小麦籽粒次生物质对麦红吸浆虫幼虫解毒酶活性及基因表达的影响
2020-11-16陈锐高贺张国军朱克岩成卫宁
陈锐,高贺,张国军,朱克岩,成卫宁
(1西北农林科技大学植物保护学院/农业农村部西北黄土高原作物有害生物综合治理重点实验室,中国陕西杨凌 712100;2Department of Entomology,Texas A&M University,Texas 77843,USA)
0 引言
【研究意义】麦红吸浆虫(Sitodiplosis mosellana)是小麦生产上间隙性猖獗成灾的重要害虫[1-2],以幼虫潜伏在颖壳内吸食正在灌浆的籽粒汁液,造成麦粒瘪疮、空壳或霉烂,一般使小麦减产10%—20%,重者30%—50%,甚至颗粒无收,严重影响小麦的产量和品质,甚至根本不能食用[3]。实践证明,选育和推广种植抗虫品种是降低吸浆虫危害最安全和经济有效的途径[4-5]。探究小麦灌浆期籽粒次生物质含量与小麦抗吸浆虫的关系,以及不同抗性品种次生物质含量对麦红吸浆虫幼虫解毒酶活性及其编码基因表达的影响,对深入研究小麦对吸浆虫的抗性机制及抗虫品种的选育具有重要意义。【前人研究进展】为了避免或减轻植食性昆虫的危害,植物与昆虫在长期的协同进化过程中形成了一系列防御机制,抗生性是植物抗虫的主要机制之一,植物的抗生性与次生物质含量关系密切,如高粱对西南玉米杆螟(Diatraea grandiosella)、小麦籽粒对玉米象(Sitophilus zeamais)、棉花对棉铃虫(Helicoverpa armigera)和绿盲蝽(Apolygus lucorum)的抗性分别与生腈糖苷、阿魏酸、单宁和棉酚等含量有密切关系[6-9]。为抵御次生物质的毒害,植食性昆虫也演化出了多种反防御机制。当取食不同寄主或不同类型次生物时,昆虫体内的解毒酶会变化,从而对摄入的有毒物质进行解毒和排毒[10]。谷胱甘肽-S-转移酶(glutathione S-transferase,GST)、羧酸酯酶(carboxylesterase,CarE)和细胞色素 P450 酶系(cytochrome P450,CYP450)是昆虫体内最重要的解毒酶,其活性和基因表达水平能够被多种次生物质诱导,以此增强对次生物质的代谢能力和对寄主的适应性[11-14]。研究表明,儿茶素和芦竹碱可以诱导麦长管蚜(Sitobion avenae)CarE和GST活性升高[15],单宁可以诱导杨小舟蛾(Micromelalopha troglodyte)GSTd1的表达[16];亚洲小车蝗(Oedaleous asiaticus)取食含单宁、黄酮、苯酚等次生物质含量高的抗性寄主冷蒿后,体内GST、CarE和P450活性及相关基因表达量显著升高[17]。【本研究切入点】多年来,国内外对小麦抗吸浆虫性进行了大量研究,发现不同品种(系)小麦对吸浆虫的抗性不同[18-20],抗性不同品种间次生物质含量亦存在差异[21-22],但小麦籽粒次生物质是否对吸浆虫解毒酶产生影响,解毒酶在吸浆虫对植物次生物质代谢及寄主适应性中所起作用尚不清楚。