白令海及西北冰洋有机质来源与新鲜程度的脂肪酸指示
2020-11-16李科金海燕赵香爱庄燕培季仲强张扬陈建芳
李科,金海燕,2,赵香爱,庄燕培,季仲强,张扬,陈建芳,2
( 1. 自然资源部第二海洋研究所 自然资源部海洋生态系统动力学重点实验室,浙江 杭州 310012;2. 卫星海洋环境动力学国家重点实验室,浙江 杭州 310012)
1 引言
过去几十年,北冰洋是全球变化最显著的海区之一,大气及海表升温、海冰消融是其主要特征[1–4]。伴随着升温与海冰消融,北冰洋的物理化学生物性质正发生一系列强烈且持续的变化;预计在将来几十年,北冰洋还将以同样、甚至更强烈的速率发生变化[5–9]。
“生物泵”是指海洋中的初级生产者通过光合作用将大气CO2固定为颗粒有机碳,沉降至海底,埋藏在沉积物中的过程,具有降低大气CO2的能力。北冰洋的“生物泵”如何响应北极快速变化,是全球碳循环研究中的一个关键问题[10–12]。
北冰洋具有全球最为宽广的大陆架,在大陆架埋藏的有机质占全北冰洋的70%以上[13]。其中,西北冰洋的楚科奇海陆架由于生产力高,生物粒径大,水温低等特点,“生物泵”效率远高于其他区域[14–15]。未来,开阔水域增加[16]与太平洋入流增强[17]导致的生产力升高可能会促进“生物泵”的运转;但浮游植物小型化[18]、水温升高[19]、浮游食物链增强[20]等又能影响“生物泵”结构,降低其效率。为了更好预测未来“生物泵”的变化,需加深对现代“生物泵”的理解。
沉积物中生源指标为理解有机质的生产、沉降与保存提供了重要信息[21–22]。白令海与西北冰洋已开展大量生源指标工作,包括有机碳(Corg)[23]、生物硅(BSi)[24–25]、绿 素(chlorin)[26]、类 脂 生 物 标 志 物[27–28]等。前人工作表明,西北冰洋陆架是非常高效的有机碳汇区,而且沉降有机碳以海洋自生来源为主,主要由较大粒径的硅藻(>20 μm)贡献[29–30],其他藻类有机质所占的比例较小。这些海源自生有机质通过硅藻壳体的“压舱”效应,快速地沉降到海底,不仅形成了高效的“生物泵”,还支撑了大量底栖生物[31]。但是,前人工作主要运用了化学性质较为保守的生源指标,虽然指示了有机质的来源,但未反映有机质新鲜程度。沉降有机质新鲜程度也是“生物泵”结构的重要特征:新鲜有机质能够支撑起更大生物量,同时对于环境的变化也更加敏感。
脂肪酸作为沉积物中含量最丰富的脂类生物标志物之一,广泛应用于有机质的来源与保存研究中[32–34]。一方面,沉积物中脂肪酸的组成,可为有机质的来源提供线索[32]。不同来源的脂肪酸,其碳链长度及构型、双键的数目及位置存在差异。特定物种还有合成特征脂肪酸的能力[32,35]:海洋浮游植物的脂肪酸以短链偶碳数脂肪酸(12:n~20:n)为主,硅藻的特征脂肪酸为16:1ω9,绿藻主要以18:1、18:2为特征脂肪酸;陆地高等植物主要为长链偶碳饱和脂肪酸(22:0~30:0);而细菌则以支链及奇数碳链脂肪酸为主。另一方面,脂肪酸,特别是短链饱和脂肪酸及不饱和脂肪酸,也是十分易于降解的有机质,脂肪酸含量的多少,可以在一定程度上代表沉积物中有机质的新鲜程度,从而为“生物泵”结构提供指示意义[10,33]。
