氮添加对不同密度入侵植物喜旱莲子草生长的影响
2020-11-23张运春欧春予张桥英
张运春,欧春予,张桥英
1.西华师范大学环境科学与工程学院,四川 南充 637009;2.西华师范大学国土资源学院,四川 南充 637009
喜旱莲子草(Alternantheraphiloxeroides)是苋科莲子草属的强入侵性草本植物,又称空心莲子草,原产于南美洲,20世纪30年代引入中国,后在人为作用下逸为野生,泛滥于中国南方大部分地区(郭连金等,2009),造成了严重的生态经济损失。喜旱莲子草能够成功入侵水生和陆生环境,主要归功于其表型可塑性(Yang et al.,2019)。
植物的表型可塑性,可以增强入侵植物对多样环境的耐受性和适应性,是入侵植物能够在新生境定殖的原因之一(王桔红等,2014;Yang et al.,2019;曹晶晶等,2020),通过调整植物的表型特性对多变的环境因子产生可塑性响应(Luo et al.,2019),包括植物的形态特性、生理特性和繁殖特性等(刘建等,2010),从而增强了其入侵能力(Spector et al.,2006)。例如,可以通过改变植株的分枝数(Wang et al.,2018)、种子发芽率(Tietze et al.,2019)以及生物量分配格局(潘玉梅等,2017)等来获取生境中的最大资源,达到成功入侵的目的。
近年来,由于工业活动加剧,农田的过度施肥等人类活动导致环境中的氮元素大大增加,使得许多地区土壤中的氮含量过剩,如华南甘蔗种植区氮沉降量超过陆地生态系统的临界负荷(黄金生等,2019)。而氮元素是植物细胞的重要组成元素,大气氮沉降的增加改变了土壤的营养环境,对植物的生长发育产生了一定影响。有研究表明,氮沉降量的增加促进了入侵植物节节麦(Aegilopstauschii)竞争能力的提升,加剧了其入侵性(王宁等,2018)。
植物种群密度依赖效应的实质是植物种群不同个体间对相似资源的竞争作用,种群密度增加,相邻植物对有限的光照、水分和养分的竞争越激烈(林婉奇等,2019)。如在土壤贫瘠条件下,入侵植物银胶菊(Partheniumhysterophorus)扩大种群密度,分枝数减少,茎生物量降低,增大根和叶的生物量,通过茎叶的形态可塑性生长来促进对新生境的入侵(蒲高忠等,2010)。同时,还有研究发现入侵植物水浮莲(Eichhorniacrassipes)种群密度增大,对土壤养分的消耗减少(申时才等,2013),有利于入侵种群的扩散。
目前,关于氮添加与密度交互作用对植物的影响研究已取得相关进展,比如高密度增加桉树(Eucalyptusrobusta)幼苗对茎的碳分配比例,降低了对叶的分配比例,同时氮添加缓解了桉树幼苗高密度下的养分限制(余明等,2020);氮沉降在一定程度上有利于入侵植物豚草(Ambrosia artemisiifolia)在种间竞争中取得优势(陆光亚等,2012)。但是,鲜有文章阐明氮添加与密度交互作用下入侵植物的种内竞争情况。本研究以入侵植物喜旱莲子草为研究对象,探讨氮添加与植株密度的交互作用下种内竞争效应及其对入侵能力的影响,为未来氮沉降环境下喜旱莲子草的入侵机理提供参考。
1 材料与方法
1.1 实验地概况
本实验地设置于四川省南充市顺庆区西华师范大学实验地内(106°4′1″E,30°48′45″N)。属亚热带季风气候,年平均气温17.4 ℃,湿度80%,年降水量1020.8 mm。土地以棕紫泥土和红棕紫泥土为主,土壤有机质含量略偏低。本次实验用土是将当地土壤与营养土(含有少量珍珠岩,优质草炭、泥炭和多种微量元素)以1:4的比例混合后制得。
1.2 实验设计
采集试验地附近喜旱莲子草的茎段(大小、长度基本相同)扦插在培养箱内,在相同的外界环境下生长12 d后,再移植到内径18.5 cm,深11 cm的塑料花盆内。采用两因素随机实验设计,因素一为密度,采用加性实验设计(Additive design)(Oliveira et al.,2018),包括3种密度,分别为低(1 plant·pot-1)、中(5 plant·pot-1)、高(9 plant·pot-1),因素二为氮添加,加氮(N 20 kg·hm-2·a-1,参考Yu et al.,2019)、不加氮。共6种处理(表1),每种处理5个重复。