【拟解决的关键问题】分析不同抗性品种(系)小麦次生物质(单宁、总黄酮、阿魏酸、总酚)含量与小麦抗吸浆虫指标,以及与取食不同品种(系)的麦红吸浆虫幼虫3种解毒酶(GST、CarE和CYP450)活性及相关基因表达的关系,明确影响小麦抗麦红吸浆虫的关键次生物质及麦红吸浆虫对次生物质的解毒机制,为麦红吸浆虫综合治理和抗虫品种的培育提供新思路。
1 材料与方法
试验于 2015—2017年分别在陕西省西安市周至县和西北农林科技大学农业农村部西北黄土高原作物有害生物综合治理重点实验室完成。
1.1 供试小麦品种(系)
选取生育期相同或相近,抽穗期(敏感期)与麦红吸浆虫羽化高峰期吻合,前期鉴定连续多年对麦红吸浆虫抗性表现稳定的抗虫小麦品种(系)晋麦47、科农1006、陕麦139和陕农33,及感虫品种(系)小偃22、西农822、西农88和小偃6号[20]。所有品种(系)均来源于西北农林科技大学农学院。
1.2 田间试验设计
将供试材料于 2015年小麦适播期播种于麦红吸浆虫重发区陕西省西安市周至县试验地(N34°9′,E109°10′),每品种3个重复,每重复10行,行长1.5 m。翌年春季淘土调查虫口密度为390万头/667 m2,试验地常规管理,不施用任何农药,各品种(系)在田间自然感虫。
1.3 小麦品种(系)被害调查与试验样品收集
小麦灌浆期,每个品种(系)小麦各重复随机取50穗,带回室内逐穗、逐粒剥查并记载麦粒中的麦红吸浆虫幼虫数;同时收集各品种(系)健康籽粒,保存于-20℃,用于次生物质含量测定;收集足够多危害各品种(系)小麦籽粒的虫龄一致的幼虫,龄期依据虫体大小、颜色和Y型剑骨片有无区分[18],收集的幼虫每30头分装于2.0 mL冻存管,液氮速冻后-80℃保存,用于解毒酶活性和基因表达分析。调查结束后,计算每个品种(系)的平均穗被害率、粒被害率、单穗幼虫数和估计损失率。计算估计损失率时,麦粒中的幼虫数分为5级:0级:无虫/粒;1级:1头/粒;2级:2头/粒;3级:3头/粒;4级:≥4头/粒[20]。

1.4 小麦灌浆期籽粒次生物质含量测定
将收集的小麦籽粒80℃烘干至恒重,研磨成粉,过100目筛子后用于次生物质含量的测定,其中阿魏酸含量以阿魏酸为标品,采用比色皿法测定[23],单宁含量以儿茶素为标准品,采用香草醛法测定[6],总酚含量以没食子酸为标准品,采用Folin-Ciocalteu法测定[24],总黄酮含量以芦丁为标准品,采用亚硝酸钠-硝酸铝显色法测定[25]。每处理重复3次。
1.5 麦红吸浆虫幼虫GST、CarE和CYP450活性分析
