基于轮廓分析的电针长强穴对FMR1基因敲除小鼠CREB表达的多脑区联动效应研究*
2020-11-25齐诗仪张志灿章思佳林丽莉
齐诗仪,张志灿,林 燊,章思佳,林丽莉,林 栋
(福建中医药大学针灸学院 福州 350122)
脆性X 综合征(Fragile X syndrome,FXS)是脆性X智力低下基因(Fragile X mental retardation 1,FMR1)发生突变引起FMR1 mRNA 低表达或不表达,导致患儿出现中度或重度的智力障碍。目前FXS 尚无有效疗法,产前诊断和选择性流产是其主要的预防手段。因此,越来越多的研究者关注到了传统针灸疗法,其通过体表干预从而改善精神类疾病[1]。《难经·二十八难》云:“督脉者…上至风府,入属于脑”。说明督脉与脑联系密切,故有“病变在脑,首取督脉”之言。本课题组在前期临床研究中通过针刺小儿脑性瘫痪伴智力低下患儿的督脉络穴、初始之处—长强穴,发现其对改善患儿认知功能障碍具有显著的疗效[2]。同时,在前期动物实验中,本课题组通过研究与FRX 患者具有相似特征,精神发育迟滞的重要动物模型FMR1 基因敲除小鼠[3-4],发现针刺长强穴分别影响了海马、皮质、小脑等不同脑区的相关蛋白表达量改变,从而改善突触可塑性[5-7]。那么,针刺长强穴所引起的不同脑区间的整体效应又是如何?故本研究拟通过观察FMR1基因敲除小鼠海马、皮质、小脑的不同脑区之间环磷腺苷效应元件结合蛋白(cAMP-response element binding protein,CREB)、磷酸化CREB(p-CREB)蛋白的表达情况,以进一步研究针刺的脑功能效应。
1 材料与方法
1.1 实验动物
将FMR1 基因敲除小鼠(美国The Jackson Laboratory 公司)进行配种与繁殖,并采用聚合酶链式反应(Polymerase chain reaction,PCR)检测技术测定新生小鼠的基因表型,选择纯合子基因型小鼠进行实验。小鼠饲养于福建中医药大学实验动物中心SPF级房间(许可证号:SYXK(闽)2014-001)。实验过程严格遵照科技部[2006]398 号《关于善待实验动物的指导性意见》文件要求执行。
1.2 试剂与仪器
水合氯醛(福州迈新生技术开发有限公司);华佗牌0.5寸针灸针(苏州医疗用品厂有限公司);HANS-200A韩氏穴位神经刺激仪(北京华运安特科技有限责任公司)等。
1.3 方法
1.3.1 PCR鉴定动物基因表型
对通过配种繁殖所得的48 日龄新生小鼠进行颌下静脉丛采血。在血液样品种加入Buffer TBP(血液样品:Buffer∶TBP=1∶2),充分摇匀后离心去上清,加入TE Buffer 离心去上清数次。在沉淀物中加入Buffer Digestion 和Proteinase,震荡混匀后进行56℃水浴1 h。样品加入Buffer PR 混匀后放置-20℃冰箱20 min。随后将样品室温离心取上清,在上清液中加异丙醇混匀后室温静置,后离心去上清。75%乙醇清洗后室温离心去上清,重复两次后室温静置10 min,加入TE Buffer并提取DNA。将提取的DNA 进行1%琼脂糖电泳(150 V、100 mA 20 min)并于化学发光凝胶成像系统中拍照观察。FMR1基因敲除小鼠可扩增出350 bp 的DNA片段,杂合子可扩增出350 bp和180 bp的DNA 片段,引物序列见表1。
