水稻早熟大穗品系7-37二次枝梗数的QTL定位研究
2020-11-27江振方王复标孙惠敏
郑 卓,江振方,王复标,孙惠敏
水稻早熟大穗品系7-37二次枝梗数的QTL定位研究
郑 卓,江振方,王复标,*孙惠敏
(井冈山大学生命科学学院,江西,吉安 343009))
通过水稻穗部性状的改良尤其是增加二次枝梗数更有助于实现水稻的大穗和高产。本研究以7-37作为二次枝梗数QTL定位的亲本材料,通过小穗广亲和品种02428与7-37杂交并自交获得02428/7-37F2,然后对该F2群体中随机选取的 284个单株主茎穗的二次枝梗数进行遗传分析,结果表明二次枝梗数呈典型的正态分布,说明二次枝梗数受多基因控制,属数量性状,与前人的研究结果是一致的;在此基础上,利用从339对SSR引物中筛选出的均匀分布于12条染色体上的 58对多态性引物对F2群体中的284个单株进行PCR扩增分析,再利用QTL Icimapping作图软件,采用复合区间作图法对7-37的二次枝梗数进行了QTL分析和作图,结果在4号染色体25.0~30.5 Mb区间内和6号染色体6.6~30.8Mb区间内分别定位出了二次枝梗数相关QTL位点,LOD值分别为5.73和2.56,贡献率分别为 8.95%和4.38%,加性效应分别为-5.71和-3.85,显性效应分别为-2.83和-1.30。这些研究结果为下一步二次枝梗数QTL位点的精细定位、克隆和功能分析提供了研究基础。
水稻;二次枝梗数;QTL定位
水稻(L.)属禾本科稻属植物,作为稻属中最古老、最具全球性影响力的农作物之一,广泛种植于我国及世界各个地区,为世界上近一半的人口提供了食物[1-2]。水稻在亚洲的种植面积一直在各大洲中稳居首位,是水稻作物的主要种植区域,约占世界水稻种植面积的九成。据国家统计局公布, 2017年我国全年粮食种植面积11798.91万hm2,其中水稻的种植面积约为3074.72万hm2,约占粮食种植面积的26.06%,所以水稻是我国重要的粮食作物。近些年来,国内水稻产出量与需求量趋近于平衡状态,但是我国拥有较大的人口基数,如今再伴随着我国二胎生育政策的推行,人口数量将保持一定的增长趋势。其次随着我国城市化进程的逐渐加快,以及局部地区环境污染日益严重等系列问题,导致实际耕地面积受到一定程度上的挤压和不良影响。故而目前在确保我国耕地面积不减少的基础上,提高水稻产量尤其是水稻的单产对保障我国粮食安全起着非常重要的作用[3-8]。
每穗颖花数的增加是水稻总体产量优势的主要体现[9-13]。然而每穗颖花数的增加程度又主要取决于一次枝梗以及二次枝梗数量的增加[13-16]。要增加每穗颖花数以及稻穗着粒密度最行之有效的途径就是增加一次枝梗数和二次枝梗数,尤其增加二次枝梗数更有助于实现水稻的大穗高产[17-18]。因此,加强对水稻的二次枝梗数的QTL定位研究,解析二次枝梗数形成的分子机制,对于获得更为理想的水稻穗部形态,提高水稻产量具有十分重要的意义。