西北冰洋陆架区表层沉积物的脂肪酸研究较为有限,仅在十几年前有加拿大的研究者对楚科奇海的两个站位的沉积物柱样进行过脂肪酸分析[33,36]。结果表明,西北冰洋陆架区表层沉积物脂肪酸含量丰富,高于北冰洋其他海区。但以上工作的样本量过小,在缺乏代表性的同时,也难以反映脂肪酸的空间分布差异与影响因素。
本文依据白令海与西北冰洋表层沉积物中脂肪酸的分布与组成,结合主成分分析(Principal Components Analysis,PCA)的统计手段,对楚科奇海以及白令海有机质的来源与新鲜程度进行了分析,并尝试探讨北极快速变化对以上区域“生物泵”可能造成的影响。
2 采样站位及研究方法
2.1 采样站位及方法
本研究所分析的表层沉积物样品依托第五次、第六次中国北极科学考察航次采集,共33个站位,分布于白令海盆、楚科奇海盆、加拿大海盆(图1),采样时间分别为2012年夏季与2014年夏季。表层沉积物用箱式采样器获取,取表面0~1 cm。沉积物在采集后保存于–20°C冷库,直至进一步处理。
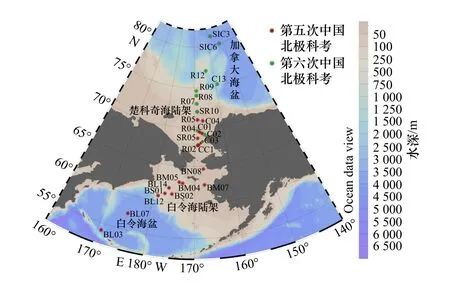
图1 采样站位Fig. 1 Sample Locations
2.2 脂肪酸化学处理及气相色谱分析方法
样品前处理及仪器分析测试工作于自然资源部海洋生态系统动力学重点实验室进行。沉积物在遮光、低温(–45°C)环境下真空干燥,而后在1个月内完成前处理及上机测试。
脂肪酸前处理方法运用Budge等[37]对Folch等[38]进行改良后的方法:取2 g干燥、研磨后的沉积物,加入内标(十九烷酸(19∶0),NU-CHEK)后,先以5 mL甲醇(CH3OH)超声萃取(200 W超声功率,5 min;3000 r/min,10 min),再用5 mL二氯甲烷/甲醇(2∶1,V/V)超声萃取两遍。将萃出液在低温水浴下氮吹浓缩,加铜粒去硫后,加入4 mL KOH / CH3OH(0.5 mol/L)溶液,在90°C下皂化2 h。用正己烷萃取出中性组分后,在剩余液体中加入10%盐酸至pH < 1,继续用正己烷萃取出酸性组分(主要为脂肪酸)。将酸性组分在低温下氮吹至1 mL,加入3 mL BF3/ CH3OH(15%)溶液,80°C下甲酯化1 h。冷却至室温后用5 mL正己烷萃出,低温氮吹,定容至0.5 mL,使用气相色谱仪器进行分析。溶剂皆为液相色谱纯,超纯水及盐酸亦皆以二氯甲烷萃取两遍后使用。
使用气相色谱−质谱检测仪(GC-MS,Agilent 7890A/5970C)鉴定化合物组成。色谱柱为HP-5毛细管柱(Agilent,30 m×0.32 mm×0.25 μm),升温程序为:初 始50°C,以5°C/min升 温 至270°C后 保 持5 min。以不分流模式进样,进样口温度250°C,载气为氦气,流速1 mL/min。质谱仪使用电子轰击源(70 eV)作为离子源,采用全扫描/选择离子检测采集模式。与标准样品(40种脂肪酸甲酯混标,安谱,N1070012)的出峰时间与特征离子谱图进行比对,确定物质种类。
