女性白塞病患者血清CD4、CD14免疫细胞中差异表达基因筛选及生物信息学分析
2020-12-02张静邢卫斌
张静,邢卫斌
天津市第五中心医院,天津300450
白塞病(BD)是口腔阿弗他溃疡、外生殖器溃疡和虹膜炎三联综合征,可导致全身多系统病变。该病发病机制不明确,有研究发现,免疫因素在BD发病中的作用不容忽视[1]。目前关于BD的免疫学研究文献并不多,多为BD合并其他疾病的病例报道或极少数遗传易感基因方面的研究[2]。有学者曾发现,在BD患者外周血中,某些关键酶类会同时在CD14、CD4免疫细胞表达上调,而且这些酶类与BD发病密切相关[3]。鉴于BD在成年女性中发病率较高,2019年5月,本研究利用公共数据库中基因芯片数据和生物信息学分析方法,观察女性BD患者与正常人群血清CD4、CD14免疫细胞基因表达谱差异,旨在进一步明确BD患者的免疫学发病机制。
1 资料与方法
1.1 样本及数据来源 本次数据通过GEO数据库(https://www.ncbi.nlm.nih.gov/gds)检索,检索截止时间2019年5月。通过此次检索发现了BD相关数据集GSE61399,包括BD患者CD4免疫细胞样本9例、CD14免疫细胞样本8例以及对应健康人群的CD4免疫细胞样本3例与CD14免疫细胞样本9例(后文中分别简称为CD4免疫细胞实验组和对照组,CD14免疫细胞实验组和对照组)。所有入选样本均为女性。
1.2 差异表达基因筛选 使用GPL570平台注释信息对芯片数据探针名称进行注释,得到对应的基因名称。对于相同的基因表达数据以取列向均值方式进行合并,得到了原始基因表达数据集。之后使用R软件(https://www.r-project.org/)平台scale函数对其进行归一化处理,通过limma R软件进行差异表达基因的筛选,引入BH方法进行统计学校正。以log2|Fold Change|>2及校正P<0.05者作为差异表达基因。
1.3 差异表达基因通路富集分析 基于KEGG数据库和Reactom数据库,使用toppgene(https://toppgene.cchmc.org/)在线工具对差异基因进行联合通路富集分析。
1.4 差异表达基因GO富集分析 基于GO数据库,使用R软件包clusterProfiler v3.8.1对差异表达基因进行GO生物过程(GOBP)、分子功能(GOMF)、细胞组分(GOCC)的富集分析,在结果筛选时使用BH校正并筛选错误发现率(FDR),FDR<0.05的富集功能项作为最后结果。
