哈尼梯田生态系统土壤微生物量碳的影响因素
2020-11-13宋维峰
李 荣,宋维峰
西南林业大学,生态与环境学院,昆明 650224
土壤中的微生物类群是重要的地下生物宝库,也是全球生态系统生物多样性的一个重要组成部分,在林业可持续发展中扮演重要角色[1]。土壤微生物可以形成土壤结构、分解矿物质、具有固氮作用,在植物根系周围生活的土壤微生物还可以调节植物生长,与植物根部营养有密切关系[2]。土壤微生物是土壤有机质的活性部分,也是土壤中最活跃的因子,是土壤有机质和土壤养分(氮、磷、硫等)转化和循环的动力,它参与有机质的分解、腐殖质的形成、土壤养分转化和循环等各个生化过程。农业中种植制度、轮作措施、有机残体的投入和施肥均对土壤微生物量有较大的影响。而微生物本身是有机物分解、养分转化的中介者,它们的数量直接影响着进入土壤中有机物和污染物的分解转化速度,因而影响作物的生长和环境的质量。土壤微生物量碳(MBC,Microbial Biomass Carbon)虽然只占土壤有机碳的1%—4%,但占微生物干物质的40%—45%,是反映土壤微生物生物量大小的重要指标,也是土壤有效养分重要的源和汇[3]。随着全球碳循环问题受到广泛关注,微生物生物量碳特别是土壤微生物量碳及影响因素开始引起人们的重视。土壤微生物量碳的影响因素繁多,土壤类型、人为活动、土地利用类型、土壤理化性质、植被类型、管理措施等都对其有影响。
哈尼梯田生态系统在2010年中国西南部的特大旱灾中因免受灾害而受到广泛的关注,其完善的生态系统和独特的地理环境使梯田蓄水充足,水资源得到充分的利用。哈尼梯田生态系统(图1)公路上方的水源区由于保护良好,基本不受人为干扰,公路下方的梯田区受到人为干扰较大。目前哈尼梯田生态系统的研究大都集中于人文、景观格局、生态系统功能、生物多样性等方面。对于哈尼梯田微生物方面虽有部分研究,梁勇等[5]发现元阳梯田稻田土壤微生物数量总体上在低海拔(1450、1500 m)稻田较高,土壤自生固氮菌和放线菌在3月、细菌和真菌在5月、纤维分解菌在9月的数量相对较多,且微生物数量受季节的明显影响,并表现出明显的垂直分布特征。冯书华等[6]研究发现在红果树和山龙眼森林群落中,微生物数量随土壤深度增加而减少,与土壤全N含量显著正相关。赵娟等[7]研究发现元阳地方水稻根部内生细菌及根际土壤细菌具有一定的种属同源性与特异性。王灿等[8]UV-B辐射增强显著降低水稻孕穗期、抽穗期和成熟期稻田土壤微生物量碳;翟精武等[9-10]发现外源碳氮输入和土壤氮素净硝化量都与元阳梯田土壤微生物量碳呈显著正相关关系。但哈尼梯田微生物方面的研究局限于森林或梯田的单独研究,对森林和梯田土壤微生物量碳的研究及对比尚未报道。