定量分析使用气相色谱−火焰离子检测器(GCFID,Agilent 7890A),色谱柱为脂肪酸甲酯专用毛细管柱(Agilent,100 m×0.25 mm×0.25 μm),升温程序为:初始50°C,保持15 min后以5°C/min升温至120°C,保持10 min后再以5°C/min升温至200°C,保持20 min后再以5°C/min升温至230°C,最后保持20 min,总运行时间101 min。以不分流模式进样,进样口温度250°C,载气为高纯氮气,流量1.4 mL/min。FID温度为280°C,氢气与空气流速分别为35与400 mL/min。最后使用内标−标准曲线法确定脂肪酸含量。
脂肪酸命名常用格式为(a/i)X∶YωZ(c/t)。其中,X为总碳数,后面跟一个冒号;Y表示双键数;ω表示从甲基末端开始排序;Z是双键距离甲基端的距离;c 表示顺式异构,t 表示反式异构;a 和i 分别表示支链的反异构和异构。
2.3 主成分分析
由于脂肪酸种类众多,为寻求控制脂肪酸分布的主要因素,使用SPSS软件对33个站位中的26个脂肪酸指标进行主成分分析,具体分析方法详见文献[33–34, 39]。
3 结果
如表1与图2所示,总脂肪酸(Total Fatty Acid,TFA;以沉积物干重计)在33个站位的含量范围在13.70 μg/g至213.96 μg/g之间;南部楚科奇海陆架表层沉积物中的TFA非常高,平均值可达123.61 μg/g。所有站位中,对各类脂肪酸进行排序,短链饱和脂肪酸最高(Short Chain Saturated Fatty Acids,SCSFA,(42.3 ±25.43) μg/g),单不饱和脂肪酸次之(Mono-Unsaturated Fatty Acids,MUFA,(23.27±24.31) μg/g),长链饱和脂肪酸(Long Chain Saturated Fatty Acids,LCSFA)、奇碳数脂肪酸(Odd FA)、多不饱和脂肪酸(Poly-Unsaturated Fatty Acids,PUFA)较少,分别为(4.79±3.89) μg/g、(3.49±2.78) μg/g与(1.80±2.47) μg/g。
如表2所示,检出的26种脂肪酸中,16:0普遍含量最高;楚科奇海陆架区,16:1ω9与16:0含量相近,但在其他部分区域,14:0与18:0、18:1ω9取代16:1ω9成为除16:0外含量最高的脂肪酸。
4 讨论
4.1 白令海及西北冰洋表层沉积物中脂肪酸来源分析
海洋中脂肪酸的来源可分为海源、陆源以及细菌来源3个大类,大量研究表明,海源脂肪酸主要包括SCSFA、MUFA与PUFA,陆源脂肪酸主要为高等植物产生的LCSFA,Odd FA以及支链脂肪酸为细菌来源[32–34, 36]。
