黄连木群落的生物多样性与天然更新
2020-11-18张孟仁苏晓迪张学献叶先亮
张孟仁,苏晓迪,张学献,叶先亮
(1.河南林业职业学院生态工程与旅游系,河南 洛阳471000;2.河南省远志林业规划设计有限公司,河南 郑州450000;3.栾川县林业局,河南 栾川471500)
黄连木(PistaciachinensisBunge)又名鸡冠木、楷树等,为漆树科黄连木属的高大落叶乔木[1-2],其生长缓慢,对生境适应性强,寿命长达百年,在干旱瘠薄、石灰岩地区均能较好地生长,以海拔400~1 200 m的山坡疏林中分布较多[3-4]。黄连木是我国,尤其是太行山区传统的优良用材树种和生物质能源树种[5],还是优良的风景园林树种[6-7]。但由于人为干扰影响,且目前没有针对黄连木群落的科学管理方法,导致其分布面积缩小,群落退化严重且生产力低下。研究黄连木群落的结构、林下植物多样性及幼苗更新特性,对于恢复黄连木的天然群落,开展黄连木群落的保育和可持续经营具有重要意义。
植物群落的天然更新在森林生态系统中是一个复杂的自然现象,也是近自然森林经营的基础,影响着森林中各群落的动态变化,包括物种的组成、结构、更新和繁殖等,在群落演替方面具有不可替代的作用[8-10]。人为干扰活动和自然界其它不利因素,对群落结构与功能、物种数量、种群年龄结构和分布格局等都有不同程度的影响。为了适应外界环境的各种干扰因素,不同物种根据自身生物学特性采取不同的自我更新策略。在我国,黄连木作为研究较多的生物质能源植物之一,目前的研究主要集中在人工选育和栽培技术[11]、生理生态分析[12-13]、生长节律及养分积累[14]等方面。为了研究黄连木群落的自然更新,在对河南省林州市南太行山区的黄连木野生种质资源进行全面调查的基础上,通过对调查区的黄连木群落物种多样性、天然更新方式等方面的分析,发现黄连木的群落特点及幼苗更新规律,为南太行山区黄连木野生种质资源的保育和恢复及可持续经营和管理提供理论依据。
1 调查区概况
调查区位于河南省林州市(属南太行山区),地理坐标为东经113°37′~114°51′,北纬35°40′~36°21′。该区气候属暖温带半湿润大陆性季风气候,雨热同期,年日照时间长达2 800 h以上,年平均气温为10 ℃左右,年平均降水量为534 mm[16-17],土壤类型以沙壤土为主,土层贫瘠,砾石含量高。南太行山区山势险峻,天然群落结构单一,乔木层构成简单,主要以黄连木(PistaciachinensisBunge)、栓皮栎(QuercusvariabilisBlume)、山槐[Albiziakalkora(Roxb.) Prain]为主;灌木层多为黄荆(VitexnegundoL.)、酸枣[Ziziphusjujubavar.spinosa(Bunge) Hu ex H.F.Chow.]、构树[Broussonetiapapyrifera(Linnaeus) L′ Heritier ex Ventenat]等;草本层植物相对种类较多,其中以黄背草(ThemedatriandraForsk.)、艾(ArtemisiaargyiLévl. et Van.)、雀儿舌头[Leptopuschinensis(Bunge) Pojark.]最多。