1.5.1 酶液制备 从冰箱中取出各样品,称重后放入1.5 mL 离心管,分别加入 500 μL 0.04 mol·L-1pH 7.0、0.1 mol·L-1pH 6.5 和 0.1 mol·L-1pH 7.8 的磷酸缓冲液(含 1 mmol·L-1EDTA、1 mmol·L-1PMSF 和 1 mmol·L-1DTT),充分研磨后,匀浆液于4℃ 12 000×g下离心10 min,上清液分别为 CarE、GST 和 CYP450 酶液[26]。测试时,酶液再用缓冲液稀释成最佳取样浓度。每处理30头试虫,重复3次。
1.5.2 酶活力测定 3种解毒酶的活力均采用上海泛柯实业有限公司生产的酶联免疫分析试剂盒测定,具体操作步骤按试剂盒说明进行。
GST活力测定原理:用纯化的昆虫GST抗体包被微孔板,制成固相抗体,往包被单抗的微孔中依次加入 GST标准品和待测酶液,再与辣根过氧化物酶(HRP)标记的GST抗体结合,形成抗体-抗原-酶标抗体复合物,经彻底洗涤后加底物四甲基联苯胺(TMB)显色。用酶标仪在 450 nm波长下测定 OD值,通过标准曲线计算样品中GST活性。
CarE和CYP450活力测定原理与GST相同,区别仅在于以相应的酶标抗体进行包被。
以37℃下每毫克组织蛋白每分钟转化1 μmol底物(TMB)的酶量为一个活力单位(U/mg protein)。
1.5.3 酶蛋白浓度测定 以牛血清蛋白(BSA)为标准蛋白,采用考马斯亮蓝G-250染色法测定[27]。
1.6 麦红吸浆虫GST、CarE和CYP450表达水平分析
1.6.1 总RNA提取和cDNA合成 分别取危害各品种(系)小麦的麦红吸浆虫幼虫30头,按照TRNzol总RNA提取试剂盒(TIANGEN,北京)操作说明提取总 RNA,PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)反转录试剂盒(TaKaRa,大连)说明合成单链cDNA。重复3次。
1.6.2 荧光定量 PCR检测 在麦红吸浆虫幼虫转录组数据库搜索GST、CarE和CYP450酶编码基因,从中挑选表达量高的GST1、CarE2和CYP6A1Unigene序列设计定量引物(表 1)。以麦红吸浆虫GAPDH(GenBank登录号:KR733066)为内参,合成的cDNA为模板,按照 SuperReal PreMix Plus(SYBR Green)试剂盒说明书(TIANGEN,北京),在iQ5 Real-Time PCR Detection System(Bio-Rad,Hercules,CA)上进行 qPCR 分析。反应体系(20 μL):2×SuperReal PreMixPlus 10 μL,上下游引物(10 μmol·L-1)各 0.8 μL,cDNA 模板 1.0 μL,RNase Free H2O 7.4 μL。扩增程序:95℃预变性 15 min;95℃变性 30 s,55℃退火30 s,72℃延伸 30 s,40 个循环;最后 95℃ 30 s,55℃1 min,95℃ 15 s,进行熔解曲线分析,反应结束后读取Ct值。每样品进行3次技术重复。

表1 qPCR中应用的引物序列Table 1 Primer sequences for qPCR