对33个站位的26种脂肪酸指标进行主成分分析,结果如表3所示:PC1至PC5的贡献率分别为45.00%、19.00%、9.19%、5.61%和4.54%,累计贡献率83.34%。将各脂肪酸与PC1、PC2的相关性作图(图3),脂肪酸主要分布在两个聚类,右下角的聚类主要包括海源、较易降解的SCSFA、MUFA与PUFA;另一个聚类以陆源、不易降解的LCSFA为主。该结果表明,LCSFA与SCSFA、MUFA、PUFA的分布由两组不同因素控制,与前人结论一致[33–34]。据此,将(SCSFA + MUFA + PUFA)定为海源脂肪酸,LCSFA定为陆源脂肪酸,Odd FA定为细菌脂肪酸(表4)。
对以上3种来源的贡献比例排序,所有站位均为海源 > 陆源 > 细菌(图4)。海源脂肪酸在TFA中占70%以上,特别是加拿大海盆,海源脂肪酸的比例达95%左右(图5a)。陆源脂肪酸比例较低,除BL07(25.53%)及CC6(26.00%)两站位以外,均不超过15%(图5b)。细菌脂肪酸占比最低,除BL14站(13.08%),均未高于8%;陆架区略高于陆坡及海盆区(图5c)。白令海陆架及西北冰洋陆架表层沉积物的细菌脂肪酸比例显著低于温暖海区,如东海、南海陆架细菌脂肪酸比例可达20%以上[41–42];这显示在白令海陆架及西北冰洋陆架,低水温抑制了细菌活动[43]。
此外,脂肪酸硅藻指数(以下简称硅藻指数,16:1ω9 / 16:0)可用来表征硅藻对有机质的贡献(表4,图5d)。16:1ω9是硅藻的特征脂肪酸,在大多数硅藻体内,16:1ω9常占TFA的40%以上;在沉积物或悬浮颗粒物中,较高的硅藻指数(16:1ω9 / 16:0 >1)指示硅藻在浮游植物群落中的主导地位[32]。硅藻指数分布范围为0~1.76,陆架区的硅藻指数普遍大于陆坡及海盆区,特别是楚科奇海陆架与白令海陆架边缘,硅藻指数大于1,指示硅藻占主要优势。
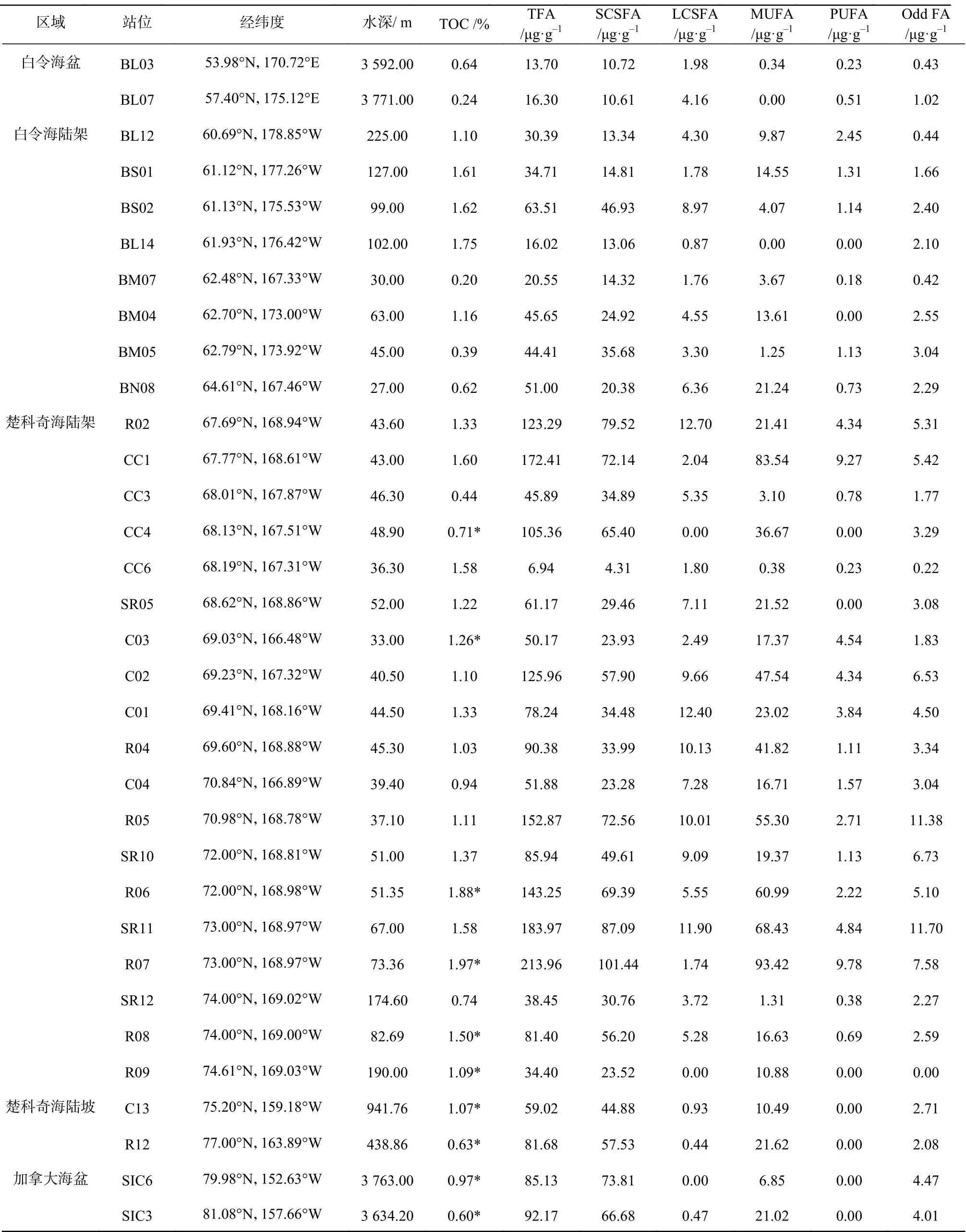
表1 白令海及楚科奇海表层沉积物站位信息及样品脂肪酸含量Table 1 FA, TOC and station properties of the Bering Sea and western Arctic Ocean surface sediments
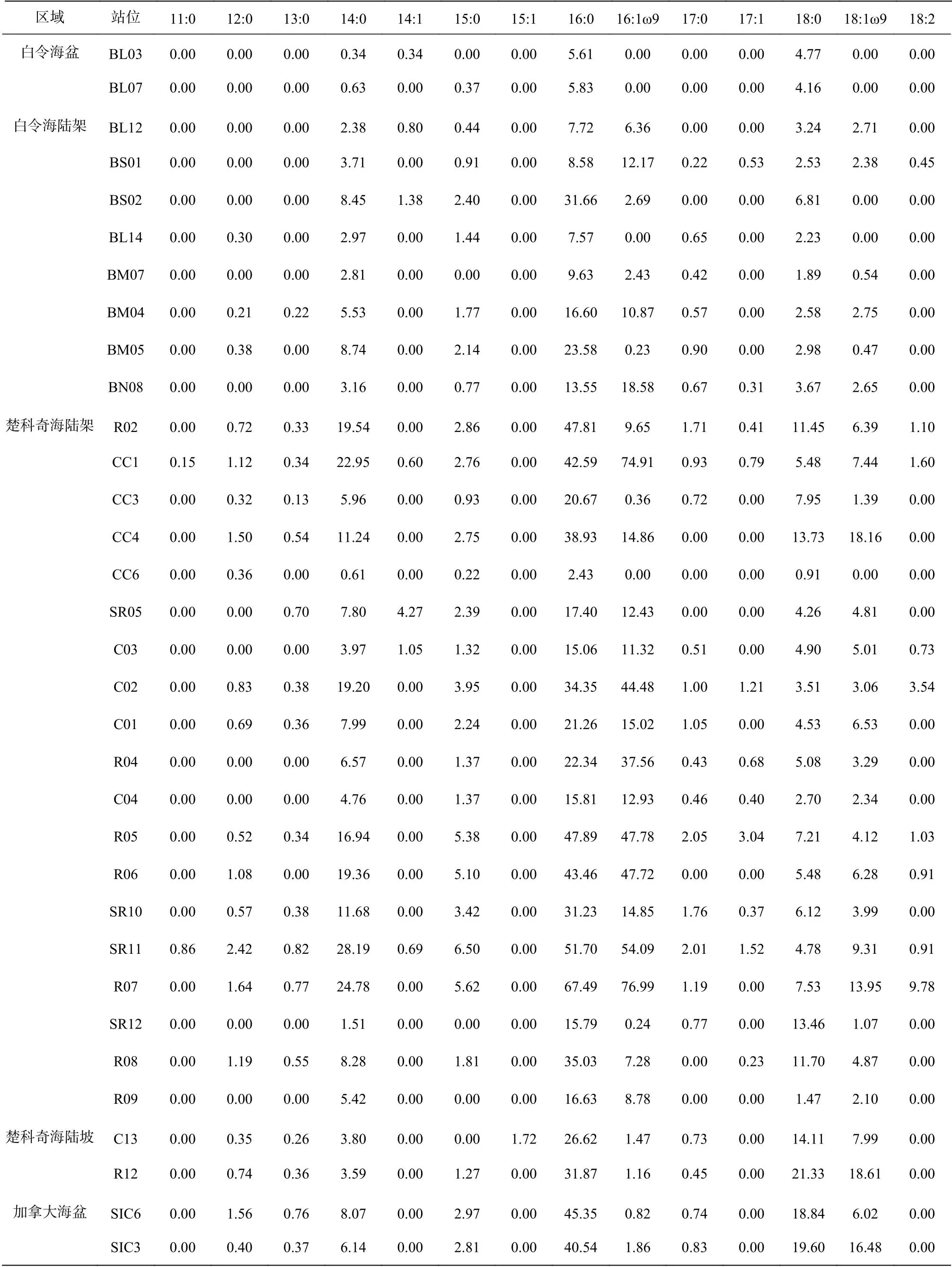
表2 白令海及楚科奇海各站位沉积物脂肪酸组成(单位:μg/g)Table 2 Individual FA compounds of the Bering Sea and the western Arctic Ocean surface sediments (unit: μg·g−1)
与本文其他海区相比,白令海盆区TFA最低((15.00 ± 1.30) μg/g),海源脂肪酸比例最低(75.33% ±7.13%),陆源脂肪酸最高(19.98% ± 5.56%),细菌脂肪酸较低(4.69% ± 1.57%),硅藻指数几乎为0。指示白令海盆区,海源有机质贡献较其他海域略少,可能与生产力较低[44]、水深较深有关;陆源有机质贡献较其他海域略高,与海源有机质比例低、且陆源脂肪酸不易降解有关[34];细菌脂肪酸含量低,指示细菌活动较弱,可能与低温以及新鲜有机质的缺乏有关[43]。