土壤微生物作为生态系统健康的指标,研究其分布对保护生态系统具有重要的意义。哈尼梯田是以哈尼族为主的民族利用哀牢山区地貌、气候、植被、水土等立体特征,创造出集森林生态系统、村寨生态系统、梯田生态系统与河流生态系统于一体的复合生态系统,其上部的森林生态为下部的梯田提供充足的水源,森林与梯田相互依存。本研究以哈尼梯田生态系统水源区乔木林、灌木林和荒草地3种土地利用类型与梯田区土壤MBC含量进行比较,选取不同土层(0—20、20—40和40—60 cm)土壤,研究了该地区土壤MBC在时空上的分布特征,分析土壤MBC在水源区的自然条件下与梯田区受到强烈人为活动影响下的含量变化,探讨相关环境因子对土壤MBC含量的影响机制,为森林和梯田生态系统合理管理提供理论依据。
1 研究区概况
研究区位于云南省哀牢山南段,地处东经101°48′—103°38′,北纬22°33′—23°08′。该区域由于受到构造侵蚀和剥蚀作用,形成深切割的中山地貌,最高海拔3074.3 m,最低海拔105 m,属山地季风气候,四季不分明,干湿季分明。年平均温度18—20℃,全年日照达2000 h以上,年降雨800—1800 mm。由于海拔高差大,气候、土壤、植被等具有明显的垂直变化特征。研究区水源区土壤为红壤,梯田区土壤为黄红壤,土层厚度约为100 cm。研究区的乔木主要为红木荷(Schimakhasiana)、旱冬瓜(Alnusnepalensis)、桤木(Alnuscremastogyne)、元江山茶(Camelliajaponica)、尖子木(Oxysporapaniculata)等;灌木主要为浆果栎(Quercuscoccifera)、叶下珠(Phyllanthusurinaria)、槐树(Sophorajaponica)、菝葜(Smilaxchina)、荚蒾(Viburnumdilatatum)、野牡丹(Melastomacandidum)、悬钩子(Rubuscorchorifolius);草本主要为鳞毛蕨(Dryopterisfilix-max)、紫茎泽兰(Eupatoriumadenophorum)、沿阶草(Ophiopogonbodinieri)、芒萁(Dicranopterisdichotoma)、荩草(Arthraxonhispidus)、车前草(Plantagoasiatica)、旱生卷柏(Selaginellastauntoniana)、窃衣(Torilisscabra)、水莎草(Cyperusglomeratus)[11-13]。
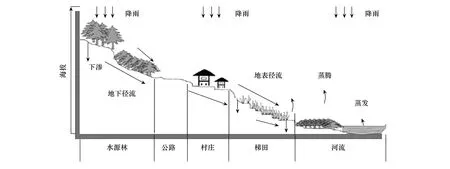