1.3.2 动物分组与干预方法
将通过PCR 技术筛选出纯合子基因敲除小鼠,随机分成三个组,每组8 只。(1)空白组仅模拟抓取动作。(2)非经非穴组选择肋弓最低点上1 cm 作为干预部位。(3)长强组根据《实验针灸学》[8]小鼠长强穴位于尾根与肛门之间的凹陷中。其中非经非穴组与长强组使用自制双极针,进针约10 mm,并连接韩式电针仪,连续波,强度2 mA,频率2 Hz,干预20 min。各组每天于固定时间进行干预,一天一次,干预14 天。
1.3.3 免疫组织化学技术
(1)于干预结束当天采用水合氯醛对小鼠进行腹腔注射,麻醉后对其进行心脏灌注,随后取出全脑进行固定。(2)依次进行不同浓度梯度酒精、二甲苯脱水、石蜡包埋。(3)将蜡块切片、捞片和烘片。(4)将切片进行脱蜡、透明、水化后进行抗原修复。(5)使用纯水冲洗切片三次后晾干,加入过氧化氢,室温下孵育10 min。PBS 溶液冲洗三次,加入山羊血清,室温下孵育 10 min。(6)去残液加一抗,室温下孵育 60 min。PBS溶液冲洗三次,加即用型快捷免疫组化试剂,室温下孵育15 min。PBS 溶液冲洗三次,去残液加LDAB显色液,室温下孵育5 min。显微镜观察后用纯水洗净,苏木素复染,PBS 冲洗,梯度酒精脱水,二甲苯透明,最后用中性树胶封片。
1.4 统计学方法
本研究使用SPSS 22.0软件分析软件对数据进行分析,所得的数据以()表示,组间比较采用单因素方差分析及最小显著差法(LSD-t),并对不同脑区的各指标变化采用轮廓分析,以P<0.05 为差异具有统计学意义。
2 结果
2.1 PCR基因鉴定结果
图1泳道可见400 bp目的片段,为纯合子小鼠,即FMR1 基因缺失。而杂合子小鼠泳道为350 bp 和180 bp,即FMR1基因存在。
2.2 免疫组化法检测
在光镜下标记细胞胞浆内充满棕褐色的阳性反应颗粒,为圆形或椭圆形且体积较大。
2.2.1 CREB在不同脑区的表达变化
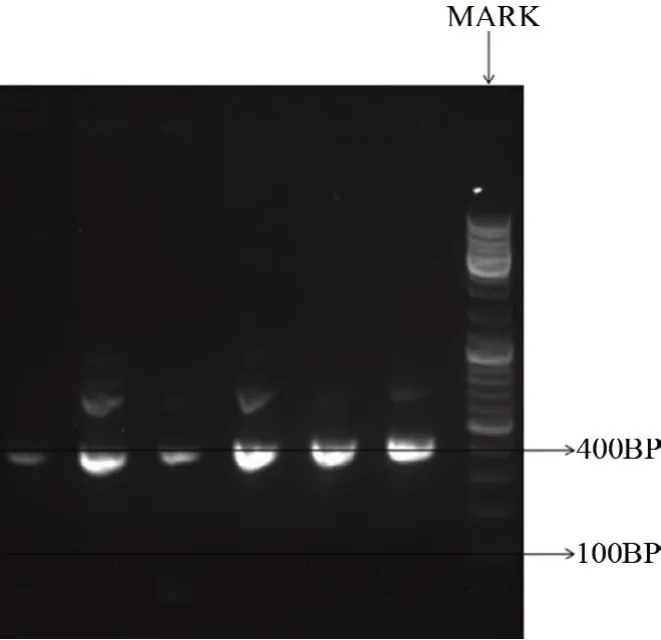
图1 PCR基因鉴定结果
不同脑区CREB 表达平均光密度值提示(见图2质的CREB 表达量较海马(P<0.05,P<0.01)、小脑(P<0.05)显著升高。
针对脑区与蛋白表达的交互作用分析,平行轮廓分析结果差异有统计学意义(F=5.094,P=0.002<0.01),表明各组CREB 表达在脑区变化轮廓不平行,即脑区的联动变化不一致,由图2 可知空白组在三个脑区的CREB 呈现出不同于非经非穴组、长强组的表达趋势,即针刺组与非针刺组的在三个脑区之间行为模式不同。
2.2.2p-CREB在不同脑区的表达变化

图2 各脑区CREB蛋白阳性目标平均光密度比较(左)及轮廓分析结果图(右)

