感染水稻瘤矮病毒的转cry1Ab水稻对电光叶蝉种群的影响
2020-11-26高云丽马璐瑶张刘通
苗 鑫,高云丽,马璐瑶,张刘通,张 榜,林 胜
(1.福建农林大学闽台作物有害生物生态防控国家重点实验室;2.福建农林大学应用生态研究所;3.教育部害虫生态防控国际合作联合实验室;4.农业部闽台作物有害生物综合治理重点实验室,福建 福州 350002)
水稻是我国第一大粮食作物,其产量占粮食总产量的45%以上,因此,水稻在我国粮食生产和农业发展中具有举足轻重的地位[1-2].然而,虫害问题始终是制约水稻稳产、高产的一个重要因素.目前,培育和使用抗虫水稻品系是一个经济、安全、有效的控制害虫的手段[3],其中,使用转基因技术培育抗虫品系是一种切实有效的方法.我国已经培育出一系列能够抗虫并具有优良品质的转基因水稻[4].虽然转基因抗虫水稻能够有效控制害虫的发生、减少化学农药的投入、降低农药对环境的污染[5-6],但是其生态安全性问题一直备受关注,其中一个主要的安全问题是转基因抗虫水稻环境释放后是否会对非靶标生物产生非预期的影响.
研究表明,转基因抗虫水稻对稻飞虱、叶蝉等非靶标害虫种群发生发展影响不显著[3,7],但感病后的转基因抗虫水稻会对非靶标昆虫相关指标产生显著影响.例如:感染水稻矮缩病毒(Ricedwarfvirus,RDV)的Bt水稻会显著影响黑尾叶蝉(Nephotettixcincticeps)卵历期、雌雄成虫鲜重、雄成虫寿命等指标[8];感染南方水稻黑条矮缩病毒(Southernriceblack-streakeddwarfvirus,SRBSDV)的Bt水稻(T1C-19)会显著缩短褐飞虱(Nilaparvatalugens)雌成虫的发育历期[9].
电光叶蝉(Reciliadorsalis)是水稻上重要的经济害虫,直接刺吸为害水稻,并能传播水稻主要病毒.水稻瘤矮病毒(Ricegalldwarfvirus,RGDV)在我国南方多个省份的稻区发生,尤其在广东省存在长达30年常态流行,严重时造成局部水稻减产50%以上[10].该病毒在我国主要由电光叶蝉以持久增殖型方式传播[11].电光叶蝉取食病株后最短在1 h内获毒,即成带毒虫,在虫体内经4~12 d循回期即具有侵染致病力,能终身传毒,若虫的传毒效力比成虫强,成虫吸食健株30 min 内即可传毒给健株而致病[12].目前,有关转基因水稻对非靶标害虫电光叶蝉的安全评价报道十分鲜见.因此,本试验研究感染RGDV的转cry1Ab水稻对电光叶蝉种群的影响,以期为进一步明确转基因抗虫水稻的生态风险和推进其商品化提供依据.
1 材料与方法
1.1 供试昆虫
不带毒电光叶蝉:于2018年8月采自福建省福州市寿山试验基地,经检测未带毒后饲养于人工气候室[温度(28±1) ℃,相对湿度70%±1%,光照14L∶10D],连续在TN1水稻上饲养5代以上.带RGDV电光叶蝉:福建农林大学病毒研究所饲养品系,经RT-PCR检测确认其携带RGDV,于2017年9月开始在福建农林大学闽台作物有害生物生态防控国家重点实验室饲养[温度(28±1) ℃,相对湿度70%±1%,光照14L∶10D].将该虫饲养在2~3 cm的TN1水稻幼苗上,每隔8 d将其转移到新鲜的水稻幼苗上,确保其获取充足的营养.带毒电光叶蝉主要用于水稻传毒.
1.2 供试水稻及其处理
转cry1Ab水稻(MF86)及其亲本非转基因水稻闽恢3301(MH3301)种子由福建省农业科学院农业遗传工程重点实验室提供.