图1 哈尼梯田立体结构图[4]Fig.1 Three-dimensional structure of Hani Terrace
2 研究方法
2.1 样地设置与调查
2018年7月在研究区进行植物多样性调查的基础上,把研究区分为水源区与梯田区。水源区分为乔木林、灌木林、荒草地3种不同土地利用类型,每个土地利用类型选择3块样地进行采样;梯田区按高程从公路(高程1880 m)下方开始,每隔20 m高程设置一个采样区,分为顶部梯田、上部梯田、中部梯田、下部梯田4个部位,每个部位选择3块样地进行采样。对每块样地进行植物调查,记录样地中所有植物的生长情况。样地基本情况见表1。
2.2 样品采集
本文研究取样方法为五点交叉取样法,频率为1次/季度,采样时间为2018年7月(夏季)、2018年10月(秋季)、2018年12月(冬季)和2019年3月(春季)。水源区每个采样点采集三个不同深度土层(即0—20 cm土层、20—40 cm土层、40—60 cm土层)的土样;梯田区因长期浸水分层采土有困难,且梯田种植水稻根系较浅,所以每个采样点采集两个不同深度土层(即0—20 cm土层、20—40 cm土层)的土样。本文研究土壤样品分成两份,一份采集完放入便携式冰箱,4℃保存,带回实验室于一周之内测定土壤MBC,于一个月之内测定铵态氮、硝态氮;一份自然风干,过筛,测定全氮、有机碳、pH等指标。每个土样每个指标分别进行3次重复试验。
2.3 测定方法
因梯田土壤长期淹水,为确保实验结果准确,故对梯田土壤MBC测定采用两种测定方法,即液氯熏蒸浸提-水浴法[14]和氯仿熏蒸法[3],结果显示两种方法所得的土壤MBC含量相差不大,故本文梯田区及水源区土壤MBC的测定采用氯仿熏蒸法[3];含水率的测定采用烘干法[15];容重和孔隙度的测定采用环刀法[15];全氮的测定采用凯氏定氮法[16];硝态氮[17]和铵态氮[18]的测定采用紫外分光光度法;pH值的测定采用电位法[15];有机碳的测定采用重铬酸钾氧化法测定[19]。
2.4 数据处理
采用Excel 2010与SPSS 24.0软件对数据进行统计分析,利用双因素方差分析(Two-way ANOVA)及Duncan多重比较法检验不同土地利用类型土壤MBC的差异性(α=0.05),采用Spearman法对土壤MBC含量与地上植物、土壤理化性质及相关环境因子进行相关分析。采用Excel 2010软件制图。图表中数据为平均值±标准差。
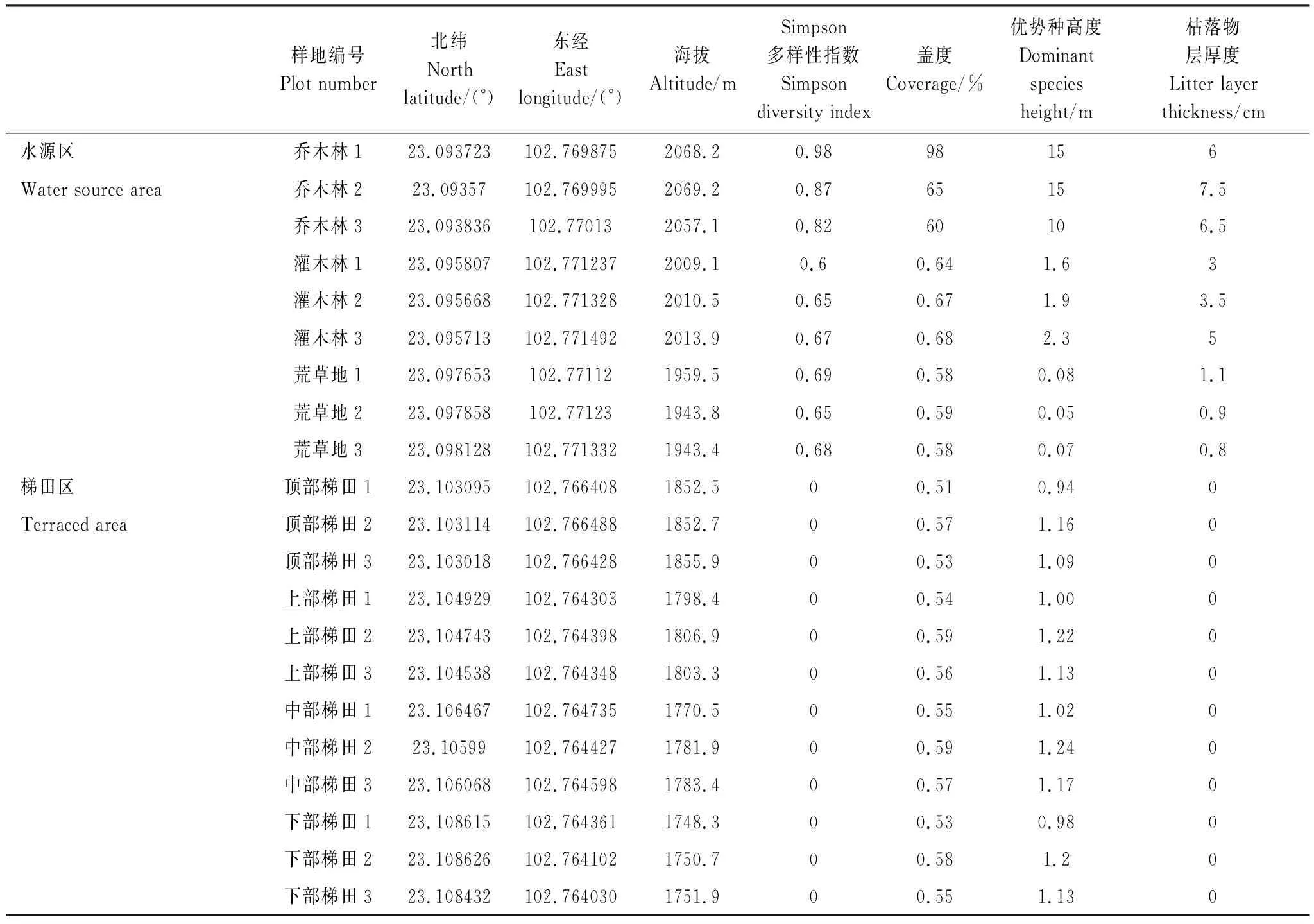
表1 采样点基本情况及植物统计表
3 结果与分析
3.1 不同土地利用类型土壤MBC含量变化