1 材料与方法
1.1 实验材料的种植
2017年本地(江西吉安)正季(5月25日)播种早熟大穗品系7-37和02428,抽穗时以02428为母本、7-37为父本采用人工去雄的方法获得02428/7-37 F1。2017年12月5 日于海南三亚播种02428/7-37 F1,成熟时收获02428/7-37F1自交F2种子。2018年本地(江西吉安)正季(5月25日)播种亲本02428、7-37以及02428/7-37 F2种子,秧龄25 d,单本插,移栽密度为20 cm×26 cm。其中,亲本02428和7-37每个材料种植5行,每行10株;02428/7-37 F2种植 40 行,每行10株。整个生育期及时防病虫害。
水稻的插秧密度为17 cm×22 cm,单本插,大田常规水作管理,及时防治病虫害和杂草。
1.2 亲本及F2二次枝梗数的考查
成熟后,每个亲本选取中间3行、每行选取1株具有代表性单株的主茎穗作为二次枝梗数调查的材料,3株主茎穗二次枝梗数的平均值分别作为两个亲本的二次枝梗数;在F2群体中去掉边行和病株,共选取284个单株的主茎穗进行二次枝梗数的考察。
1.3 水稻DNA提取方法
于分蘖期剪取两亲本7-37和02428以及杂交构建的F2群体的水稻幼嫩叶片2-3片,迅速放入冰盒带回实验室,采用CTAB法[19]分单株提取亲本及F2单株叶片的DNA。用超微量核算蛋白分析仪(Q5000,Quawell Technology Inc)检测DNA浓度。
1.4 PCR扩增及电泳检测
PCR反应体系为20 µL,其中含2 µL 10×buffer (Mg2+)、0.4 µL 10 Mm dNTP、0.25µL 5 µM Primer 5´、0.25µL5 µMPrimer 3´、0.4 µL 1UEnzyme、0.4 µLDNA模板、余量以ddH2O补足。配制完成后,使用PCR仪(Bio-Rad)进行扩增。
PCR反应程序为:95 ℃预变性5 min;94 ℃变性30 s,50~60 ℃(随引物的不同而变化)退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸5 min。
PCR扩增产物用2.5%的琼脂糖凝胶电泳或8.0%非变性聚丙烯酰胺凝胶电泳(PAGE)检测。在检测中,根据多态性片段的大小及其差异程度,其中片段差异较大的(>20 bp)选择用2.5%的琼脂糖凝胶进行电泳分离检测,电泳缓冲液为1×TAE,电压为100 V,电流为100 A,电泳时间为30 min;差异较小的(< 20 bp)选择用8.0%非变性聚丙烯酰胺凝胶电泳检测,电泳缓冲液为1×TBE,电压为250 V,电流为120 A,电泳时间为3 h,电泳完毕银染显色。
1.5 数据处理