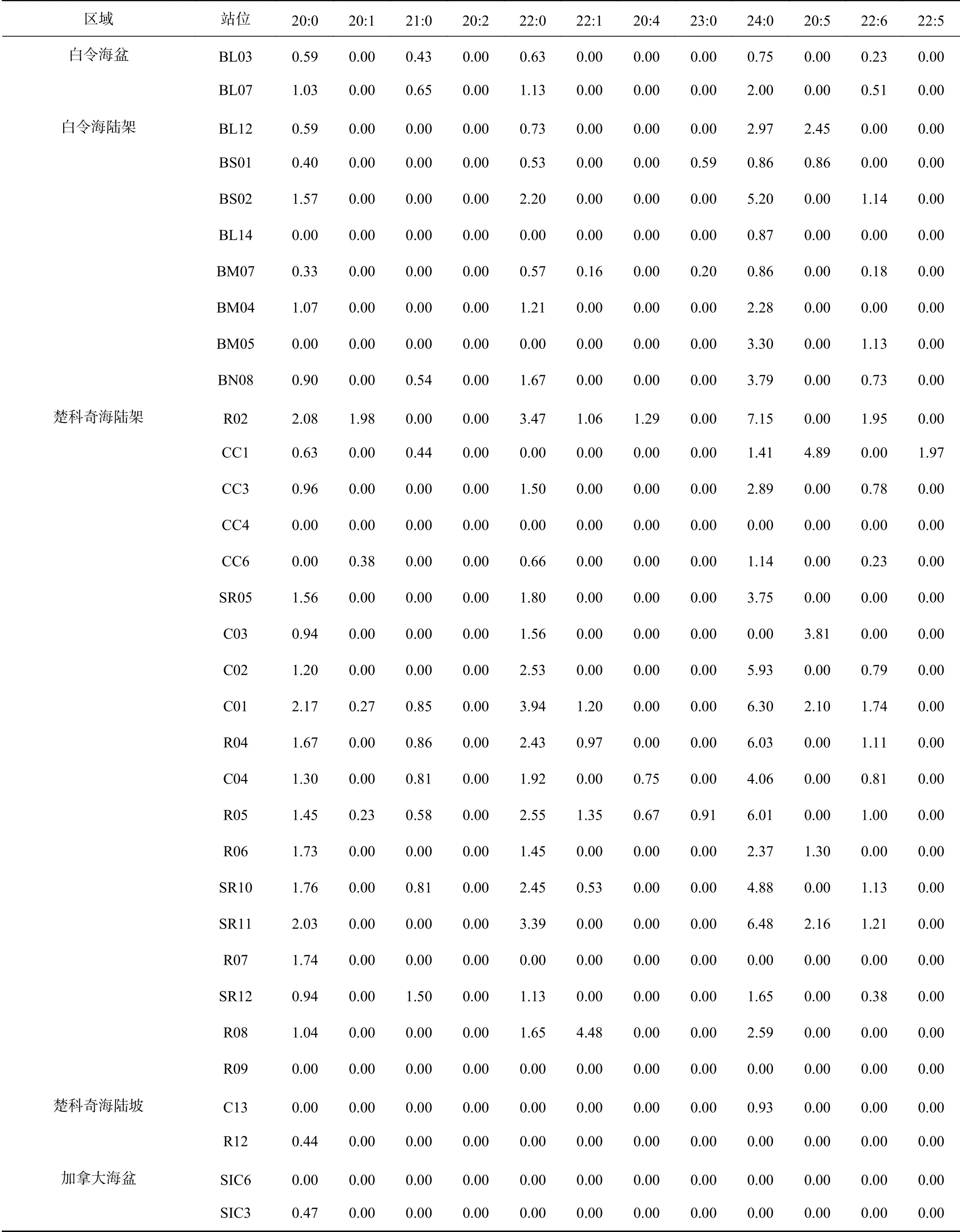
续表2
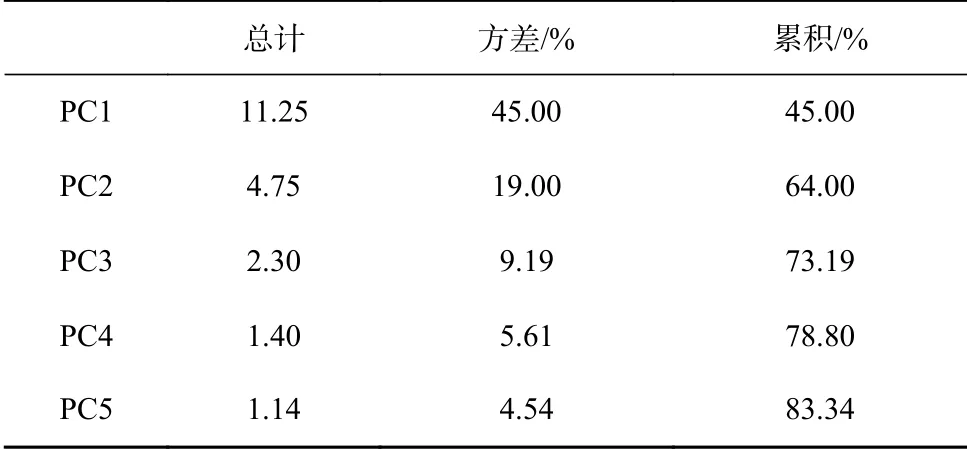
表3 主成分分析结果Table 3 PCA results
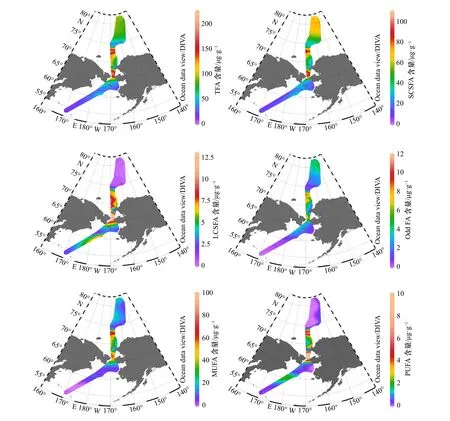
图2 白令海及西北冰洋表层沉积物中总脂肪酸及各类脂肪酸含量Fig. 2 TFA, SCSFA, LCSFA, Odd FA, MUFA and PUFA contents of the Bering Sea and the western Arctic Ocean surface sediments
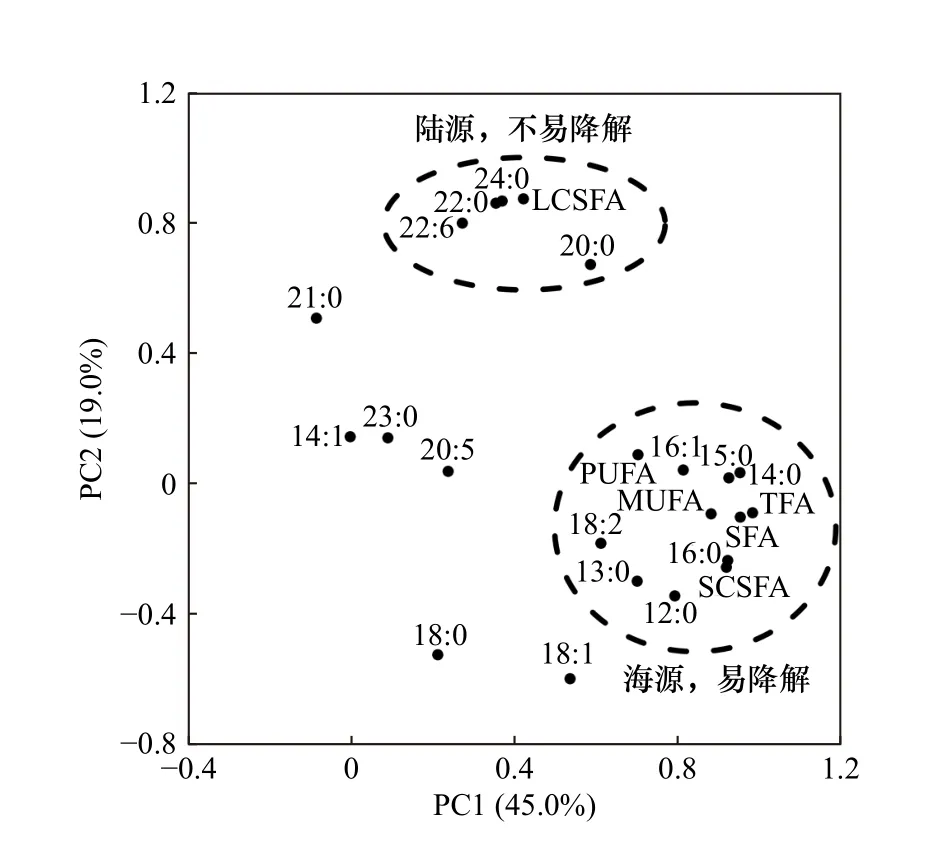
图3 各脂肪酸与PC1、PC2的相关性Fig. 3 First two principal components score plot of FAs
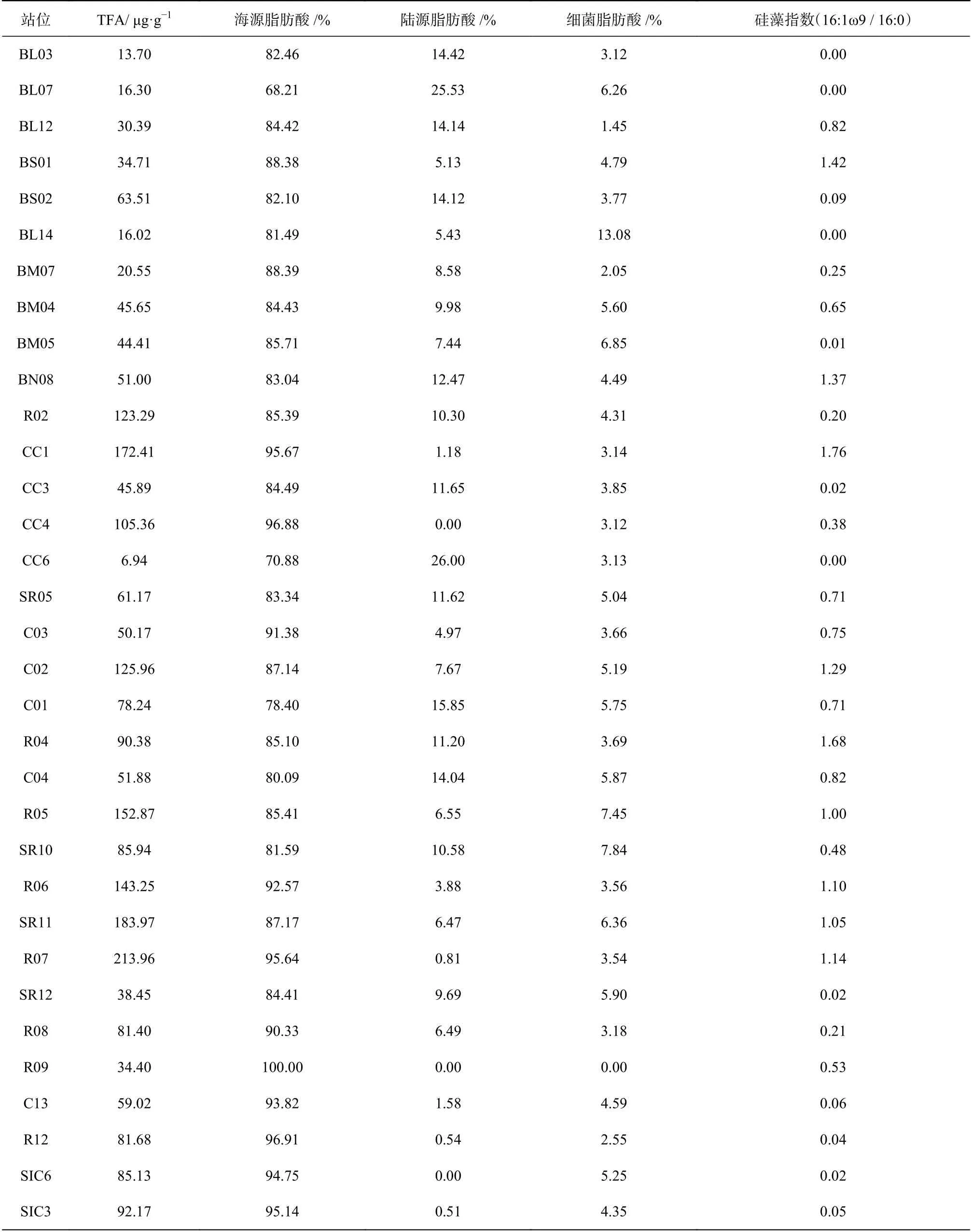
表4 各站位不同来源脂肪酸百分比及硅藻指数Table 4 Percentage of FA sources and diatom index of the Bering Sea and the western Arctic Ocean surface sediments
白令海陆架区,TFA((38.28 ± 14.89) μg/g)略高于白令海盆,海源脂肪酸含量较高(84.75% ± 2.45%),陆源脂肪酸含量低(9.66% ± 3.40%),细菌含量略高(5.26% ± 3.38%),硅藻指数在白令海陆架北缘与南缘较高(> 0.82),中部较低(< 0.65)。白令海陆架区陆源脂肪酸绝对含量与白令海海盆相当,但陆源脂肪酸的百分比要显著小于后者:这一方面是由于白令海陆架区生产力高于海盆区;另一方面,也因为白令海陆架区水深远小于海盆区,使得海盆区原本就贫瘠的海源脂肪酸在更长的沉降时间中进一步被分解,而陆源脂肪酸的化学性质则相对较稳定;两种因素叠加,陆架区的陆源脂肪酸比海盆区受到更多海源的稀释,从而比例更低。脂肪酸结果指示白令海陆架区有机质以海源自生为主,陆源贡献小;硅藻在陆架边缘区占主要优势;细菌脂肪酸与海源有机质的同步增加,可能指示细菌活动随新鲜有机质的增加而略为增强[43]。
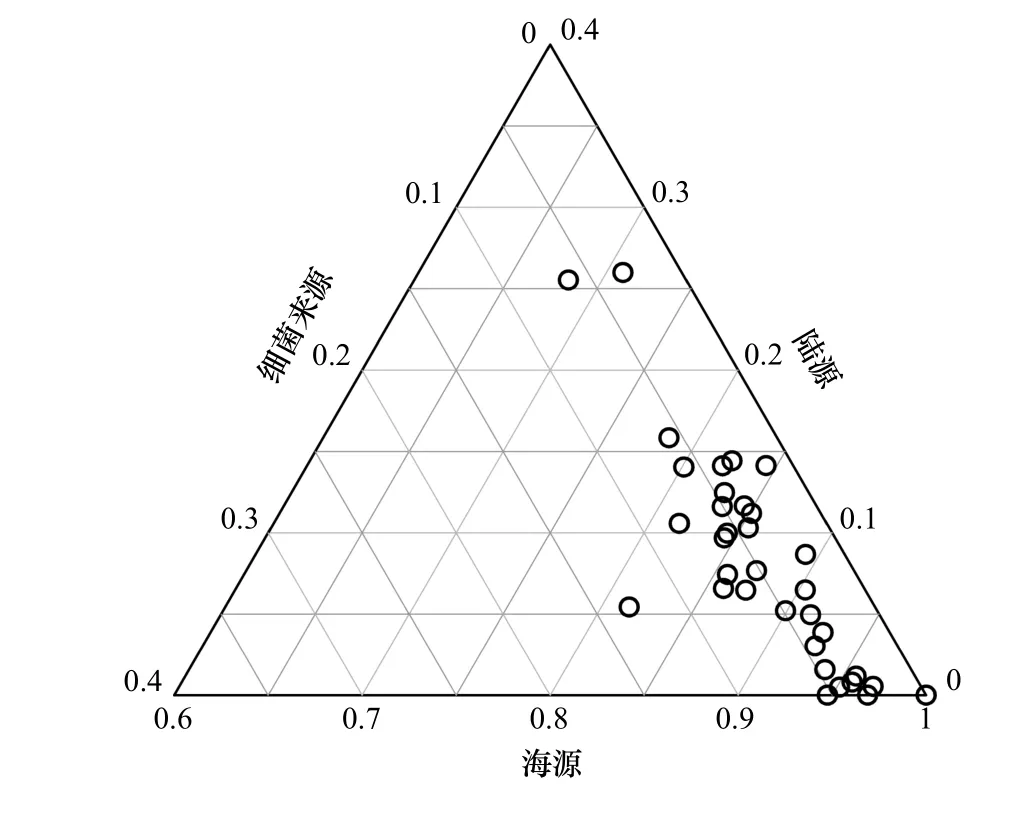
图4 脂肪酸来源三端元图Fig. 4 Ternary figure of FA sources