图2是不同季节样品的平均值,从图中可以看出同一土层中4种不同土地利用类型之间的差异显著。4种不同土地利用类型的表层土壤(0—20 cm)的土壤MBC含量都显著高于下层土壤(20—40 cm、40—60 cm),乔木林、灌木林和荒草地20—40 cm土层的土壤MBC含量显著高于40—60 cm土层,说明随着土层的加深,4种不同土地利用类型土壤MBC含量也随之下降,乔木林最为明显,0—20 cm土层土壤MBC含量是40—60 cm土层的3.19倍。同一土地利用类型不同土层之间的差异显著,水源区不同立地类型不同土层之间土壤MBC的含量变化较为一致,除40—60 cm土层荒草地略高于灌木林外,其他皆为乔木林地最高,灌木林次之,荒草地最低,乔木林0—20 cm的土壤MBC依次是灌木林、荒草地的1.78和3.46倍,20—40 cm的是灌木林、荒草地的1.72和2.29倍,40—60 cm的是灌木林、荒草地的2.19和1.90倍。梯田区4个不同部位的梯田土壤微生物量含量基本一致,顶部梯田整体土壤MBC含量都低于下部梯田,其0—20cm土壤MBC含量是40—60cm的1.17—1.63倍。

图2 4种土地利用类型不同土层土壤微生物量碳含量Fig.2 Soil microbial biomass carbon content in different soil layers of four land use types不同大写字母表示同一土层不同立地类型差异显著;不同小写字母表示同一立地类型不同土层差异显著
3.2 不同土地利用类型土壤MBC季节变化
图3是水源区不同土地利用类型不同季节土壤微生物量碳含量情况,从图3可以看出,水源区3种不同土地利用类型的土壤MBC具有明显的季节动态规律,夏季MBC偏高,冬季MBC含量偏低。3种土地利用类型不同季节的土壤MBC含量都随土层的加深而降低,但不同立地类型随季节的变化也各不相同,乔木林不同土层随季节的变化基本一致,秋季的表层(0—20 cm)土壤及深层(40—60 cm)土壤MBC含量较其他三季低,乔木林表层(0—20 cm)和深层(40—60 cm)土壤MBC含量最大值分别是462.62 mg/kg与182.10 mg/kg;灌木林夏季表层土壤MBC含量远远高于其他三季,春、秋、冬三季相同土层含量基本一致,灌木林表层(0—20 cm)和深层(40—60 cm)土壤MBC含量最大值分别是311.37 mg/kg与65.40 mg/kg;荒草地秋季表层土壤MBC含量高于夏季,在深层土壤时急速下降,远远低于夏季,荒草地表层(0—20 cm)和深层(40—60 cm)土壤MBC含量最大值分别是146.82 mg/kg与117.06 mg/kg。