利用Microsoft Excel2016软件对二次枝梗数次数作分布图以及其他相关性分析。在此基础上,对双亲本及F2群体的单株使用实验室订购的339个覆盖水稻基因组的SSR标记进行多态性筛选,结果发现其中有83对SSR标记表现出较为明显的多态性。分别利用筛选出的83对与目的基因可能存在连锁关系的分子标记,从中每个染色体上选择多态性强的5对分子标记(11号和12号上各选择了4对),共计58对分子标记,接着用这些分子标记对随机抽取的0284株单株样本进行基因型检测,通过凝胶电泳分离PCR产物,并对基因型读数,亲本1型记为“0”,杂合型记为“1”,亲本2型记为“2”,利用QTL IciMapping软件进行数据分析。在利用QTL IciMapping软件进行数据分析时,将基因型数据和二次枝梗性状的表型数据相结合,应用复合区间作图法(composite interval mapping,CIM)进行QTL分析,检测QTL阈值为LOD值大于2.5,同步进行对单个QTL估算贡献率和加性效应值等。
2 结果与分析
2.1 二次枝梗数遗传分析
本次实验中,常规粳稻亲本02428是江苏省农业科学院遗传生理研究所选育的矮秆广亲和粳稻,是由粳稻螃蟹谷与粳稻吉邦谷杂交选育而成[20],平均二次枝梗数为42,而亲本7-37的平均二次枝梗数为64,两者二次枝梗数相差22个,差异显著。对F2分离群体二次枝梗数进行遗传分析,其峰度为0.60,均值为56.54,标准差12.00,呈明显的正态分布(图1,图2,表1),说明二次枝梗数受多基因控制,对二次枝梗数进行QTL定位是可行的。

A: 02428(左)和 7-37(右)穗部性状比较;B: 02428(左)和 7-37(右)二次枝梗数比较
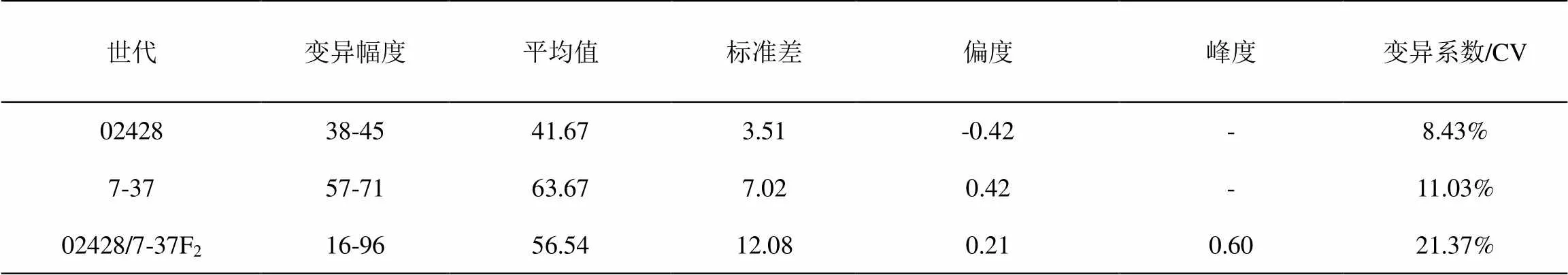
表1 亲本及其F2 群体二次枝梗数的表型分析

图2 02428/7-37F2群体二次枝梗数频度分布
2.2 特异引物筛选结果
本研究所用的SSR标记均来源于Gramene网站(http://www.gramene.org/)。在水稻数据库(https://shigen.nig.ac.jp/rice/oryzabase/)也可查询其序列信息以及在染色体上的物理位置。在覆盖整个基因组12条染色体上设计了339对标记,从中选择每条染色体上多态性强的5对SSR标记(11号和12号上各选择了4对),共筛选出58对分子标记用于二次枝梗数的QTL定位。部分引物筛选见图3。

两两成对,左为亲本02428,右为亲本7-37。引物名称标注
2.3 二次枝梗数QTL初步定位结果