播种前在室温条件下浸种24 h,室温催芽48 h,待露白后进行旱育秧.于每个塑料盆中装入适量土壤,整理平整后将胚芽向上放入,再覆盖一层土壤(团粒),置入人工气候箱中[温度(28±1) ℃,相对湿度70%±1%,光照14L∶10D]生长.10 d后取长势相当的秧苗,移栽至30 mm×200 mm的玻璃试管中,试管中盛有pH为4.5~5.0的Kimura B水稻营养液[13],每管置入2株稻苗,然后在每个试管内放入带RGDV电光叶蝉4~5龄若虫10头,试管顶端用丝袜罩住,放入人工气候室内,待2 d后除去虫源.除虫后在同样条件下继续培养,并每隔3 d更换营养液,10 d后分单株,取少量水稻叶组织进行RGDV检测,阳性植株为感RGDV水稻植株,用于后续试验.另外,取同一批水稻植株,在相同培养条件下经不带毒电光叶蝉取食处理,获得未感病RGDV水稻植株.
1.3 方法
选取150头不带毒电光叶蝉雌雄成虫(性比接近1∶1)分别置于4组水稻上,即感病MH3301、未感病MH3301、感病MF86、未感病MF86,待产卵24 h后取出水稻.将各组水稻单独置于养虫笼中,每天观察水稻上电光叶蝉卵孵化情况,并将初孵若虫转移到对应的处理组水稻上,进行单虫单苗单管饲养,每个处理组需80个虫体,每天记录若虫蜕皮情况、羽化情况,并将同一处理组的电光叶蝉进行雌雄两两配对.剩余的初孵若虫继续饲养于各组水稻上,用于后期配对不均时候补.每天更换新苗,并在显微镜下观察产卵量,直至虫体死亡.
1.4 统计与分析
统计卵、若虫、成虫的发育历期,以及产卵前期、产卵期、产卵后期、日繁殖率、最大繁殖率、达最大繁殖率时的成虫年龄和总繁殖力等生殖相关参数[14];种群生命表参数包括特定年龄-龄期存活率(Sxj,表示种群中个体能存活到年龄x到龄期j的概率)、种群特定年龄存活率(lx,表示Sxj曲线在不考虑龄期分化情况下的简化形式)、种群年龄繁殖率(mx,表示种群在年龄x时的平均产卵量)、雌成虫年龄-龄期繁殖率(fxj,表示一头雌成虫在年龄x和龄期j所产的能够孵化的卵粒数)、种群年龄净增殖率(lxmx)、种群世代净增殖率(R0,表示种群在一定条件下经过一个世代后的每个存活个体所产生的后代总数)、内禀增长率(r,表示种群的最大瞬时增长率)、周限增长率(λ,表示种群内平均每一个个体能产生的后代数)、平均世代周期(T,表示在稳定的龄期阶段分布中,种群增长到其规模的R0倍所需要的时间长度)[15-16].
采用双因素方差分析水稻品种、RGDV及二者交互作用对电光叶蝉发育历期、生殖相关参数和种群生命表参数的影响.另外,采用单因素方差分析电光叶蝉发育历期、生殖相关参数和种群生命表参数在不同处理间的差异,并采用LSD进行多重比较.采用年龄-龄期两性生命表分析软件TWOSEX-MSChar中的Timing-MSChart预测以100粒电光叶蝉有效卵为起始的100 d种群动态[16].
2 结果与分析
2.1 感染RGDV的转cry1Ab水稻对电光叶蝉发育历期的影响
水稻品种对电光叶蝉发育历期的影响不显著;RGDV显著影响了电光叶蝉的卵及1、2、3龄若虫和雌成虫的发育历期以及总寿长;RGDV和水稻品种对卵期、1和5龄若虫期有显著的交互效应(表1).与取食未感病MH3301相比,取食未感病MF86的电光叶蝉1和5龄若虫期及总寿长均显著延长;与取食未感病MF86相比,取食感病MF86的电光叶蝉1、2、3龄若虫期及雄成虫期和总寿长显著缩短;与取食感病MH3301相比,取食感病MF86的电光叶蝉卵期及1、2、3龄若虫期均显著缩短(表2).