1.5 核心基因筛选 使用String数据库(https://string-db.org/cgi/input.pl)对所有差异表达基因建立相互作用网络,并选取可信度在0.9以上的基因互作对。之后将这些互作对导入Cytoscape v3.6.1软件(http://www.cytoscape.org/)中,建立差异基因相互作用网络,使用节点筛选工具,将相互作用网络筛选节点度≥10的基因作为核心基因。
2 结果
2.1 差异表达基因筛选结果 CD14免疫细胞实验组和对照组的差异表达基因1 013个,其中506个表达上调、507个表达下调。CD4免疫细胞实验组和对照组的差异表达基因2个(NOXA1、MEGF8),均表达下调。
2.2 CD14免疫细胞差异表达基因KEGG和GO富集分析结果 由表1~4可见,大量差异基因参与了膜运输通路及囊泡介导的转运通路;大量基因参与了蛋白酶体分解代谢过程、RNA的剪接、定位等生物学过程,参与核膜构成,以及关键酶的结合、修饰、激活等。CD4免疫细胞差异表达基因数量少,无法做功能及通路富集。
2.3 核心基因筛选结果 将CD14免疫细胞组差异表达基因使用String数据库匹配建立基因相互作用网络,在此之中也将CD4免疫细胞组差异表达基因引入,观察其所处网络节点的重要性,筛选出节点度≥10的基因(核心基因)分别为RPS27A、GTPBP4、NDUFAB1、MRPL18、NIFK、MRPS5、MRPL46(节点度分别为16、12、12、10、10、10、10)。
3 讨论
BD是一种以血管炎为病理基础的慢性、复发性、多系统受累的全身性疾病。女性外生殖器溃疡很容易与外阴炎性溃疡相混淆,甚至曾经有宫颈受累的BD的报道[4],所以BD容易被误诊、误治。BD发病机制尚不明确,有学者[5]在建立的BD眼受累致盲小鼠模型中发现,CD4免疫细胞表达水平明显高于正常组,并且与IL-2、IL-7水平明显相关。同时,活动期的BD患者CD4免疫细胞中GATA3、TGF-β基因启动子区甲基化水平明显升高,且与其mRNA表达水平呈负相关关系;而GATA3表达水平进一步影响IL-4表达水平,从而导致CD4免疫细胞分泌更多细胞因子参与BD发病过程[6]。

表1 CD14免疫细胞差异表达基因KEGG分析结果

表2 CD14免疫细胞差异表达基因GOBP富集分析结果

表3 CD14免疫细胞差异表达基因GOCC富集分析结果
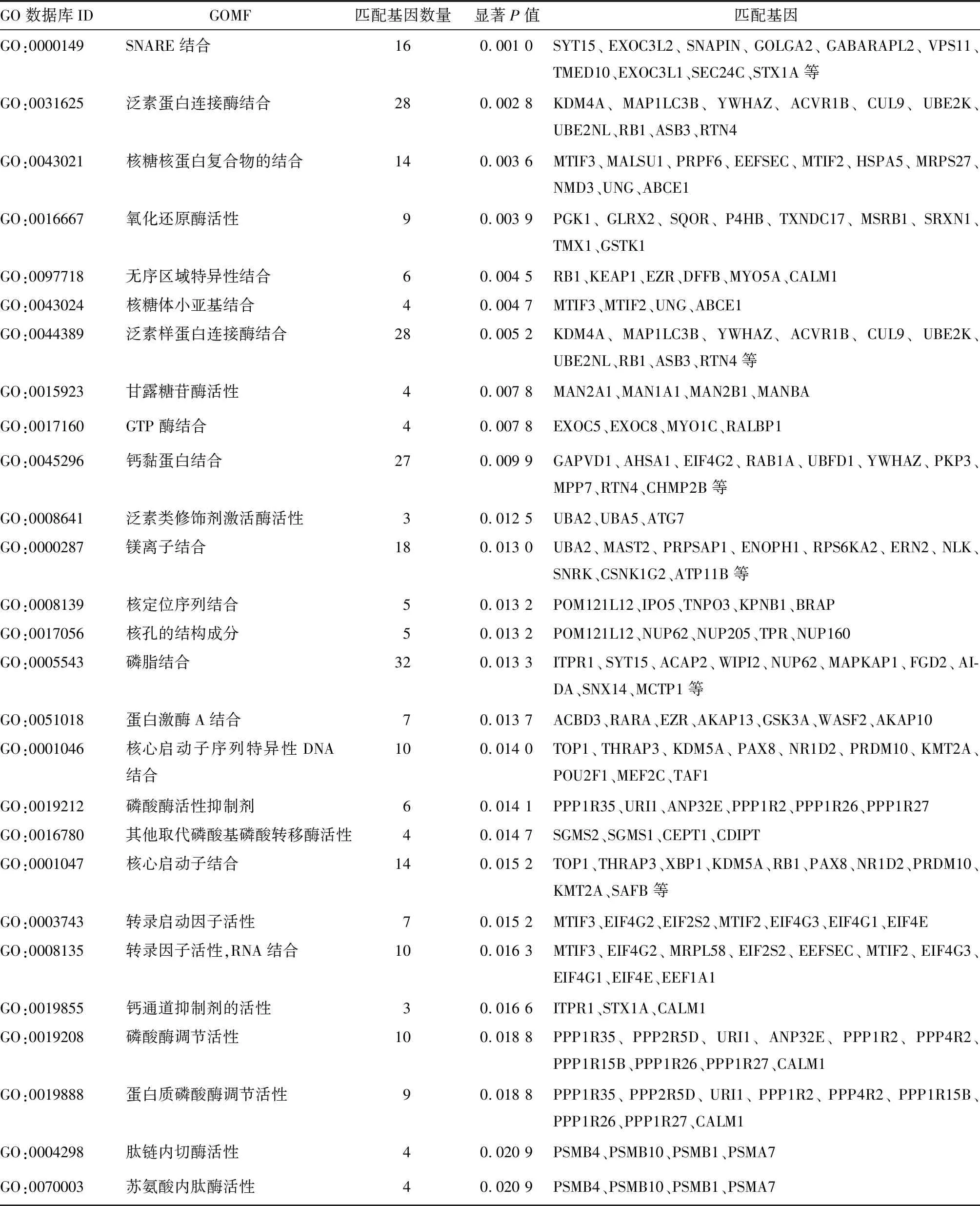
表4 CD14免疫细胞差异表达基因GOMF富集分析结果