利用均匀分布在12条染色体上的具有多态性的58对SSR引物对F2群体中的 284个单株进行PCR,然后使用QTL IciMapping软件对PCR电泳结果进行数据处理。在数据处理时,应用复合区间作图法(CIM)进行QTL分析,以LOD值大于 2.5作为QTL存在的阈值进行检测(如图4),并获得定位位置图像(如图5),图中qeczg4-1和qeczg4-2标注位置即为QTL定位位置。qeczg4-1和qeczg4-2分别位于4号和6号染色体上,物理距离分别为79.4Mb和45.7Mb,LOD值分别为5.73和 2.56,贡献率分别为8.95%和4.38%,加性效应分别为-5.71和-3.85,显性效应分别为-2.83和-1.30(表2)。

表2 检测到的穗部性状相关QTL
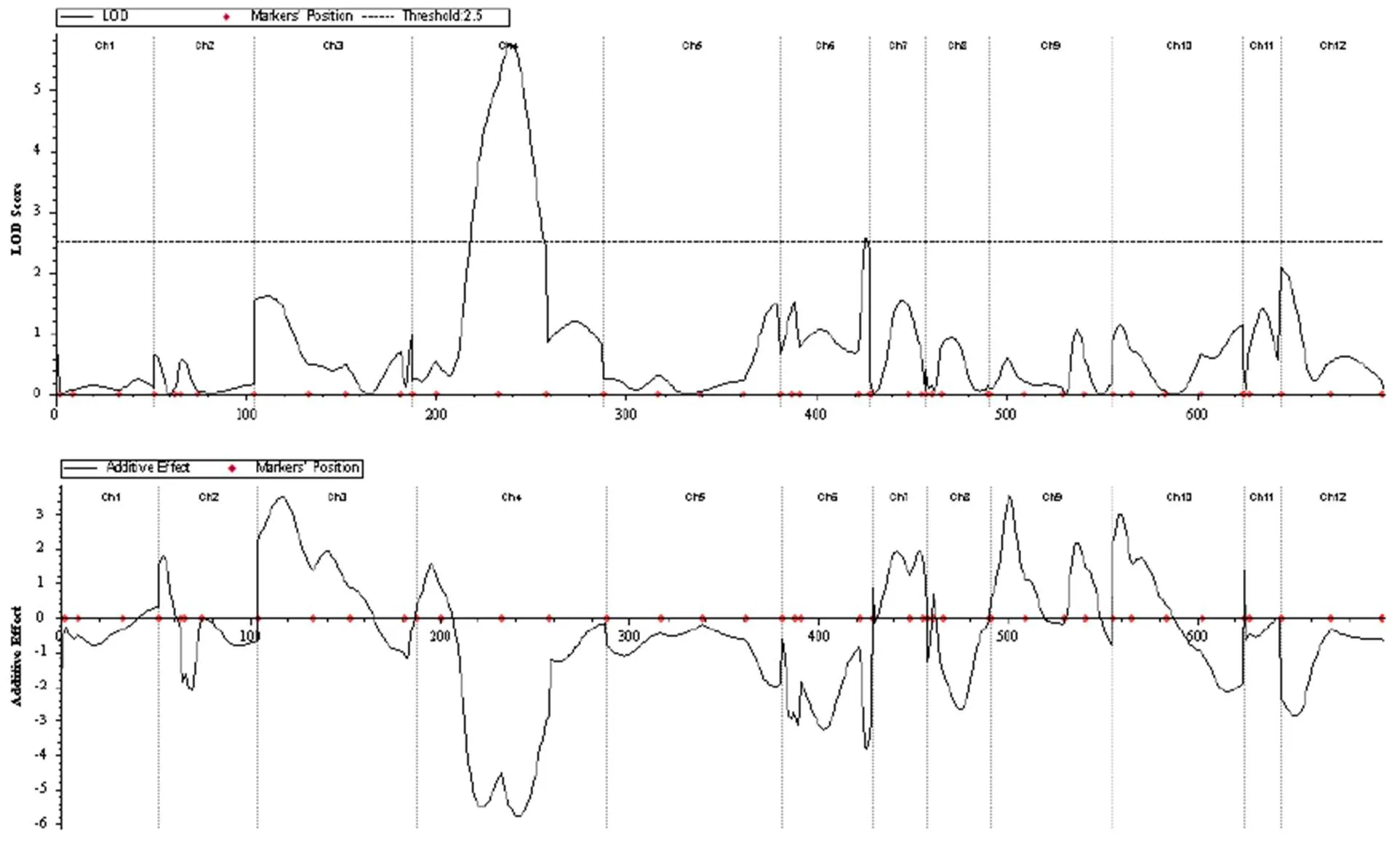
图4 LOD和ADD的检测值

3 讨论
穗部性状是十分重要且复杂的数量性状,将穗部性状进行定向改良是提高水稻产量的重要途径[21-24]。水稻穗部性状为数量性状,在不同遗传群体中检测到的穗长QTL结果差异较大,而且易受到环境影响。例如章志宏[25]等运用经典数量遗传学方法研究后认为控制二次枝梗数性状的基因在6对以上,是由微效多基因控制的。本研究通过图示基因型分析表明,两对被定位的QTL对穗形相关性状具有增效作用,并能促进单株产量的提高。
本研究通过构建大容量的F2遗传群体,对穗部性状进行了QTL分析,结果得到了在第4和第6染色体上的QTL定位的范围。其中4号染色体上的QTL定位在RM5709和RM1388之间,区间的物理距离宽度为6.8 Mb,6号染色体上的QTL定位在RM8242和RM4128之间,区间物理距离宽度为24.2 Mb。通过与前人的研究结果比较分析来看,本次实验所检测到的2个QTL位点,与韩保林[23]等之前研究定位的二次枝梗数QTL位点不同,其没有定位到4号染色体的QTL位点。而本研究定位到6号染色体上的qeczg-6-1 与吴亚辉[24]定位的qSB-6-1区间接近。本次实验丰富了利用不同的作图群体的穗部相关QTL定位数据库,为进一步研究穗部性状QTL提供了参考和方向。
4 小结
本研究利用自选早熟、大穗材料7-37作为二次枝梗数的定位亲本,通过对广亲和粳稻品种02428与大穗亲本7-37F2群体二次枝梗数的QTL定位分析,共检测到2个与二次枝梗数相关的QTL位点,这两个位点分别位于4号染色体上的RM5709-RM1388区间内和6号染色体上的RM8242- RM4128区间内,贡献率分别为8.95%和4.38%。
[1] Yuan L P. Development of Hybrid Rice to Ensure Food Security[J]. Rice Science, 2014, 21(1): 1-2.
[2] 赵凌,赵春芳,周丽慧,等. 中国水稻生产现状与发展趋势[J]. 江苏农业科学, 2015, 43(10): 105-107.
[3] 汪欲鹏,段里成,龙启樟,等. 大穗型水稻穗部性状的QTL定位[J]. 南方农业学报,2016, 47(9): 1445-1449.