2 材料与方法
2.1 样地设置
在调查区内采用典型取样法设置样地,选择群落结构完整、层次分明、生境条件相近的地段作为固定调查样地。根据黄连木在南太行山区间断性片状分布特性,随机在黄连木分布区选取12个具有代表性的样地。用罗盘仪进行样地的布设,在样地四角树立标志,大于5°的坡度进行坡度改正,要求测量闭合差不大于1/250,样地水平投影面积为20 m×30 m。调查前,用塑料绳把样地分割成6个10 m×10 m的调查小样方,并进行编号,以小样方为调查单元进行调查。样地基本概况见表1。
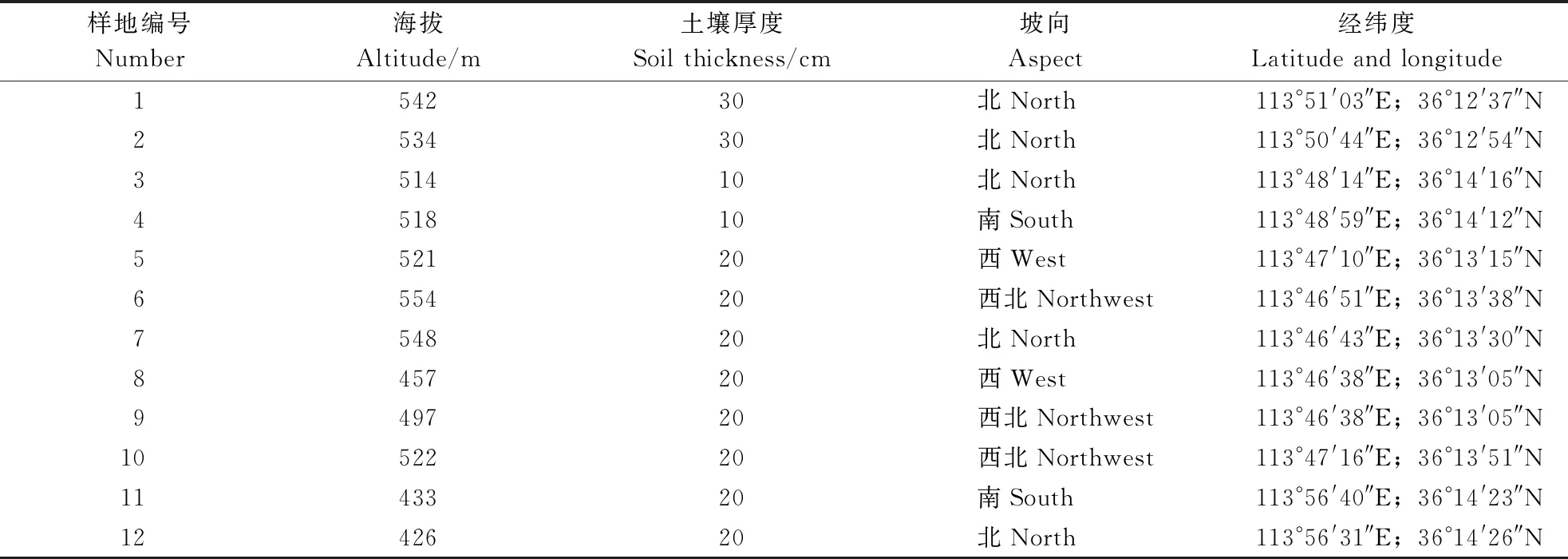
表1 样地基本概况
2.2 研究方法
2.2.1 样地调查 对每个20 m×30 m典型标准地中的乔木树种进行每木检尺(起测径级为5 cm),记录各树种名称、胸径、树高、枝下高、冠幅(东西-南北)等。以每个20 m×30 m的典型调查样地内布设的6个10 m×10 m的小样方为基础,作为灌木调查样方,整个调查区域内共调查72个灌木样方,记录样方中全部的灌木树种、株数、盖度等基本信息,对于达不到起测径级的乔木,作为灌木处理。其中,固定样地中的林下黄连木幼树通过直接观察或无损伤刨挖根颈部的方式确定其繁殖方式,分别记录实生幼树与萌生幼树株数。记录样方中黄连木株数时,高度180 cm以上长势良好的幼树按1株记数,高度180 cm以下的幼树以及生长不健康的植株按半株记数,萌生一丛按1株记数。在每个10 m×10 m的典型调查样地内各设置4个1 m×1 m草本小样方,每个样地共调查24个草本小样方,整个调查区域内共调查288个草本小样方,记录草本植物的名称等基本信息。
2.2.2 群落各层次物种重要值计算 样地中各层次物种重要值计算参考方精云等[18-20]的计算方法:乔木树种的重要值=(相对多度+相对显著度+相对频度)/3;灌木树种的重要值=(相对高度+相对频度+相对盖度)/3;草本植物的重要值=(相对盖度+相对频度)/2。
2.2.4 样地群落中黄连木幼树更新方式分析 按照黄连木幼树的生长情况,把黄连木幼树的高度划分为6级。其中,高度30 cm以下的划为Ⅰ级苗;高度30~80 cm划为Ⅱ级苗;高度80~130 cm划为Ⅲ级苗;高度130~180 cm划为Ⅳ级苗;高度180~230 cm划为Ⅴ级苗;高度230 cm以上的划为Ⅵ级苗。群落中的黄连木幼树高度转化率计算方法为相邻苗级间,大一苗级幼树株数与小一苗级株数之比,记为Ⅱ/Ⅰ、Ⅲ/Ⅱ、Ⅳ/Ⅲ、Ⅴ/Ⅳ、Ⅵ/Ⅴ。
利用SPSS 17.0软件对不同样地、不同更新方式下黄连木的株数与苗级进行方差分析及多重比较。结果表示为平均数±标准误。
3 结果与分析
3.1 样地基本数据分析