本研究发现,CD4免疫细胞组筛选出的差异基因仅有2个,可能与样本量少、可用数据有限有关。然而,本研究发现CD14免疫细胞显著性差异基因数量非常大,可以推测在BD致病因素中,CD14免疫细胞可能发挥着极为重要的作用。CD14是一种主要存在于单核细胞、巨噬细胞等细胞表面的白细胞分化抗原,作为LPS/LPS结合蛋白复合物受体与其结合,发挥细胞毒作用、介导炎症反应。有研究显示,BD患者血清可诱导CD14免疫细胞表型出现极化反应,产生M1样巨噬细胞,从而发挥更强的病理学效应[7]。国内有研究发现,与BD有着同样病理学基础的重度子痫前期的产妇分娩后蜕膜组织和正常产妇蜕膜组织相比,CD14免疫细胞的表达水平明显增高[8]。本研究发现,更多的CD14免疫细胞差异表达基因主要富集在RNA剪接转运等通路,参与蛋白酶体蛋白分解过程。因此,我们有理由推测BD患者是通过这些差异表达基因上调CD4和CD14免疫细胞的表达水平,从而引起它们释放更多的细胞因子和细胞毒性物质,并且这些基因也可能直接参与血管内皮细胞损伤及细胞吞噬过程,最终导致血管闭塞、坏死,引起BD相关症状。
本研究中,与健康对照组相比,BD患者血清CD14免疫细胞中差异表达基因1 013个,以下调基因所占比例较高,其中RPS27A、GTPBP4下调最为明显;在上调基因里以NDUFAB1最为显著。而CD4免疫细胞实验组和对照组之间差异表达基因只有2个,这与以往文献报道有所不同,有待后续进一步探索。目前关于以上差异表达基因在BD中的作用研究极少,较多研究集中在肿瘤方面。例如,RPS27A是一种核糖体蛋白,除了参与蛋白质合成,还参与转录、修复、细胞发育和凋亡调控等多种生物学过程,在肝癌、骨肉瘤、宫颈癌等细胞中均有过量表达[9]。BD患者中RPS27A表达下调,这一点符合BD的病理特征,肿瘤组织需要更多的血液供应,与BD的血管闭塞坏死过程截然相反。但RPS27A基因是否通过间接调控CD14+免疫细胞表达水平从而对血管内皮细胞产生影响抑或直接调控血管内皮细胞增殖、凋亡而导致BD发生还需要进一步证实。研究显示,GTPBP4基因沉默后,结肠癌细胞增殖能力降低,迁移距离缩短,凋亡率高,GTPBP4可能通过调节p53和survivin蛋白的表达来实现上述效应[10]。因此我们推测,BD患者GTPBP4基因下调也可能通过以上机制导致血管内皮细胞凋亡加速从而致病。本研究发现的表达上调基因NDUFAB1是参与线粒体能量和活性氧(ROS)代谢的重要调节器,可以有效保护高脂饮食小鼠免受肥胖和胰岛素抵抗的影响,也可以减少ROS对心脏的损伤;而在BD患者中,NDUFAB1过度表达是否因为免疫炎症反应刺激而反馈性启动机体保护机制,进一步使血管内皮细胞免受损伤,还需要进一步证实[11,12]。
综上所述,BD患者CD4、CD14免疫细胞中差异表达基因主要参与膜运输通路及囊泡介导的转运通路,参与蛋白酶体分解代谢、RNA剪接、定位等生物学过程,参与核膜构成,以及关键酶的结合、修饰、激活等。筛选出与BD发病相关的核心基因主要有RPS27A、GTPBP4、NDUFAB1、MRPL18、NIFK、MRPS5、MRPL46。