图3 各组小鼠三个脑区抗体阳性目标平均光密度值比较(10X)
不同脑区p-CREB 表达平均光密度值提示(图4和图3):空白组海马的CREB 表达量较皮质(P<0.05)、小脑(P<0.001)显著升高;非经非穴组与长强组在皮和图5),空白组小脑的p-CREB 表达量较皮质(P<0.01)、海马(P<0.05)显著升高;非经非穴组小脑的p-CREB 表达量较海马(P<0.01)显著升高;长强组各脑区p-CREB表达量无显著性差异(P>0.05)。
针对脑区与蛋白表达的交互作用分析,平行轮廓分析结果差异无统计学意义(F=1.085,P=0.378>0.05),表明各组p-CREB 表达的脑区变化轮廓相互平行—三组的脑区联动变化一致;重合轮廓分析结果差异无统计学意义(F=1.880,P=0.177>0.05),表明各组p-CREB的脑区变化轮廓重合—各组间脑区联动变化程度一致;水平轮廓分析结果差异有统计学意义(F=9.870,P=0.001<0.05),表明各组p-CREB的脑区变化轮廓不水平—各组中三个脑区的蛋白表达不相等,其中小脑最高。
2.2.3p-CREB率在不同脑区的变化
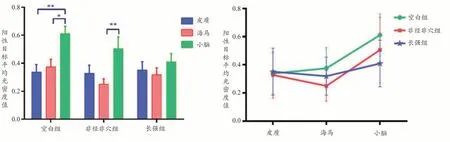
图4 各脑区p-CREB蛋白阳性目标平均光密度比较(左)及轮廓分析结果图(右)

图5 各组小鼠三个脑区抗体阳性目标平均光密度值比较(×10)
不同脑区p-CREB率提示(图6),空白组与非经非穴组在小脑的p-CREB 率较皮质(P<0.001,P<0.01)、海马(P<0.001,P<0.05)显著升高;长强组在各脑区的p-CREB率无显著性差异(P>0.05)。
针对脑区与p-CREB 率的交互作用分析,平行轮廓分析结果差异无统计学意义(F=1.803,P=0.148>0.05),表明各组p-CREB 率在脑区变化轮廓相互平行—三组的脑区联动变化一致;重合轮廓分析结果差异无统计学意义(F=2.465,P=0.109>0.05),表明p-CREB 率在三个脑区表达轮廓重合—各组间脑区p-CREB 率程度基本一致;水平轮廓分析结果差异有统计学意义(F=12.126,P<0.001),表明p-CREB 率在各脑区变化轮廓不水平—各组中三个脑区p-CREB 率不相等,其中小脑最高。

图6 各脑区p-CREB率比较(左)及轮廓分析结果图(右)
3 讨论
3.1 长强穴针刺效应脑区的CREB表达特点
CREB 参与学习、长时程(Long-term potentiation,LTP)记忆的相关过程[9],并介导了神经的保护作用,其在成瘾、抑郁、焦虑等方面发挥着不可忽视的作用[10]。同时,CREB 被认为与早期海马神经干的分化、存活和迁移有关[11],其核蛋白作为细胞内酶信号级联系统以及效应基因的反式激活系统的关键信号枢纽,故大多数神经元细胞内信号能激活CREB 使其磷酸化[12],磷酸化的CREB以二聚体的形式与CRE目标基因序列结合[13],通过调控神经营养因子的表达及其诱导的基因转录,从而在组织细胞修复、再生中起重要作用[14]。故在针灸的脑功能效应研究中CREB 逐渐成为关注的焦点。