根据调查结果(表2),12个样地中,黄连木纯林占7个,黄连木混交林占5个,典型群落为黄连木群系。黄连木纯林与以黄连木为优势种的混交林的林龄在20~50 a之间。以20 a为1个龄级,10~20 a林龄的幼龄林占调查总样地的16.67%,21~40 a林龄的中龄林占58.33%,40 a以上的近熟林占25.00%。

表2 样地林分概况
3.2 样地物种多样性分析
3.2.1 不同垂直结构物种重要值分析 由表3可知,在样地群落中,乔木层共有5科6属6种,其中,黄连木的重要值为49.919 0,高于其他乔木,且黄连木幼树的重要值也高达30.324 2,是该调查区的优势树种,在林分中占据主导地位;次优的为栓皮栎和山槐,重要值分别为36.074 8和4.494 8,均远低于黄连木;其他乔木的重要值之和仅为9.511 4,在样地群落中表现为零星生长。样地群落的灌木层共有11科15属15种,较乔木层种类丰富,其中,重要值大于10的黄荆(24.014 8),为该调查区群落中主要灌木组成树种。样地群落中,草本层植物比乔木层与灌木层种类更加丰富,调查区内共记录到26种草本植物,分属15科24属,重要值大于10的草本植物有黄背草(16.053 7)和艾(14.456 9),是组成该调查区群落草本层的主要植物。

表3 黄连木群落不同垂直结构优势物种重要值
3.2.2 群落物种多样性分析 由表4可知,调查区的物种丰富度(S)、Simpson多样性指数(D)均为乔木层<灌木层<草本层,说明林下灌木层与草本层的植物种类较乔木层丰富,且草本层植物复杂程度最高,主要是太行山区黄连木群落郁闭度较小(少量郁闭度大于0.7,大部分郁闭度为0.4),林下光照充足,有利于灌木层和草本层植物的生长。乔木层的Simpson优势度指数(C)最大,为0.467 3,在调查区形成了以黄连木为主要优势树种的群落类型。
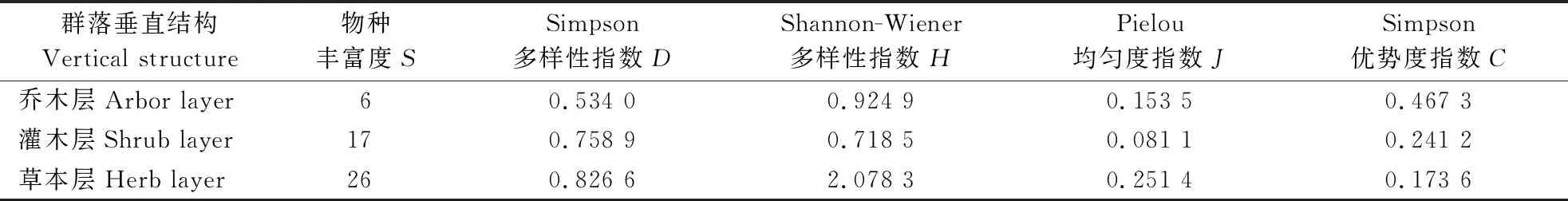
表4 黄连木群落垂直结构物种多样性分析
3.2.3 不同坡向群落垂直结构的多样性特征 黄连木在自然环境下具有一定的自我更新能力,但是在不同的立地条件下,调查区内不同坡向的物种多样性指数的变化不同。由表5可知,乔木层中,阴坡的物种丰富度(S)、Simpson 多样性指数(D)、Pielou 均匀度指数(J)较大,其次为阳坡,半阴坡最小。灌木层的物种丰富度(S)、Simpson 多样性指数(D)与乔木层的变化相同,为阴坡>阳坡>半阴坡。可能是乔木层和灌木层的阴坡土层相对较厚,有利于植物的生长,阳坡土层虽然瘠薄,但生长的树种以阳性树种为主,半阴坡不适宜黄连木等阳性树种的生长。草本层的物种丰富度(S)与乔木层、灌木层不同,表现为半阴坡>阴坡>阳坡,Simpson 多样性指数(D)表现为阴坡>半阴坡>阳坡,应是是阴坡土层相对较厚,有利于草本植物的生长。灌木层和草本层的Simpson 多样性指数(D)和Shannon-Wiener 多样性指数(H)均比乔木层高。
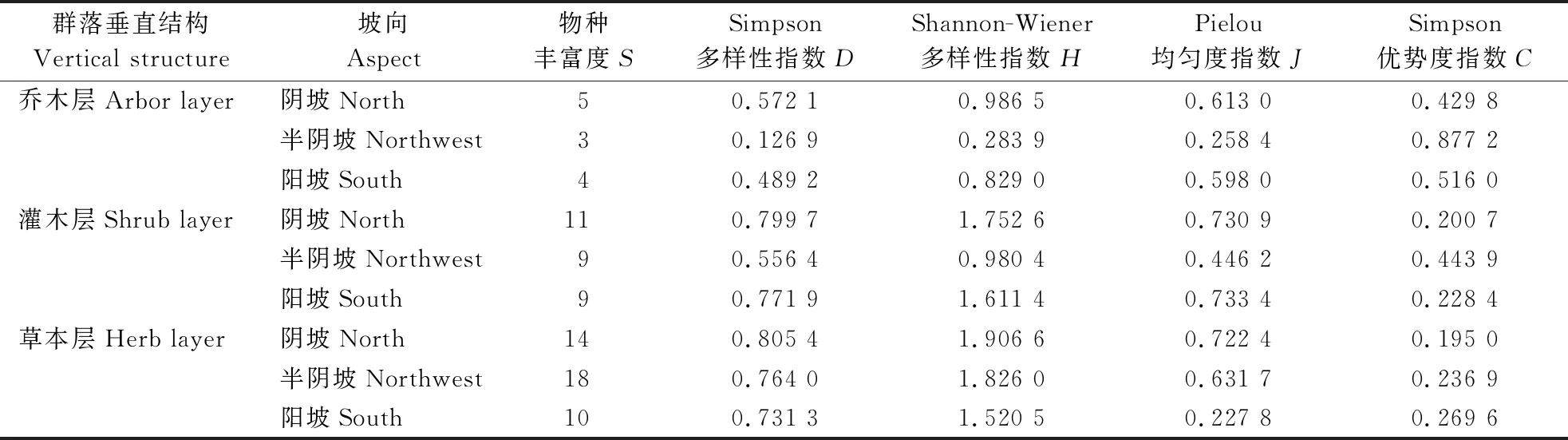