楚科奇海陆架,TFA非常高((97.15 ± 55.31) μg/g),特别是南部陆架,最高可达213.96 μg/g,与前人调查结果相同[33],比东海长江口区域高一个量级[41];海源脂肪酸占其中绝大部分(87.15% ± 6.97%),陆源脂肪酸与细菌脂肪酸比例均很低(8.37% ± 6.20%与4.45% ±1.79%),硅藻指数非常高。而且硅藻指数高值区与TFA高值区域重合,表明楚科奇海陆架的高脂肪酸含量主要与硅藻有关:一方面,充足的营养盐支撑了极高的硅藻生物量,且硅藻在浮游植物群落中占绝对优势,与现场水柱生产力、浮游植物群落调查结果具有相似性[29,45],也与沉积物中的其他生源指标重建结果相同[24–26];另一方面,硅藻在高营养盐环境下通常形成较大的个体(> 20 μm)[29],为有机质的快速沉降提供了“压舱物”,减少有机质在水柱中的分解,使表层生产的新鲜有机质快速地沉降到沉积物表面。但值得注意的一点是,在空间分布上,沉积物中脂肪酸与绿素[26]、生物硅[25]的高值区吻合得较好,但与表层生产力高值区[29,45]存在一定的错位,这很可能由于表层颗粒物在垂直沉降过程中同时也在发生着侧向输运,强劲的太平洋入流将颗粒物沉降的区域带离了生产区域。偏低的细菌脂肪酸比例亦表明,细菌活动在低温下受到抑制,有利于新鲜有机质的保存。
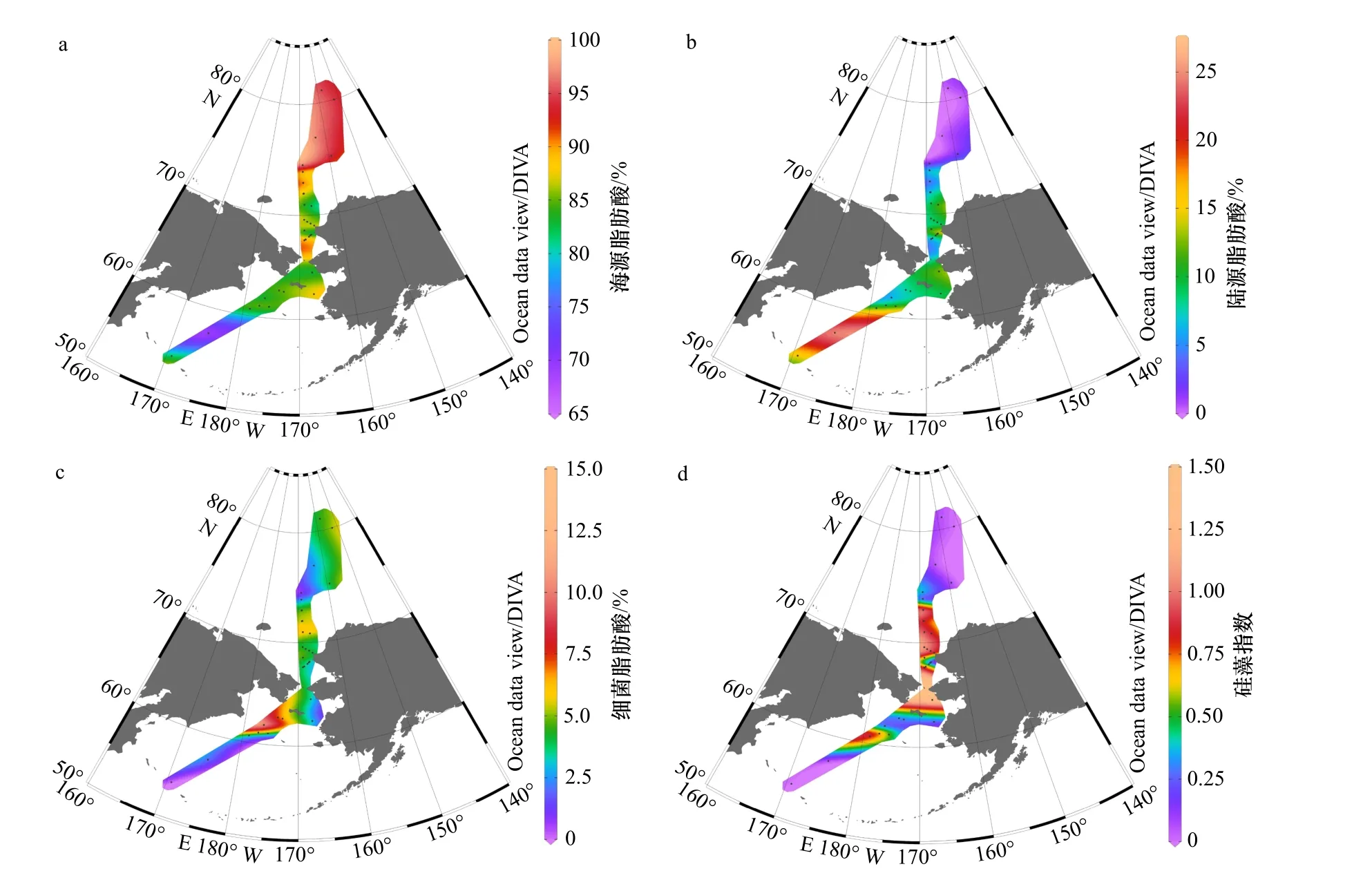
图5 各来源脂肪酸百分比及硅藻指数分布图Fig. 5 Percentage of FA sources and diatom index of the Bering Sea and the western Arctic Ocean
楚科奇陆坡、加拿大海盆与深度相似的白令海盆存在明显差异:TFA非常高((70.35 ± 11.32) μg/g与(88.65 ± 3.52) μg/g),海源脂肪酸几乎占全部(95.37% ±1.54%与94.95% ± 0.20%),陆源脂肪酸非常少(1.06% ±0.52%与0.25% ± 0.25%),细菌脂肪酸也较低(3.57% ±1.02%与4.80% ± 0.45%),硅藻指标几乎为0,18:0、18:1ω9明显高于其他海区。根据这些特点进行推论,首先,可以得到与Honjo等[46]相似的结论,即楚科奇陆坡、加拿大海盆由于海冰的存在,有机质沉降机制与白令海盆不同:持续的海冰覆盖、稀少的“压舱物”阻止了大部分陆源有机质在海盆与陆坡的沉积;其次,偏低的硅藻指数与较高的18:0、18:1ω9脂肪酸(表2)指示,硅藻在该海区不占主导,绿藻与金藻占浮游植物群落的主要部分[32,35],与该区域叶绿素极大层色素分析[47]以及沉积物碳酸钙含量[48]的证据一致。然而,Honjo等[46]认为海盆区的有机质来源可能主要由侧向输运贡献,垂向沉积较弱;冰藻在融冰期的快速沉降可能也贡献了较多的新鲜有机质。但以上观点在脂肪酸结果中无法反映,还需要更多证据。