3.1.1 皮质
额叶皮质作为认知功能的核心区域,有学者[15]发现其在算术与第二语言流利性的研究中激活明显。而本课题组前期研究表明针刺长强穴上调FMR1基因敲除小鼠皮质脑源性神经营养因子(BDNF)、CREB 等相关蛋白表达[7],并在浅针干预失眠患者的临床研究中通过多元统计分析方法提取到了额颞枕区的特征脑电图(EEG)信号[16]。
3.1.2 海马
作为重要的信息处理区域的海马,其与认知、情绪、行为、学习、记忆以及位置导航等功能密切相关[17]。故诸多研究者们在如抑郁症等疾病研究中发现干预体表穴位能够引起海马区CREB的改变[18-19],而本课题组在针刺FMR1基因敲除小鼠及端粒酶基因敲除小鼠的实验研究中亦观察到了海马CREB 等相关蛋白表达量增加。
3.1.3 小脑
随着生命科学的不断发展,研究[14]表明小脑涉及了语言、时间感以及空间记忆等认知过程,因此越来越多的研究者们关注到了针刺对小脑及其与大脑之间关联的影响。有学者[20]通过脑功能影像学观察到针刺引起小脑的活动变化,这与本课题组前期通过分子生物学技术观察针刺长强穴对FMR1基因敲除小鼠小脑的影响变化结果一致[6]。同时,本课题组采用多元统计分析方法观察了针刺即刻与针刺后20 min小脑与扣带回的影响[21],其研究结果表明针刺增强p-CREB 蛋白表达,且该变化在小脑与扣带回之间存在较为一致的时间变化规律。
有鉴于此,本研究通过采用多元统计分析手段对FRM1基因敲除小鼠皮质、海马及小脑的CREB及其磷酸化蛋白进行研究,以揭示不同脑区之间针刺效应的变化特征。
3.2 针刺长强多脑区联动效应的研究
组化结果显示长强组与非经非穴组的皮质CREB表达量均较海马与小脑高且差异具有统计学意义,而空白组海马区的CREB 表达量较皮质与小脑高且差异具有统计学意义,同时其轮廓分析结果进一步说明了长强组、非经非穴组同空白组在三个脑区中CREB 蛋白表达变化趋势不平行(图4)。有学者[22]利用血氧水平依赖功能磁共振成像(BOLD-fMRI)观察到在静息状态时大脑存在着较试验状态的高激活区域,该区域与海马密切相关。这与本研究结果一致,在无干预状态(空白组)fMR1 基因敲除小鼠海马区CREB 表达量较其他脑区高。而针刺刺激(长强组、非经非穴组)引起脑区的活动状态发生改变,形成以皮质为高激活区域,小脑相对增高,海马相对降低的脑功能活动状态。该结果亦与脑功能影像学研究结果相呼应:针刺穴位引起左侧额上回、小脑扁桃体等区域的负激活[20]。也就是说,本研究结果提示CREB 蛋白的表达量改变与脑功能影像学结果相仿,其对针刺刺激做出了响应。然而效应不同于响应,产生响应不一定具有效应[23],故而本团队认为其可能作为响应蛋白与穴位效应并无较密切关系。