[4] 匡勇,罗丽华,周倩倩,等. 水稻籼粳交重组自交系群体穗部性状的相关和遗传分析[J].华北农学报,2011, 26(3): 72-78.
[5] 郎有忠,王美娥,吕川根,等. 水稻叶片形态、群体结构和产量对种植密度的响应[J].江苏农业学报,2012, 28(1): 7-11.
[6] 顾克军,陶君毅,谭立云,等. 永业生命素浸种与拌种对水稻生长及产量的影响[J]. 江苏农业科学, 2012, 40(12): 94-95.
[7] 徐正进,陈温福,张龙步, 等. 水稻高产生理研究的现状与展望[J].沈阳农业大学学报, 1991(S1): 115-123.
[8] 张巍巍,王伯伦,姚乾,等. 水稻穗部性状与产量的关系[J].安徽农业科学, 2009, 37(25): 11906-11908,11936.
[9] 吴光南,张云桥. 稻穗发育过程及其控制途径的研究[J]. 作物学报, 1962(1): 43-52.
[10] Komatsu M, Chujo A, Nagato Y, et al. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets[J]. Development, 2003. 130(16): 3841-3850.
[11] 王昌华,徐正进,夏永胜,等. 水稻穗部性状及其与产量相关分析[J].辽宁农业科学,2007(3): 54-56.
[12] Geldermann H. Investigations on inheritance of quantitative characters in animals by gene markers II. Expected effects[J]. Theoretical and applied genetics, 1976, 47(1):1-4.
[13] 彭友林. 水稻穗部性状QTL分析及精细定位研究[D]. 杨州:扬州大学, 2014.
[14] 辛业芸. 水稻产量性状QTL定位研究进展[J]. 湖南农业大学学报:自然科学版, 2007(4): 396-402.
[15] Zeng Z B. Theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci. [J]. Proceedings of the National Academy of Sciences, 1993, 90(23): 10972-10976.
[16] 朱军. 运用混合线性模型定位复杂数量性状基因的方法[J]. 浙江大学学报:自然科学版, 1999(03): 111-119.
[17] 陈达. 水稻穗粒数QTLGN1.2的遗传分析和精细定位[D]. 扬州:扬州大学, 2017.
[18] 周红菊. 籼粳亚种间染色体片段代换系的构建及其产量性状杂种优势效应研究[D]. 武汉:华中农业大学, 2009.