脂肪酸结果显示白令海及西北冰洋表层沉积物中的有机质主要由海源自生来源产生,陆源贡献不显著,与表层沉积物中甘油二烷基甘油四醚(GDGT)对有机质来源的结果相似[49]。细菌脂肪酸比例偏低,表明细菌活动在低温下受到抑制[43]。楚科奇海陆架区TFA与硅藻指数均非常高,与其他生源指标类似,均指示海源有机质的大量沉积主要由高生物量的硅藻构成[4,25–26];硅藻壳体的“压舱物”效应、受抑制的细菌活动,也是新鲜有机质快速沉降、高效保存的原因。楚科奇陆坡、加拿大海盆的浮游植物群落由绿藻与金藻主导,硅藻不占主要优势。
4.2 脂肪酸所指示白令海及西北冰洋表层沉积物中有机质的新鲜程度
脂肪酸分析结果显示,楚科奇海陆架由于生产力高,硅藻壳体“压舱物”丰富、细菌活动受低温抑制等原因,是新鲜有机质的高效沉降区;而白令海陆架新鲜有机质沉降效率明显较低。为了比较新鲜有机物沉降的差异,图6总结了北冰洋几个特征陆架区表层沉积物中TFA与总有机碳(Total Organic Carbon,TOC)的关系[33,50]。需要说明的是,前人工作[33–34]将SCSFA、MUFA、PUFA定义为新鲜有机质的指标,但西伯利亚海陆架区的文献资料[50]只提供了TFA,未细分脂肪酸种类。需要先分析易降解脂肪酸(SCSFA +MUFA + PUFA,表征有机质新鲜程度)与TFA之间的相关关系。分析结果显示,本研究中33个表层沉积物中易降解脂肪酸与TFA呈显著正相关(图7),因而TFA在本研究区域可作为新鲜有机质的指标。

图6 北冰洋部分陆架海区表层沉积物中TFA与TOC的关系[33, 50]Fig. 6 Relationship between TFA and TOC among the Bering,Chukchi, Beaufort and Siberian shelves[33, 50]
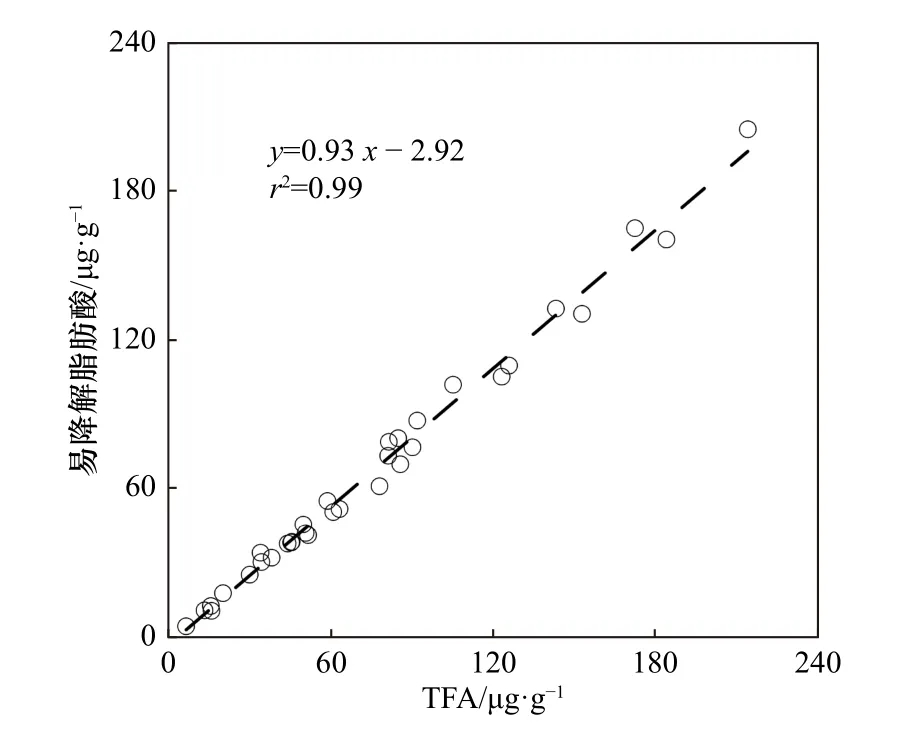
图7 本研究表层沉积物中易降解脂肪酸(SCSFA +MUFA + PUFA)与TFA的相关关系Fig. 7 Corrlation between labile FA (SCSFA + MUFA +PUFA) and TFA among all surface sediments in this study
由图6可见,楚科奇海陆架表层沉积物中的新鲜有机质占总有机质的比例明显高于白令海陆架与西伯利亚海陆架,3条拟合线的斜率分别是82.99,21.46与14.33,表示在相同的TOC含量下,楚科奇海陆架区新鲜有机质含量是白令海陆架的近4倍,是西伯利亚海陆架的近6倍。这是由于楚科奇海陆架区的生产力高,又有大粒径的硅藻壳体作为“压舱物”,因而有大量的新鲜有机质沉降到沉积物表面;同时,楚科奇海陆架区缺少陆源输入,对新鲜有机质的稀释效应弱,而西伯利亚海生产力较低,又有大量的陆源有机质通过河流输入,本就有限的海源新鲜有机质在陆源有机质的稀释作用下,占TOC的含量更低[50];白令海陆架虽然没有大量陆源物质输入,但生产力略低,且硅藻“压舱物”不如楚科奇海陆架充足[44],因而新鲜有机碳沉降速率不如后者。
楚科奇陆坡、加拿大海盆的有机质新鲜程度亦非常高,推测主要与有机质在融冰期快速沉降[51]与楚科奇陆架有机质的侧向输运[46]有关。但可惜脂肪酸解释能力有限且海盆区站位较少,无法深入讨论海冰与侧向输运对有机质沉降的影响。由于此区域是主要的海冰消退区,未来工作需更加注重对该区域的有机质沉降机制的研究。