表5 黄连木群落不同坡向群落垂直结构多样性特征
3.3 黄连木不同更新方式与幼树苗级转化率分析
3.3.1 黄连木群落幼树更新方式变化分析 由图1、表6可知,黄连木的天然更新以萌生为主,12块调查样地中,黄连木的萌生幼树明显多于实生幼树,且大部分样地的萌生幼树株数达到实生幼树的2倍以上,萌生幼树与实生幼树间存在极显著差异(DF=11,P=0.000 2),这可能是黄连木适应于南太行山区自然环境的结果。

图1 不同样地黄连木萌生幼树与实生幼树株数对比

表6 黄连木幼树不同更新方式的差异显著性比较
3.3.2 更新方式对黄连木幼树苗级数量分配的影响 通过对黄连木群落内黄连木幼树的统计和高度分级,把黄连木幼树的高度分为Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ级。调查分析发现,萌生的黄连木幼树不同苗级的数量分配均大于实生的,且两种更新方式的黄连木幼树苗级数量分配均在Ⅱ级达到最高。不同苗级间,黄连木萌生幼树Ⅰ、Ⅱ、Ⅲ级的数量分配达到显著水平,Ⅳ、Ⅴ、Ⅵ级的数量分配不显著;在实生幼树中,仅有Ⅰ、Ⅱ级的数量分配存在极显著差异,其他苗级的数量分配不显著(表7)。

表7 不同更新方式下幼树苗级数量分配与多重比较
3.3.3 更新方式对黄连木幼树高度转化率的影响 两种更新方式下,黄连木幼树各苗级高度转化率影响主要表现在Ⅱ级苗与Ⅲ级苗之间,其他4组苗级高度转化率没有明显差异,萌生幼树不同苗级的高度转化率均高于实生幼树。随着苗级转化升高,不同更新方式对黄连木幼树高度转化率的影响逐渐减小。在萌生更新方式下,Ⅱ级苗向Ⅲ级苗转化的高度转化率与Ⅲ级苗向Ⅳ级苗转化的高度转化率差异不显著外,其他3组各苗级间高度转化率与Ⅱ级苗向Ⅲ级苗转化的高度转化率、Ⅲ级苗向Ⅳ级苗转化的高度转化率存在极显著差异;在实生更新方式下,Ⅰ级苗向Ⅱ级苗转化的高度转化率与Ⅱ级苗向Ⅲ级苗转化的高度转化率差异不显著,其他3组各苗级间高度转化率与Ⅰ级苗向Ⅱ级苗转化的高度转化率、Ⅱ级苗向Ⅲ级苗转化的高度转化率存在极显著差异(表8)。

表8 不同更新方式下幼树高度转化率及多重比较
4 讨论与结论
重要值是评估物种在植物群落中地位和作用的综合指标,其数值大小可作为衡量群落中物种相对重要性的一个重要度量指标[25]。对南太行山区样地群落乔木层、灌木层、草本层3个群落垂直结构的物种重要值进行分析发现,调查样地形成了以黄连木为主要单优势树种的黄连木群落。调查区内群落演替结果说明,南太行山区的生态环境条件不太适合大多数树种生存而比较适于具有耐干旱、瘠薄和适应性强等特点的黄连木生存。
