,相当于Ⅲ型有机质类型的生烃潜力。
(2)在海相优质烃源岩中保存下来的生物骨壁壳有机碎屑(纤维质、几丁质、骨胶原、肽聚糖和胶质等生物有机高分子)多为动物类有机骨壳碎屑,如笔石壳屑镜状体、底栖藻类丝壁碎屑、浮游藻类和疑源类纤维质细胞壁及其碎屑、真菌地衣类的几丁质或纤维质丝壁碎屑、细菌类肽聚糖的壁壳及其分泌物形成的胞外聚合物、水生高等植物纤维质细胞骨架壁碎屑以及镜状体、镜质体、丝质体等,它们在低成熟—成熟阶段的热解氢指数一般小于200 mg/g,其H/C原子比都小于1.2(表2),属典型的Ⅲ型有机质类型。它们在热演化生烃过程中,热解氢指数在高成熟—过成熟阶段早期迅速降低,即海相优质烃源岩中保存下来的生物骨壁壳有机碎屑在该阶段早期仍具有一定的生烃气能力。

表2 不同生物骨壁壳有机碎屑地球化学特征Table 2 Geochemical characteristics of organic detritus from different organisms (mainly skeleton, cell wall and shell)
4 结论
(1)对生物体起支持或保护作用的有机骨壁壳等生物高分子主要是由纤维素、几丁质、果胶、肽聚糖等无效碳水化合物和硬蛋白等非活性蛋白质构成,化学结构稳定,不溶于有机溶剂和水,在优质烃源岩形成过程中以非脂类的不溶有机质形式保存下来。
(2)有机骨壁壳及其碎屑包括浮游藻类及疑源类纤维素+果胶质的细胞壁;褐藻等底栖藻类纤维素+藻胶质的细胞壁、真菌类几丁质或纤维质的细胞壁和细菌类肽聚糖(或杂多糖)的细胞壁;甲壳类等节肢动物几丁质的骨壳、笔石类等半索动物骨胶原的皮壳和部分原生动物纤维素+几丁质的外壳;水生陆生高等植物纤维素+果胶质的或纤维+木质+半纤维+果胶质的细胞壁及蛋白质纤维质的细胞骨架。
(3)生物骨壁壳有机碎屑属非脂类的生物高分子不溶有机质,不具备生油能力,在高成熟—过成熟早期具有一定的生烃气能力,烃气转化率一般低于15%,为Ⅲ型有机质类型或与镜质组生烃气能力相当。