4.3 陆架区有机质在北极快速变化下的敏感性
硅藻生物量大、细菌活动弱、新鲜有机质高效沉降的特点,使得楚科奇海陆架区“生物泵”在未来的北极快速变化下比北冰洋其他海区更为敏感。
由于太阳辐射量[7]与温暖太平洋水入流[17]的增加,楚科奇海陆架水将会升温[19],增强细菌活动,从而加快新鲜有机质的降解[43],将更多的二氧化碳释放回水柱中,降低“生物泵”的效率。目前,楚科奇海陆架区的高效碳沉降,除了生产力高、沉降快速等原因外,很大程度上依靠低温抑制的细菌活动,使得新鲜有机质在沉降过程中较少地受到降解。未来陆架水的升温,细菌活动的增强,很可能会影响有机碳的保存,降低“生物泵”的效率。
浮游植物的群落变化,也会影响有机质的沉降速率,从而影响“生物泵”的效率。目前楚科奇海的大量新鲜有机质沉降很大程度上得益于硅藻壳体的“压舱物”效应[24,44],若是由于温度、光照、营养盐等条件的变化,使硅藻在浮游植物群落中占比减小,硅藻壳体“压舱物”减少,在生产力不变的情况下,由于沉降速率的降低,造成有机质在水柱中停留、分解的时间增加,也会大大降低有机碳向沉积物输出的效率。
当然,最近国际上已有共识,北冰洋的浮游植物生物量在最近十几年来一直是增加的,预期未来的相当一段时间内也都会上升[16]。生产力的增加,是否能与“生物泵”效率的降低相抵消?楚科奇海,以及整个北冰洋的“生物泵”究竟如何变化?脂肪酸能够指征沉降颗粒物及沉积物中有机质的来源与新鲜程度,结合其他一系列指标,有望监测与研究未来北冰洋的“生物泵”变化。
5 结论
在白令海、西北冰洋各个海区中,楚科奇海陆架区表层沉积物中脂肪酸的含量非常高,特别是中部、南部陆架区,达到213.96 μg/g,是全球沉积物中脂肪酸含量最高的海区之一;白令海陆架的脂肪酸含量稍低于楚科奇海陆架;楚科奇海陆坡、加拿大海盆的浮游植物群落由绿藻与金藻主导。白令海陆架及西北冰洋表层沉积物中的脂肪酸指示了有机质主要由海源贡献,陆源、细菌来源都较低。楚科奇海陆架中南部、白令海陆架边缘硅藻指数高,有机质主要由硅藻贡献,其他海区硅藻不占主要优势。
楚科奇海陆架区由于生产力高、硅藻“压舱”效应明显、低温抑制细菌活动等原因,是新鲜有机质高效沉降的一个海区。这些新鲜有机质,在构成高效的“生物泵”,供养丰富的底栖生物的同时,也使得楚科奇海陆架区的有机质沉降对未来的北极快速变化较其他海区更为敏感:浮游植物群落变化、水温升高很可能会减少新鲜有机质的沉降量,降低“生物泵”的效率。未来楚科奇海陆架区乃至北冰洋的“生物泵”究竟会如何变化,还需进行长期、连续、全方位的监测与研究。脂肪酸作为有机质来源与新鲜程度的指标,有望应用于未来北冰洋“生物泵”变化研究中。
致谢:感谢中国第五次、第六次北极科学考察队为样品采集所付出的艰辛劳动;感谢自然资源部第二海洋研究所、自然资源部海洋生态系统动力学重点实验室各位老师的帮助;感谢中国极地研究中心及“雪龙”号给予的支持。