物种多样性是衡量一定生境范围内物种资源丰富程度的重要客观指标,反映了植物群落中物种的丰富度、均匀度等指标变化,可以表征森林生态系统中群落的特征及其演化规律[26]。文中对南太行山区黄连木群落乔木层、灌木层、草本层的物种多样性进行分析发现,林下灌木层与草本层植物的种类较乔木层丰富,且调查样地中灌木层和草本层的Simpson多样性指数(D)越大,林下更新情况越弱,这与高晓琳[27]的研究结果一致。而乔木层的优势度指数(C)最高,为0.467 3,在调查区形成以黄连木为主要优势树种的群落类型,且大部分样地为黄连木纯林,这与李献生等[28]对河南省黄连木资源分布调查结果中豫北太行山黄连木集中分布区通常为纯林结果一致,且代表性群落一致。从不同坡向来看,灌木层的物种丰富度(S)、Simpson多样性指数(D)与乔木层相同,均为阴坡>阳坡>半阴坡,草本层的物种丰富度(S)则为半阴坡>阴坡>阳坡,Simpson 多样性指数(D)为阴坡>半阴坡>阳坡,这可能与调查区内阴坡土壤较阳坡肥沃有关。而不同坡向(阴坡、半阴坡、阳坡)的乔木层优势度均高于灌木层和草本层,且表现为半阴坡>阳坡>阴坡,这可能与黄连木为阳性树种有关。
黄连木具有较强的萌芽更新能力,黄连木群落在不同的样地中,各苗级间存在显著差异,且黄连木的萌生幼树株数均高于实生幼树,这一结果与张宏文等[15]、高晓琳[27]的研究结果相同,与黄连木的更新策略有关。另外,实生幼树的生长是从种子开始的,种子从生产到长成幼苗再到幼树的更新是植物生活史中最为敏感且脆弱的阶段,而动物取食种子[29]、种子受到病原体侵袭引起霉变与幼苗叶片脱落[30]、林下凋落物影响种子接触土壤减少种子萌发与定居的可能性[31]等各种生物或非生物制约因素[32],都影响种子的萌发和实生苗生长,而南太行山天然的陡峭地势可能减少了动物取食活动与林下凋落物的堆积,使得该调查区实生幼树株数也占据一定比例,也在一定程度上说明了太行山区适合黄连木生长繁殖。
黄连木幼树各苗级的数量分配在Ⅱ级达到最高,高度转化率主要表现在Ⅱ级苗与Ⅲ级苗之间,说明在两种不同更新方式下,黄连木幼树生长均在Ⅱ级以后开始趋于稳定。苗级在Ⅳ级以上的黄连木幼树,数量较少,出现了苗级缺失现象,不同更新方式对黄连木幼树的高度分布作用的影响也随之减少,这种苗级缺失现象在朱学灵等[33]对栎林群落幼苗库和张亚芳等[34]对杜松种群的研究结果中也有相似表现。另一方面,实生更新在黄连木的自我更新中也占据一定的比例。黄连木在天然林下的这种双重更新方式可能与其本身可以在生境砾石含量高,植被破坏严重,生态条件恶劣的环境中表现出很强生长优势的生理特性有关,能成为南太行山区的主要优势树种。
综上所述,黄连木的天然更新除了受到自身生物学特性影响,可以适应恶劣的自然环境外,还受到包括坡向、局部小气候、林下生物多样性等生境多种因素的综合影响。另外,黄连木幼树更新过程中可能受到动物取食、病原体侵袭、林下凋落物等影响种子和幼苗死亡机制的因素制约,出现了萌生幼树多于实生幼树的更新现象。本研究对集中分布区黄连木群落的生物多样性及该区黄连木群落的不同更新方式下不同苗级更新进行分析,这有助于进一步了解黄连木种群的维持和更新机制,探究该地区黄连木群落的更新演替过程,为黄连木群落的保育与可持续利用提供理论依据。在今后对南太行山区黄连木群落的更新保护工作中,可以通过采取适当更新措施进行人工干预,促进种子萌发,科学管理,保护萌生幼树生长,使黄连木的双重更新策略发挥更大作用。