表1 电光叶蝉发育历期双因素方差分析结果Table 1 Outputs of two-way ANOVA analysis of the developmental duration of R.dorsalis
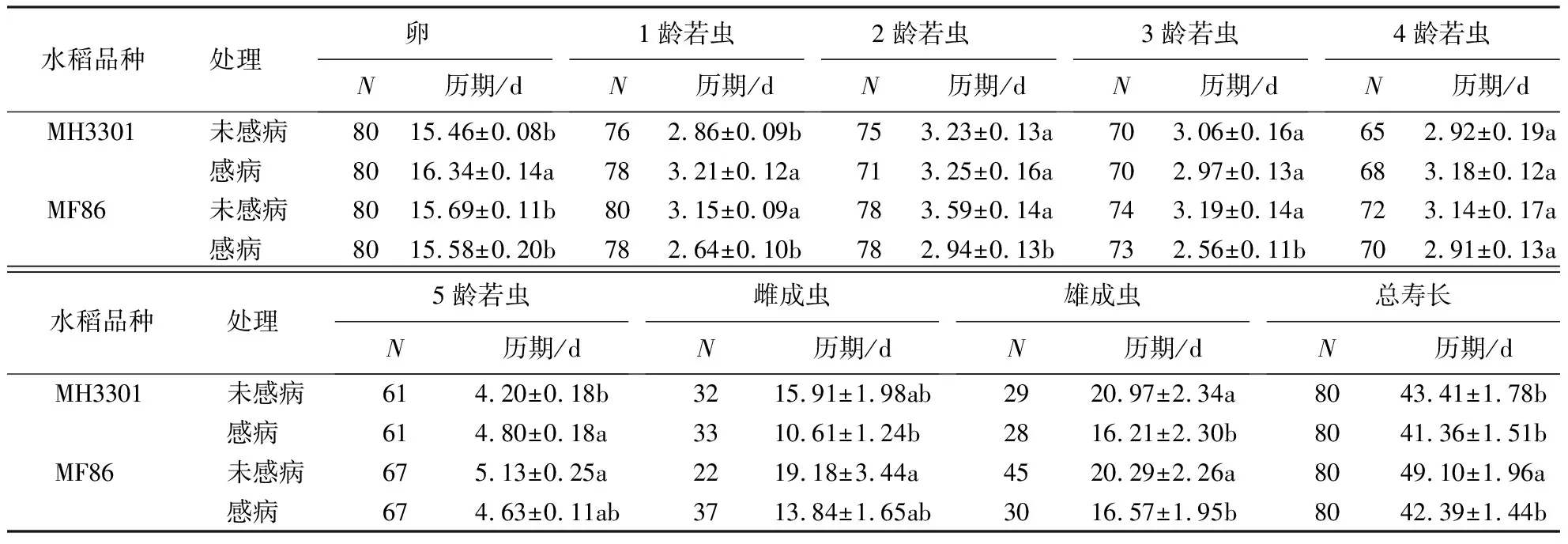
表2 电光叶蝉取食感病或不感病水稻后的发育历期1)Table 2 Developmental duration of R.dorsalis feeding on healthy or RGDV-infected rices
2.2 感染RGDV的转cry1Ab水稻对电光叶蝉生殖相关参数的影响
水稻品种对电光叶蝉产卵前期有显著影响;RGDV对电光叶蝉产卵期、产卵后期、达最大繁殖率时的成虫年龄有显著影响;RGDV和水稻品种对生殖相关参数无显著交互效应(表3).与取食未感病MH3301相比,取食未感病MF86的电光叶蝉产卵期显著延长;与取食未感病MF86和感病MH3301相比,取食感病MF86的电光叶蝉产卵前期和产卵期显著缩短(表4).