1.3 采样与测定
实验时间为2019年4月10日—2019年6月30日。实验结束后,测定每盆中心喜旱莲子草的叶量、茎长、茎节数、茎分支数、叶面积(扫描仪扫描后用Image J测量其叶面积);将根、茎、叶分别装入信封袋中,编号,在 65 ℃烘箱中烘至质量恒定后称量其质量,记录各部分的生物量。测定指标包括:(1)形态:叶量(Number of leaves)、茎长(Stem length)、茎节数(Number of stem nodes)、茎分支数(Number of branches)、叶面积(Leaf area);(2)生物量:单株生物量(Biomass)、根生物量比(Root mass ratio,RMR)为根生物量与总生物量的比值、茎生物量比(Stem mass ratio,SMR)为茎生物量与总生物量的比值、叶生物量比(Leaf mass ratio,LMR)为叶生物量与总生物量的比值、根冠比(Root:shoot ratio,R/S)为地下生物量与地上生物量的比值;(3)可塑性指数:根据Valladares et al.(2000)计算所测量指标的可塑性指数,即某一变量在不同的氮素或密度条件下最大平均值和最小平均值的差值与最大平均值的比。

表1 实验设计Table 1 Design of the experiment
1.4 数据处理
所有数据用SPSS 23.0进行分析,将部分数据进行合适的数据转换,以满足方差齐性,用双因素方差分析(Two way ANOVA)检验不同植株密度和氮素水平对入侵植物喜旱莲子草的影响的差异性,采用SNK进行多重比较,用Microsoft Excel完成作图。
2 结果
2.1 植株密度和氮素水平对喜旱莲子草形态特征的响应
植株密度和氮素水平对喜旱莲子草的叶量、茎长、茎节数、茎分支数、叶面积均影响显著(P<0.05),植株密度和氮素水平对叶量、茎长和叶面积的交互作用显著(P<0.05)(表2)。由图1可知,密度增加,喜旱莲子草的叶量、茎长、茎节数、茎分支数减小,密度对喜旱莲子草的生长具有明显的抑制作用;不加氮处理显然不利于植物的生长,与加氮处理相比,都有不同程度降低,而这种差异在低密度时表现最为显著;除叶面积外,在低、中密度下,氮素的促进作用较为明显,而高密度下氮素的促进作用不显著。
2.2 植株密度和氮素水平对喜旱莲子草生物量积累和分配的响应
植株密度对喜旱莲子草单株生物量、叶生物量比影响显著(P<0.01),氮素水平对生物量、根生物量比、根冠比、叶生物量比影响显著(P<0.01);植株密度和氮素水平仅对生物量影响的交互作用显著(P<0.01)(表2)。由图2可知,低密度下的植株生物量显著高于中、高密度(P<0.05);低密度时,加氮处理与不加氮之间有显著差异(P<0.05),而中、高密度时则差异不显著(P>0.05)。叶生物量比在低密度加氮处理下具有最大值,加氮处理组之间没有显著差异(P>0.05);在不同密度下,加氮处理的叶生物量比都大于不加氮处理,在中密度时差异最小。不加氮处理的根生物量比都大于加氮处理,在高密度时,差异达到最大;高密度不加氮处理下的根生物量比达到最大值。根冠比具有和根生物量比一样的变化趋势。

表2 植株密度和氮添加对喜旱莲子草形态特征和生物量影响的F值和显著性分析Table 2 F-values and significance levels for factorial ANOVAs of the effects of density and nitrogen on morphological traits and biomass of A.philoxeroides

图1 不同植株密度和氮素添加对喜旱莲子草形态特征的影响Fig.1 Morphological characteristics of A.philoxeroides under different plant densities and nitrogen additions
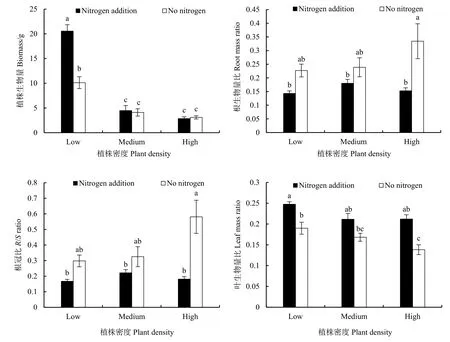
图2 不同植株密度和氮素添加对喜旱莲子草生物量积累和分配的影响Fig.2 Plant biomass and biomass allocation of A.philoxeroides under different plant densities and nitrogen additions