1.7 数据分析
以取食陕麦139的麦红吸浆虫幼虫为基准,采用2-ΔΔCT法[28]计算危害各品种(系)小麦的麦红吸浆虫幼虫解毒酶基因相对表达量。小麦品种(系)间受害程度、次生物质含量、解毒酶活性和基因表达量进行单因素方差分析(P<0.05),Duncan氏新复极差法进行多重比较;Pearson法分析次生物质含量与小麦抗虫指标、解毒酶活性,以及基因表达量与酶活性间的相关性,所有分析在SPSS 19.0(Chicago,IL,美国)统计软件包上进行。
2 结果
2.1 不同品种(系)小麦受麦红吸浆虫危害程度
由表2可知,不同品种(系)小麦受害程度差异明显,其中前期鉴定为抗虫的4个品种晋麦47、科农1006、陕麦139和陕农33穗被害率、粒被害率、单穗虫口数和估计损失率分别为30.67%—69.67%、3.20%—7.11%、1.10—5.07头和 0.42%—2.43%,均显著低于4个感虫品种小偃22、西农822、西农88和小偃6号对应的 89.33%—95.33%、11.51%—15.15%、7.57—14.24头和 4.25%—6.12%(P<0.05)。抗虫品种和感虫品种内部受害程度也存在一定的差异,在抗虫品种中以陕麦139受害最轻,科农1006和晋麦47次之,陕农33受害较重;感虫品种中以小偃22受害最重,除穗被害率外,其他几个指标均显著高于西农 822、西农88和小偃6号。
2.2 不同品种(系)小麦籽粒次生物质含量比较
由表 3可知,不同小麦品种(系)灌浆期籽粒阿魏酸、单宁、总酚和总黄酮含量差异明显,其中阿魏酸和单宁含量在抗虫品种科农 1006中最高,分别为6.57和2.17 mg·g-1,显著高于所有感虫品种,在感虫品种西农822和小偃6号中最低,分别为4.61和1.02 mg·g-1,显著低于其他品种(系)。总酚含量在抗虫品种晋麦47中最高(24.86 mg·g-1),低抗品种陕农33中最低(12.56 mg·g-1)。总黄酮含量在感虫品种西农 88 中最高(3.93 mg·g-1),显著高于其他品种(系),小偃 6 号中最低(1.91 mg·g-1)。
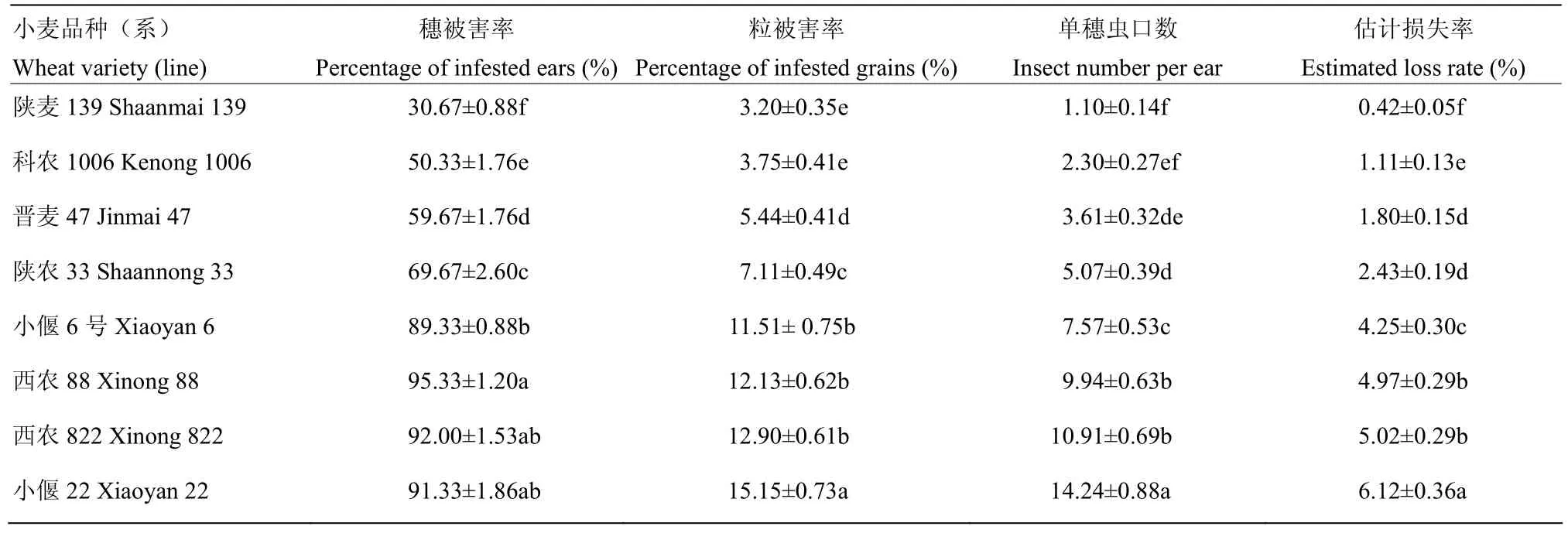
表2 麦红吸浆虫幼虫对8个品种(系)小麦的危害程度Table 2 Damage degree of eight wheat varieties (lines) caused by S. mosellana larvae
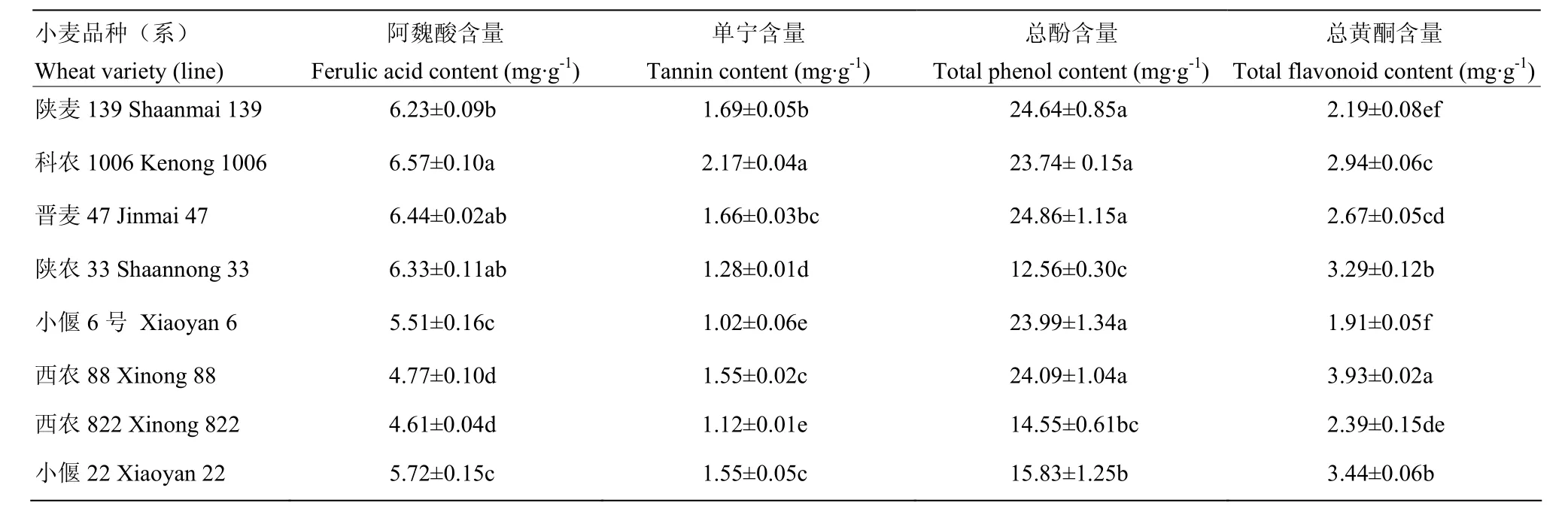
表3 8个品种(系)小麦籽粒次生物质含量Table 3 Contents of secondary metabolites in wheat kernels of eight varieties (lines)
2.3 小麦籽粒次生物质含量与抗虫性的关系