表3 电光叶蝉生殖相关参数双因素方差分析结果Table 3 Outputs of two-way ANOVA analysis of the reproductive parameters of R.dorsalis
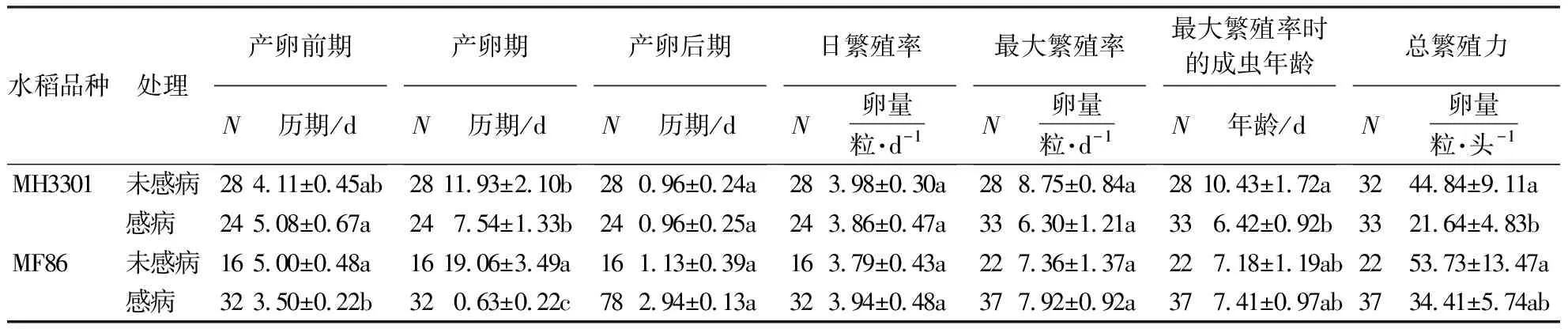
表4 电光叶蝉取食感病或未感病水稻后的生殖相关参数1)Table 4 Reproductive parameters of R.dorsalis feeding on healthy or RGDV-infected rices
与取食未感病MH3301相比,取食未感病MF86的电光叶蝉fxj、mx和lxmx在产卵前期表现较高,45 d后有所下降,最长存活时间延长(图1A、1C);与取食未感病MF86相比,取食感病MF86的电光叶蝉fxj表现较低,50 d后fxj和mx均有所上升,但最长存活时间缩短(图1C、1D);与取食感病MH3301相比,取食感病MF86的电光叶蝉fxj、mx和lxmx均较高,最长存活时间延长(图1B、1D).
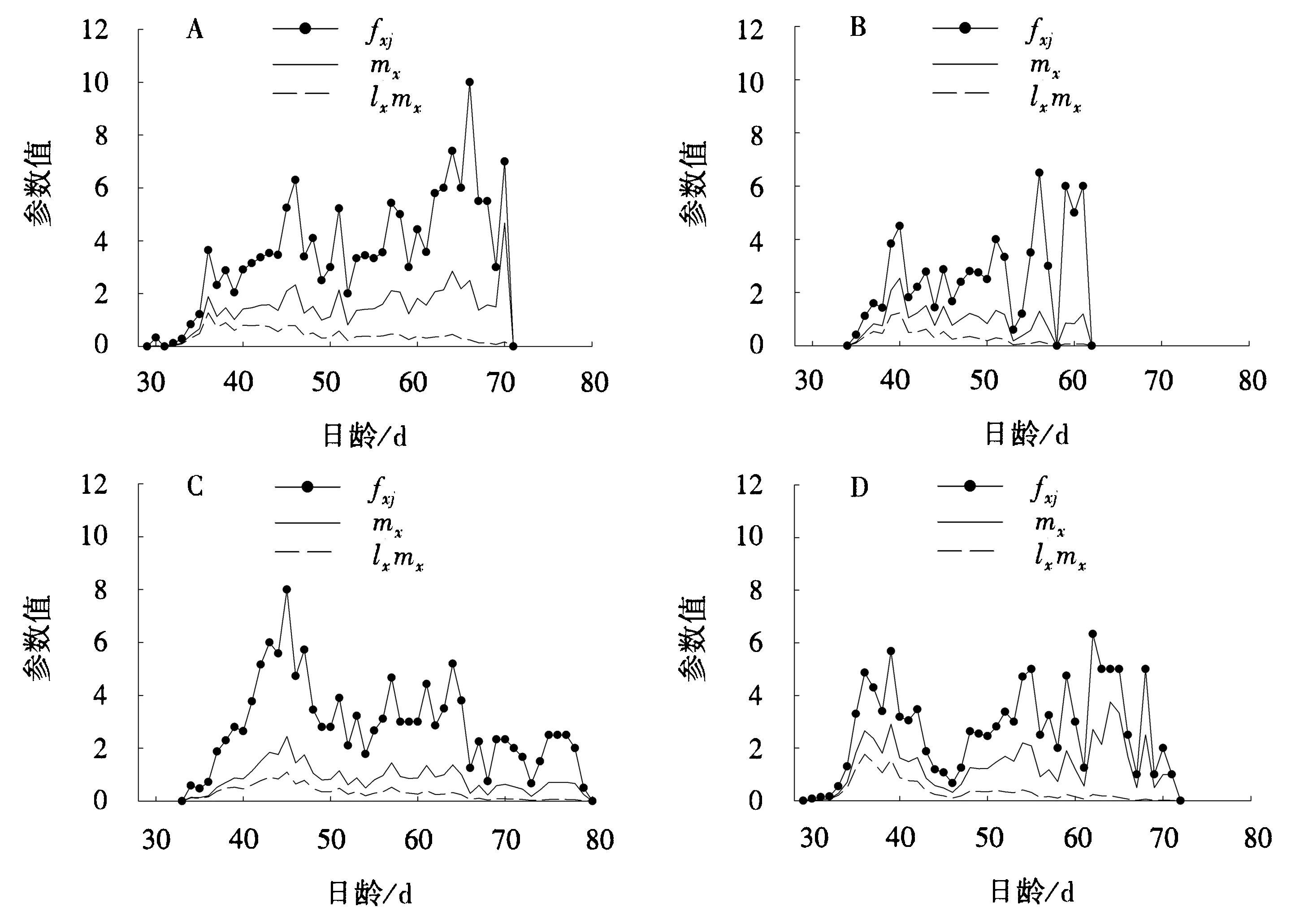
A.未感病水稻MH3301;B.感病水稻MH3301;C.未感病水稻MF86;D.感病水稻MF86.图1 电光叶蝉雌成虫取食感病或未感病水稻后的年龄-龄期繁殖率(fxj)、种群年龄繁殖率(mx)和种群年龄净增殖率(lxmx)Fig.1 Age-developmental stage specific fecundity (fxj),age-specific fecundity (mx),and age-specific maternity (lxmx) of female R.dorsalis feeding on healthy or RGDV-infected rices
2.3 感染RGDV的转cry1Ab水稻对电光叶蝉存活率的影响
与取食未感病MH3301相比,取食未感病MF86的电光叶蝉3~4龄若虫存活率均有所上升,雌成虫存活率有所下降,存活历期总体延长(图2A、2C);与取食未感病MF86相比,取食感病MF86的电光叶蝉卵、1~4龄若虫和雄成虫存活率均呈下降趋势,雌成虫存活率有所升高,存活历期总体呈缩短趋势(图2C、2D);与取食感病MH3301相比,取食感病MF86的电光叶蝉卵、1~4龄若虫和雄成虫存活率均有所下降,雌成虫存活历期有所延长,雄成虫存活历期有所缩短(图2B、2D).