2.3 植株密度和氮素水平对喜旱莲子草可塑性的影响
由表3可以看出,除根生物量比、根冠比、茎生物量比、叶生物量比之外,其他指标都是加氮处理下对密度响应的可塑性指数较大;喜旱莲子草的叶量、茎长、茎节数、茎分支数以及单株生物量随着密度增大,对氮素响应的可塑性指数呈减小趋势;而且对密度响应的可塑性指数要高于对氮素的响应值,说明植株密度对喜旱莲子草形态生长的影响占主导地位。

表3 喜旱莲子草各变量对密度和氮素响应的表型可塑性指数Table 3 Phenotypic plasticity indices for the A.philoxeroides to density and/or nitrogen additions
3 讨论
在本研究中,植株密度对喜旱莲子草的生长表现出显著的负效应,而氮添加表现出显著的正效应,二者对喜旱莲子草的生长产生了明显的交互作用。植株密度增加,喜旱莲子草的叶量、茎长、茎节数、茎分支数减小,而加氮处理都大于不加氮处理,说明氮添加缓解了密度增加的负效应。氮素通过促进喜旱莲子草叶片生长,伸长茎长,产生分支来占据地上资源,增强光合作用,进行生物量的积累,资源利用率大大提高;同时喜旱莲子草产生的分支往往能够形成荫蔽环境抑制其他物种的生长繁殖(全国明等,2014),在生产力较高的生境中,喜旱莲子草占有绝对优势(潘晓云等,2006)。已有研究表明,氮沉降在一定范围内可以促进植物的生长,但氮含量过高会对植物的生长产生负效应(莫凌梓等,2018;樊后保等,2006)。目前的氮沉降量还未超过喜旱莲子草对氮的需求,所以其形态特征和生物量积累还未产生氮过量的负效应。
入侵植物的生物量分配对策对新环境产生的可塑性响应有利于提高其适应性(Portela et al.,2019)。研究表明,喜旱莲子草的入侵策略可能表现为通过调节各构件生物量的投资来增强氮素吸收能力,进而增强其入侵竞争能力(黄成成等,2018)。植株密度增加,不加氮处理的根生物量比增加,而加氮处理之间差异不明显。根冠比具有与根生物量比一样的变化趋势,而叶生物量比变化趋势完全相反。密度增加,种内竞争加强,由于养分的限制,抑制了叶片的生长,为了最大限度地获取必需资源,喜旱莲子草极大地增殖地下部分,促进对营养物质的吸收,满足生长发育;而氮添加为植物提供了充足的养分,喜旱莲子草将更多的生物量分配到碳同化器官,提高对光源的捕获和吸收,增强光合作用,占据地上空间资源,进而提高植物的竞争力(荀挚峰等,2017)。本研究结果与前人对唯一入侵到南极洲的外来物种早熟禾(Poaannua)的研究结果一致,即在恶劣环境下地下器官的生物量比例更高(Rudak et al.,2019)。研究表明入侵植物可通过生物量的再分配(从地下到地上)提高入侵能力(Liao et al.,2019),本研究中喜旱莲子草在加氮条件下也出现生物量的再分配,表明其在氮沉降环境下入侵能力更强。
表型可塑性在生物对异质生境的适应过程中发挥了重要作用(耿宇鹏等,2004)。研究表明,可塑性指数越大,其表型可塑性就越大(杨贺雨等,2016)。在本研究中,喜旱莲子草的叶量、茎长、茎节数、茎分支数、叶面积、单株生物量对密度响应的可塑性指数在加氮处理下较大,说明氮添加有助于喜旱莲子草快速适应种内竞争,促进入侵种群的扩散。与之相反,喜旱莲子草的根生物量比、根冠比、茎生物量比、叶生物量比对密度响应的可塑性指数则是不加氮处理下较大,说明在氮素贫瘠的生境,生物量的分配策略就显得尤为重要,主要通过增大根生物量的比例来积累营养物质供自身生长,以适应资源的缺乏。
4 结论
本研究中,氮添加缓解了密度增加的负效应,削弱了种内竞争的强度,促进喜旱莲子草地上部分的生长来达到成功入侵的目的;另一方面,氮添加改变了喜旱莲子草适应种内竞争的生物量分配策略,促进碳同化器官对光源的利用,有利于在地上资源的竞争中取得优势。由喜旱莲子草在氮添加条件下的一系列可塑性响应可预测未来氮沉降增加会加剧喜旱莲子草的入侵。本研究设置的氮添加量是当前环境中的氮沉降量,但目前氮沉降还有增加的趋势,未来应长期持续地跟踪研究,以期能更准确地探讨喜旱莲子草在氮沉降下的种内竞争效应及其入侵能力变化,为进一步加强对入侵物种的防控提供理论支撑。