以抗、感程度不同的各品种(系)小麦穗被害率、粒被害率、单穗虫口数和估计损失率为抗虫指标,与小麦籽粒4种次生物质含量进行相关性分析,结果表明,阿魏酸含量与 4个抗虫指标均呈显著负相关(P<0.05),单宁、总酚和总黄酮含量与4个抗虫指标的相关性均未达显著水平(P>0.05)(表4)。
2.4 小麦品种(系)对麦红吸浆虫GST、CarE和CYP450活性的影响
由表5可知,取食抗、感程度不同品种(系)小麦的麦红吸浆虫幼虫体内GST、CarE和CYP450活性差异明显。除小偃6号外,取食感虫品种小偃22、西农822和西农88的幼虫GST和CarE活性均低于4个抗虫品种,其中取食西农88的GST和CarE活性最低,分别为0.0963和 0.1016 U/mg protein,取食科农 1006和陕麦 139的最高,分别为 0.2144和 0.2284 U/mg protein。对于 CYP450,以取食小偃6号的活性最高,其次为西农 822,分别为 0.1885和 0.1791 U/mg protein,取食其他品种的幼虫间差异不显著。
2.5 麦红吸浆虫幼虫体内解毒酶活性与小麦籽粒次生物质含量的关系
对取食不同小麦品种(系)的麦红吸浆虫幼虫 3种解毒酶活性与小麦籽粒次生物质含量进行相关性分析,结果表明,GST和CarE活性与阿魏酸含量呈显著正相关(P<0.05),与单宁、总酚和总黄酮含量相关性不显著(P>0.05);CYP450活性与单宁含量呈显著负相关(P<0.05),与阿魏酸、总酚和总黄酮含量相关性不显著(P>0.05)(表6)。

表4 小麦籽粒次生物质含量与小麦抗吸浆虫指标的相关性Table 4 Correlation between resistance indicators of wheat to S. mosellena and contents of secondary metabolites in wheat kernels
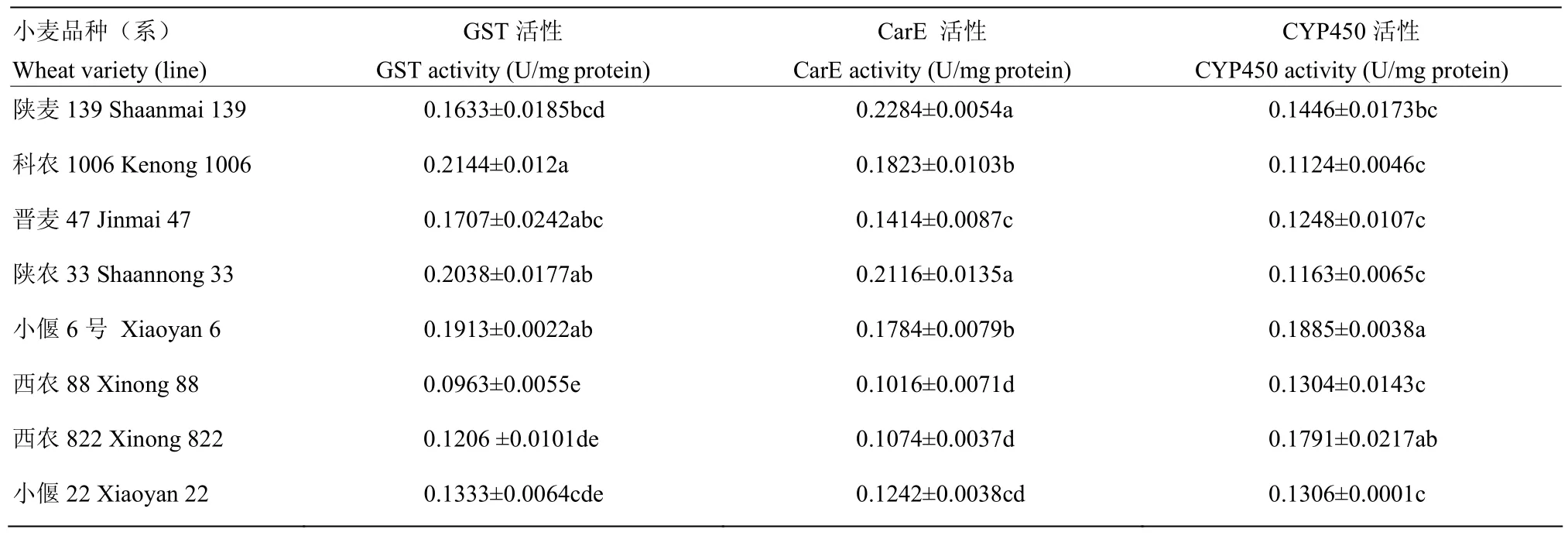
表5 取食8个品种(系)小麦的麦红吸浆虫幼虫解毒酶活性Table 5 Detoxification enzyme activities of S. mosellana larvae feeding on eight wheat varieties (lines)

表6 麦红吸浆虫幼虫体内解毒酶活性与小麦籽粒次生物质含量的相关性Table 6 Correlation between detoxification enzyme activities of S. mosellana larvae and contents of secondary metabolites in wheat kernels
2.6 小麦品种(系)对麦红吸浆虫幼虫GST1、CarE2和CYP6A1表达的影响