A.未感病水稻MH3301;B.感病水稻MH3301;C.未感病水稻MF86;D.感病水稻MF86.图2 电光叶蝉取食感病或未感病水稻后的特定年龄-龄期存活率(Sxj)Fig.2 Age-developmental stage specific survival rates (Sxj) of R.dorsalis feeding on healthy or RGDV-infected rices
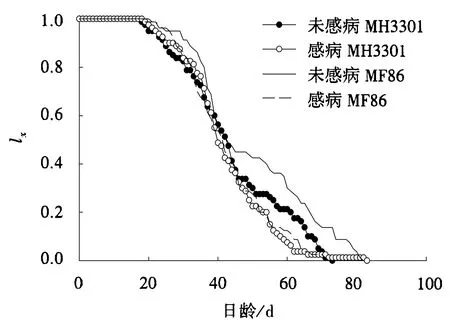
图3 电光叶蝉取食感病或未感病水稻后的种群特定年龄存活率(lx)Fig.3 Population age-specific survival rates (lx)of R.dorsalis feeding on healthy or RGDV-infected rices
与取食未感病MH3301相比,取食未感病MF86的电光叶蝉存活率较高,存活历期较长;与取食未感病MF86相比,取食感病MF86的电光叶蝉存活率较低,存活历期较短;与取食感病MH3301相比,取食感病MF86的电光叶蝉存活率和存活历期基本一致(图3).
2.4 感染RGDV的转cry1Ab水稻对电光叶蝉种群动态相关参数的影响
水稻品种对电光叶蝉的r、R0和T有显著影响;RGDV对电光叶蝉的λ、R0和T有显著影响;二者对这4个参数均有显著的交互效应(表5).与取食未感病MF86相比,取食感病MF86的电光叶蝉T显著缩短;与取食未感病MH3301相比,取食未感病MF86的电光叶蝉r、λ、R0和T均无显著变化;与取食感病MH3301相比,取食感病MF86的电光叶蝉r和R0均显著升高(表6).
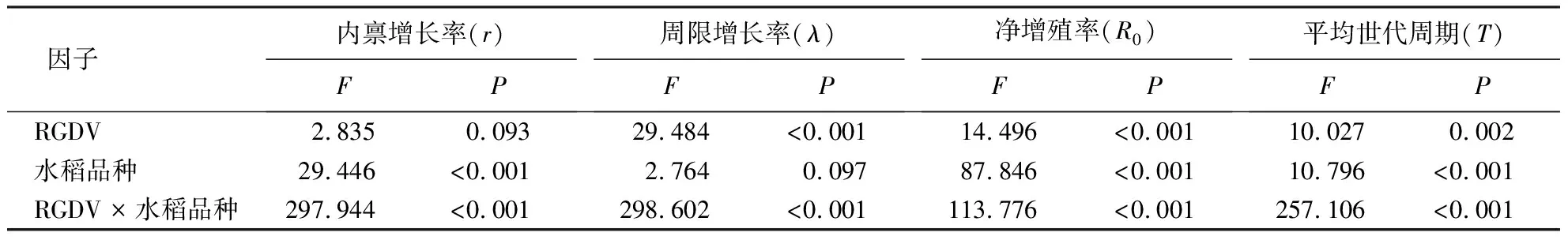
表5 电光叶蝉种群参数双因素方差分析结果Table 5 Outputs of two-way ANOVA analysis of the population parameters of R.dorsalis

表6 电光叶蝉取食感病或未感病水稻后的种群参数1)Table 6 Population parameters of R.dorsalis feeding on healthy or RGDV-infected rices
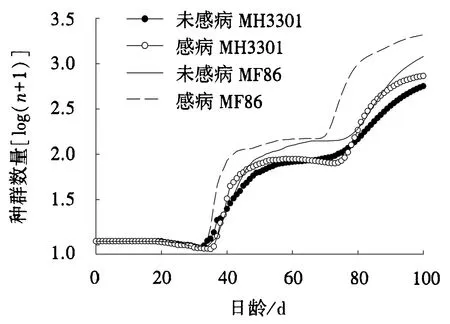
图4 取食感病或未感病水稻的电光叶蝉种群动态预测Fig.4 Predictions on the population dynamics of R.dorsalis feeding on healthy or RGDV-infected rices
取食不同处理水稻的电光叶蝉均在38 d左右出现第1代,76 d左右出现第2代.之后开始出现差异,与取食未感病MF86相比,取食感病MF86的电光叶蝉种群预测世代周期明显快,种群预测曲线较高.MF86与亲本MH3301相比,对电光叶蝉的影响发生显著变化:与取食未感病MH3301相比,取食未感病MF86的电光叶蝉种群增长速度快,增长率高;与取食感病MH3301相比,取食感病MF86的电光叶蝉种群增长速度快,增长率高(图4).