[19] 孙惠敏,夏米,陈清玲,等. 基于InDel分子标记的20个水稻品种的籼粳成份分析[J]. 井冈山大学学报:自然科学版, 2016, 37(6):100-106.
[20] 严宗卜,葛必庆,曹绍书,等. 耐寒杂交稻新组合K优267[J]. 杂交水稻, 2004(2): 77- 78.
[21] 陈明亮,熊焕金,胡兰香,等. 水稻产量相关数量性状基因研究进展[J]. 江西农业学报, 2014, 26(12): 16-20.
[22] 董桂春,居静,于小风, 等. 不同穗重类型常规籼稻品种产量形成的差异研究[J]. 扬州大学学报:农业与生命科学版, 2010,31(1): 49-54.
[23] 韩保林,顾朝剑,张洪凯,等. 水稻穗部性状的QTL定位及上位性分析[J]. 分子植物育种,2017,15(6): 2218-2227.
[24] 吴亚辉,陶星星,肖武名,等. 水稻穗部性状的QTL分析[J]. 作物学报, 2014, 40(2): 214-221.
[25] 章志宏,李平,王玲霞,等. 与水稻籼粳分化有关的穗颈维管束性状基因的分子标记定位[J].遗传学报, 2002, 29(11): 995-1000.
QTL MAPPING OF SECONDARY PANICLE BRANCH NUMBER OF A PREMATURE AND LARGE-EAR VARIETY 7-37 IN RICE
ZHENG Zhuo, JIANG Zheng-fang, WANG Fu-biao,*SUN Hui-min
(School of Life Sciences, Jinggangshan University, Ji'an, Jiangxi 343009, China)
The improvement of panicle traits, especially the increase of the numbers of secondary branches, is more conducive to the realization of the large ear and high yield of rice. In this study, 7-37 was used as the parental material for QTL mapping of secondary panicle branch number. The hybrid between the variety 02428 and 7-37 was carried out by spikelet broad-affinity, and 02428/7-37F2was obtained by self-crossing of the hybrid 02428/7-37F1. The numbers of secondary panicle branches of each main stem panicle selected from the F2population randomly were genetically analyzed. The results showed that the numbers of secondary panicle branches were normally distributed, indicating that the number of secondary branches was controlled by multiple genes and belonged to quantitative traits. On this basis, PCR was performed on the 284 individual plants selected from the F2population randomly by the 58 pairs of the SSR primers which were uniformly distributed on 12 chromosomes, and QTL analysis and mapping were performed on the secondary panicle branches number of 7-37 using the QTL Icimapping software. The results showed that the QTL locus associated with the secondary panicle branche number are located in the 25.0-30.5 Mb range of chromosome 4 and the 6.6-30.8Mb range of chromosome 6, the LOD values are 5.73 and 2.56, the contribution rates are 8.95% and 4.38%, the additive effects were -5.71 and -3.85, and the dominant effects are -2.83 and -1.30 respectively. These results provide a basis for the fine mapping, cloning and functional analysis of the QTL locus of the secondary panicle branch number in the future.
rice; secondary panicle branch number; QTL mapping
S511.2
A
10.3969/j.issn.1674-8085.2020.05.007
1674-8085(2020)05-0031-07
2020-03-06;
2020-05-09
国家自然科学基金项目(31660382)
郑 卓(1973-),男,湖北竹溪人,副教授,博士,主要从事水稻遗传育种研究(E-mail:zhengzhuodai@126.com);
江振方(1998-),男,河南濮阳人,井冈山大学生命科学学院生物科学专业2015级本科生(E-mail:jiangzf123@qq.com);
王复标(1988-),男,江西兴国人,讲师,博士,主要从事植物生理学研究(E-mail:509371055@qq.com);
*孙惠敏(1985-),女,河北唐山人,副教授,博士,主要从事水稻分子标记开发与育种应用研究(E-mail:huimin729@163.com).