取食不同小麦品种(系)的麦红吸浆虫幼虫GST1和CarE2表达量存在明显差异,其中取食抗虫品种晋麦47、科农1006、陕麦139和陕农33的表达水平显著高于取食感虫品种小偃22、西农822、西农88和小偃6号(P<0.05)。GST1表达量在取食陕农33的幼虫中最高,为陕麦139的1.42倍,显著高于其他品种;在取食小偃22的幼虫中最低,为陕麦139的0.15倍。CarE2表达量在取食陕麦139的幼虫中最高,显著高于取食其他品种,在取食小偃22的幼虫中最低,仅为取食陕麦139幼虫的0.07倍(图1-A、1-B)。
取食不同抗虫性小麦品种(系)的麦红吸浆虫幼虫CYP6A1表达量没有表现出一定的规律,取食抗虫品种陕麦139、晋麦47和感虫品种小偃22、西农88的表达量较高且无显著差异,取食抗虫品种科农1006和感虫品种西农822的表达量较低且无显著差异(图1-C)。

图1 取食8个小麦品种(系)的麦红吸浆虫幼虫GST1、CarE2和CYP6A1相对表达量Fig. 1 Relative expression level of GST1, CarE2 and CYP6A1 in S. mosellana larvae feeding on eight wheat varieties (lines)
2.7 取食不同小麦品种(系)的麦红吸浆虫解毒酶活性与基因表达水平的关系
相关性分析结果表明,取食不同小麦品种(系)的麦红吸浆虫幼虫GST1表达水平与 GST活性(r=0.772,P=0.043)、CarE2表达水平与 CarE活性均呈显著正相关(r=0.756,P=0.030),CYP6A1表达水平与 CYP450活性相关性不显著(r=0.558,P=0.150)。
3 讨论
植物次生物质是化学防御的物质基础,能影响或帮助植物抵御昆虫的侵害,植物品种间抗生性差异与体内各类型次生物质组成和含量密切相关[6-9]。本研究发现小麦对麦红吸浆虫的抗性与小麦灌浆期籽粒单宁、总酚和总黄酮含量相关性不显著,但与阿魏酸含量呈显著正相关,即阿魏酸对吸浆虫抗性的贡献较单宁、总酚和总黄酮大,其含量越高,对吸浆虫抗性越强。这与小麦对麦长管蚜、玉米象及水稻对褐飞虱(Nilaparvata lugens)抗性与阿魏酸含量显著正相关的结果一致[9,29-30]。研究表明,阿魏酸不仅是构成植物细胞壁的重要成分,而且可显著抑制麦二叉蚜(Schizaphis graminum)和斜纹夜蛾(Spodoptera litura)的取食和生存[31-32],并能降低棉铃虫幼虫、蛹的体重和影响幼虫对食物的吸收与利用[7]。据此推测,阿魏酸作为小麦抗麦红吸浆虫的关键次生物质,可能通过降低麦红吸浆虫对食物的吸收效率,抑制其取食和生长发育,以此达到抵御侵害的目的。