3 讨论
外源cry基因在水稻染色体中的随机插入,可能会导致转基因抗虫水稻某些物理性状以及营养成分等发生改变,从而对植食性昆虫的生长发育、取食和产卵行为产生影响[17].陈茂等[18]研究发现,转cry1Ab基因的抗虫水稻对褐飞虱产卵量无显著影响;周霞等[19]研究表明,转cry1Ab基因水稻会对非靶标昆虫的产卵期和产卵量产生影响;徐雪亮等[20]研究发现,转Bt基因水稻“赣绿1号”不会对非靶标昆虫的生长发育产生不利影响.本试验发现,与取食未感病的非转基因水稻MH3301相比,取食未感病的转cry1Ab基因水稻的电光叶蝉产卵量无显著变化,而产卵期显著延长.可见,相同的转基因水稻会对不同的非靶标昆虫产生不同的影响.
当水稻受到病毒的入侵时,会产生防御反应以及改变营养成分,从而在一定程度上影响植食性昆虫的生长发育.Tu et al[21]研究表明,取食感染SRBSDV的水稻后,无毒的和携带SRBSDV的白背飞虱(Sogatellafurcifera)成虫寿命显著缩短,生殖力下降.本试验也发现,与取食未感病MH3301相比,取食感病MH3301的电光叶蝉总繁殖力显著降低.与此相反,将B型烟粉虱(Bemisiatabaci)分别饲养于健康和感染木薯花叶病毒(Cassavamosaicvirus)的木薯植株上,结果发现病株上的烟粉虱种群数量明显高于健康植株上的数量,且植株在田间的发病程度与烟粉虱数量呈正相关,即病毒能提高介体昆虫的生殖能力[22].此外,本试验发现,与取食未感病MF86相比,取食感病MF86的电光叶蝉雄成虫寿命及总寿长均显著缩短.He et al[23]也发现,取食感染SRBSDV的转基因水稻后,白背飞虱雌成虫寿命和卵历期显著缩短,而取食感染SRBSDV的非转基因水稻后白背飞虱的适应性提高.因此进一步说明,受病毒侵染的植株会对非靶标昆虫的生理生化特性造成一定的影响.
王前进等[24]研究发现,取食感染RDV的水稻后,非靶标昆虫黑尾叶蝉净生殖率显著提高.本研究发现,与取食未感病MH3301相比,取食未感病MF86的电光叶蝉种群参数无显著变化;与取食感病MH3301相比,取食感病MF86的电光叶蝉内禀增长率及净增殖率均显著升高.种群预测结果表明,取食感病或未感病MF86的电光叶蝉均表现为种群增长快且数目多的特点,推断转入基因及感染RGDV改变了水稻的特性,从而对非靶标昆虫的种群产生影响.此外,电光叶蝉通过口针吸食植物汁液存活[25],而水稻感染RGDV后,其汁液成分或防御途径可能发生改变,从而对叶蝉的营养利用或取食行为造成影响,导致叶蝉的寿命、存活率等生物特性发生变化[26].其具体机制有待进一步研究.