在p-CREB 及其磷酸化率的轮廓分析的结果显示空白组、非经非穴组与长强组在三个脑区变化趋势呈现重合而不水平,形成以小脑为高激活区域,海马与皮质相对较低的脑功能活动状态。方差分析结果进一步显示其在空白组与非经非穴组呈现小脑较皮质和(或)海马显著升高,但其在长强组中三个脑区之间差异无统计学意义。初步表明三组中磷酸化蛋白及其变化率的脑区变化趋势基本一致,且小脑可能参与了认知能力的加工处理过程,这与本课题组前期的针刺对小脑蛋白表达影响的结果一致[6]。同时,在脑区变化趋势基本一致的前提下,在非穴位的组别中(即空白组与非经非穴组)磷酸化蛋白(率)差异具有统计学意义,而穴位组(长强组)差异则无统计学意义,这提示针刺长强可能出现穴位效应,并通过蛋白磷酸化而体现。
3.3 穴脑功能效应研究
近年来,针灸的脑功能效应中枢联动机制研究逐渐成为了穴脑功能研究的热点(图7)。诸多研究者通过脑功能影像学技术探索针刺的脑功能中发现“功能连接(Functional connectivity)”现象[24],即针灸作用于体表穴位产生疗效并非由单一脑区完成,而是由多个脑区协同调节共同发挥作用[25]。同时,越来越多学者关注到了“大脑-小脑环路”是小脑参与认知的基础[26-27],其动态交互作用与短时空间记忆关系密切[28-29]。然而,通过检测脑区之间血氧水平依赖从而判定时域相关性的脑功能影像学仅能提示针刺刺激体表的响应脑区。同时,目前大多数采用分子生物学技术探索针灸穴脑效应的研究多采用某一较有限的局部指标,如某蛋白的表达、某递质的水平等,以及采用较为单一的方差分析以探寻效应机制的差异中特定蛋白或基因的特异性响应,或衡量针灸方案取得的效果,而缺乏对脑功能整体性效应的分析。
故为进一步对针刺脑功能效应的时空变化特征进行表述,本课题组前期研究采用了[16,21]多元统计分析方法对研究数据进行了提取与分析(图8),从时间、空间等多维度、多角度对穴脑效应进行探索[30]。本研究通过采用轮廓分析对海马、皮质、小脑不同脑区之间的行为模式改变进行了分析,并呈现其直观的可视化结果。本课题组将该脑区间的行为模式改变称之为多脑区联动效应,即将空间、时间等因素作为研究参数,在针刺作用的某个(某几个)时刻,不同脑区之间的激活模式及其累积效应的呈现。不同于脑功能影像学显示多个脑区响应模式—即“有(无)”的“点亮”行为;脑功能联动效应则涵盖了针刺效应“量”在不同脑区中的程度变化,凸显了针刺后脑区之间的相对变化特征。同时,本研究采用大脑发育程度较成熟的48日龄小鼠,故其存在着对外界刺激更高的反应状态,拥有更强的脑区联动效应。
本研究通过采用轮廓分析的方法对不同脑区之间的行为模式联动改变进行了分析,并呈现其直观的可视化结果。本研究结果初步表明针刺刺激可能通过影响CREB 表达使得fMR1 基因敲除小鼠形成以皮质为高激活区域的脑功能活动状态。而针刺长强穴则引起p-CREB 表达量在三个脑区中特异性改变,提示其可能存在穴位效应。未来,本团队计划通过多元统计分析方法对疾病、时间、空间等因素展开多角度、多维度的分析,以期丰富针灸的脑功能效应中枢联动机制研究。
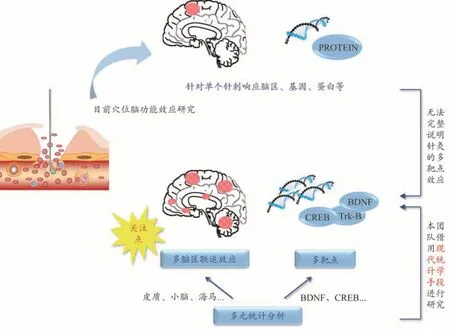
图7 穴脑功能效应研究框架及潜在策略
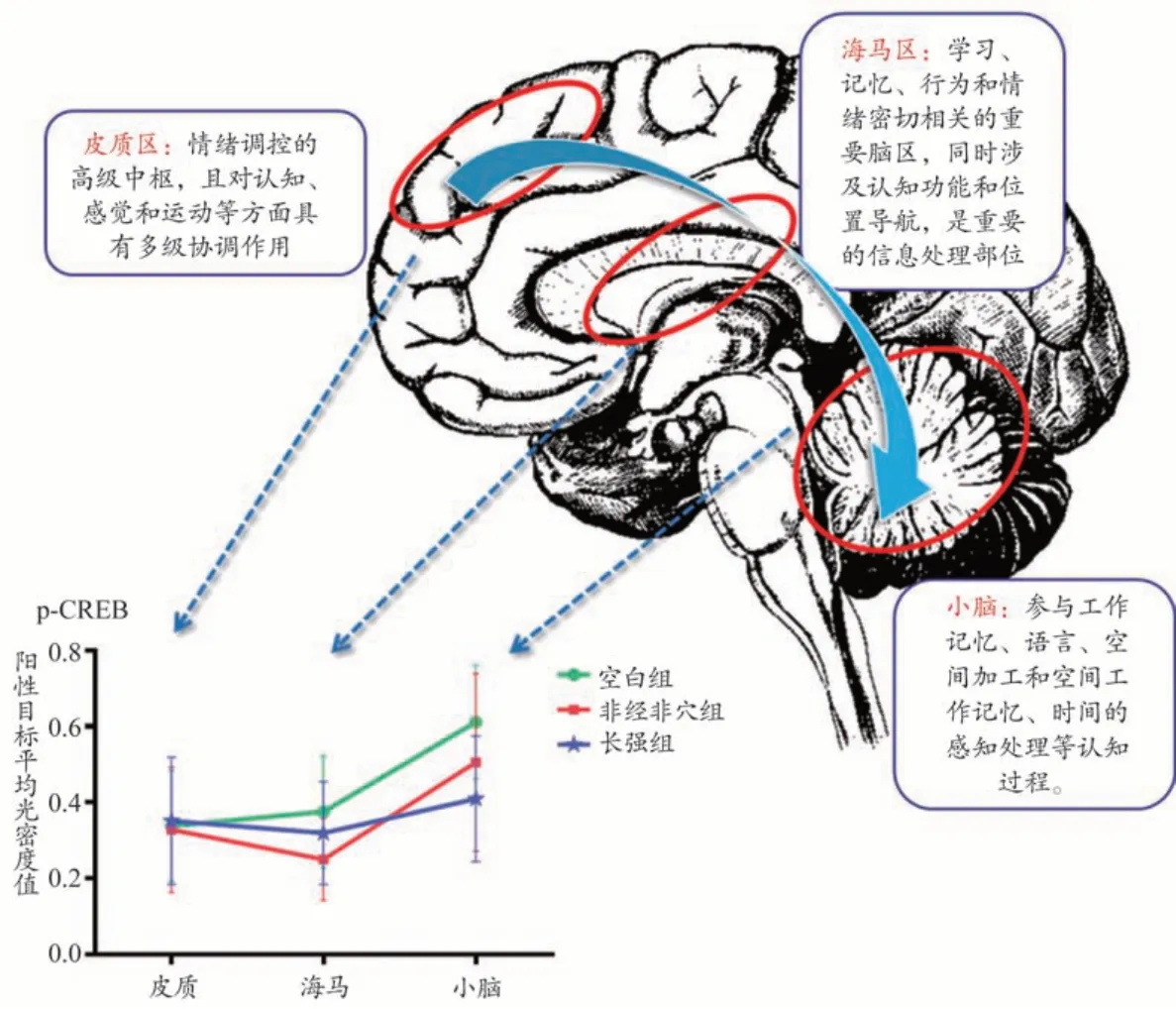
图8 脑区联动效应示意图