解毒酶及解毒机制在昆虫对植物次生物质的代谢和寄主适应中起着重要作用。作为昆虫重要的解毒酶,谷胱甘肽-S-转移酶(GST)可催化还原型谷胱甘肽与各种亲电有毒次生物进行加成反应,形成亲水性更强的无毒或低毒物而排出体外[33];羧酸酯酶(CarE)则能催化含酯键、酰胺键的外源与内源次生代谢物质水解[15]。许多研究证明寄主抗虫性和植物次生物质可影响解毒酶活性及相关基因表达,以此增加对次生物质的防御适应性。绿豆象(Callosobruchus chinensis)取食抗虫绿豆品种、麦长管蚜取食抗虫品种小麦后体内CarE和GST活性显著高于取食感虫品种[34-35];饲喂阿魏酸可显著提高褐飞虱若虫GST和CarE活性及其编码基因NlGSTe1、NlGSTd1和NlCarE的表达[30];用含0.5%单宁、0.2%芦丁或1%没食子酸的饲料饲喂斜纹夜蛾后,5龄幼虫中肠和脂肪体GST和CarE活性及SlGSTe1和SlCarE表达量显著增加[12]。本研究表明,麦红吸浆虫幼虫取食阿魏酸含量高的抗虫品种小麦后,GST和CarE活性及GST1和CarE2表达量均显著高于取食感虫品种,GST和CarE活性与阿魏酸含量及对应基因表达水平显著正相关,与以上结果相符,说明GST和CarE及其基因GST1和CarE2在麦红吸浆虫对寄主小麦有毒次生物质的代谢和防御中起着重要作用。
细胞色素 P450酶系(CYP450)在昆虫对植物次生物质的解毒代谢中也发挥了重要作用,许多研究报道了植物次生物质对昆虫 CYP450活性的影响。萜类植物次生代谢物棉酚可诱导棉铃虫CYP450的活性,棉铃虫取食棉酚含量高的寄主植物后,CYP450活性显著升高[36];吲哚-3-甲醇、花椒毒素、2-十三玩酮等多种植物次生物质可诱导草地贪夜蛾(Spodoptera frugiperda)幼虫 CYP450的活性[37]。与之不同,2-十三烷酮却通过降低棉铃虫 6 龄幼虫CYP450活性,使其在含量较低的情况下具有较高的毒性[38]。本研究发现,麦红吸浆虫幼虫 CYP450活性与小麦籽粒单宁含量显著负相关,即单宁可降低CYP450活性。单宁具有靴化蛋白的特性,是多种酶促反应的抑制剂[39-40],据此推测单宁可能与吸浆虫CYP450相关蛋白结合,通过抑制其活性和作用过程,阻止或降低其解毒代谢,使虫体生命力“弱化”而发挥毒杀作用。CYP6亚家族基因代谢解毒作用是昆虫寄主适应的重要机制,不同次生物质所诱导的CYP6不同。棉酚能够诱导棉铃虫CYP6AE11和CYP6B7过量表达,而槲皮素和单宁酸仅能诱导CYP6B6的表达[36,41]。本研究发现单宁虽然能够抑制麦红吸浆虫CYP450的活性,但是对CYP6A1表达无显著影响,这可能是由于 CYP450活性由多基因控制,特定的植物次生物质对不同CYP6诱导或抑制作用不同[11,41-42],而CYP6A1可能不是麦红吸浆虫幼虫响应单宁胁迫的关键基因。
4 结论
小麦灌浆期籽粒阿魏酸是影响小麦对麦红吸浆虫抗性的主要次生物质,阿魏酸含量高的寄主能够诱导麦红吸浆虫幼虫GST和CarE活性及编码基因GST1和CarE2的表达,这些酶及基因在麦红吸浆虫对阿魏酸的解毒代谢和防御反应中发挥着重要作用。单宁却通过降低吸浆虫幼虫CYP450的活性使其在含量较低的情况下具有相对高的毒杀作用。