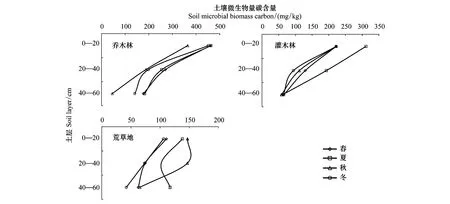
图3 水源区不同土地利用类型不同季节土壤微生物量碳含量Fig.3 Soil microbial biomass carbon content in different seasons of different land use types in water source area
图4是梯田区不同季节土壤微生物量碳含量情况,从图4可以看出,梯田区的土壤MBC具有明显的季节动态规律,顶部和上部梯田体现出“夏高冬低”的特点;中部和下部梯田体现出“夏秋高冬春低”的特点。4个不同部位不同季节的土壤MBC含量最大值也各不相同,梯田上部不同土层的含量变化不大,秋季的深层(20—40 cm)较表层(0—20 cm)土壤MBC含量下降较为明显,梯田上部表层(0—20 cm)和深层(20—40 cm)土壤MBC含量最大值分别是79.01 mg/kg与76.69 mg/kg;梯田上部不同土层随季节变化规律基本一致,其表层(0—20 cm)和深层(20—40 cm)土壤MBC含量最大值分别是138.81 mg/kg与103.66 mg/kg;梯田中部不同土层之间土壤MBC变化幅度较大,其表层(0—20 cm)和深层(20—40 cm)土壤MBC含量最大值分别是134.99 mg/kg与81.85 mg/kg;梯田下部秋季表层土壤微生物含量最高,夏季深层土壤MBC含量最高,其表层(0—20 cm)和深层(20—40 cm)土壤MBC含量最大值分别是166.29 mg/kg与118.03 mg/kg。
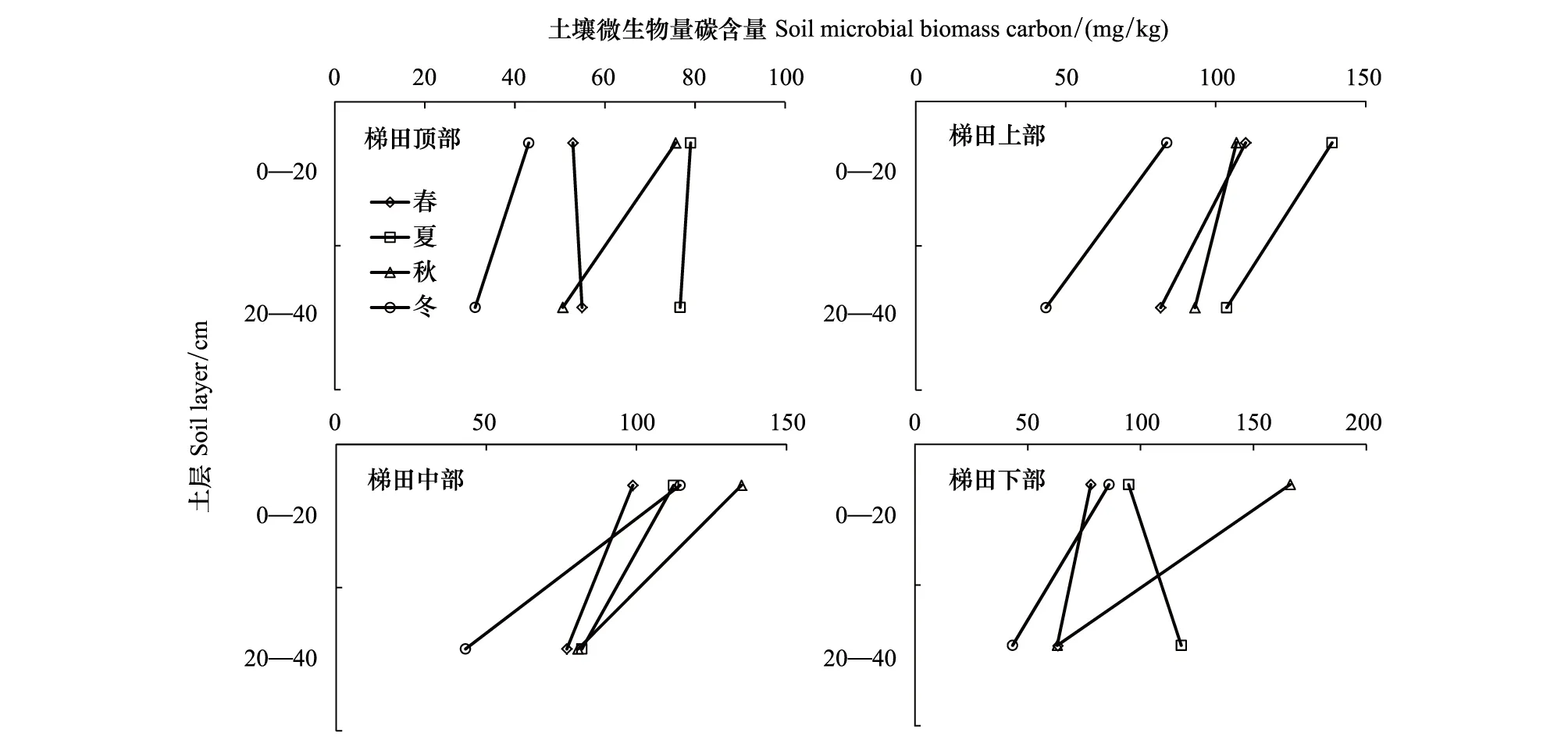
图4 梯田区不同季节土壤微生物量碳含量Fig.4 Soil microbial biomass carbon content in different seasons of terraced area
3.3 地上植被对土壤MBC的影响
对4种不同土地利用类型的土壤MBC含量与地上植被的多样性指数、盖度和优势种平均高度进行Spearman相关分析,结果如下。
3.3.1水源区地上植被对土壤MBC的影响
(1)就不同土壤层次来看
表2是水源区不同土层土壤微生物量碳与地上植被的相关系数,由表2可以看出,水源区0—20 cm土层的土壤MBC含量与其地上植被各项因子呈显著的正相关性,说明0—20 cm土层的土壤MBC与地上植被的植物多样性指数、盖度、优势种高度和枯落物层厚度都有很强的相互关系,地上植被的生长状况会直接影响土壤MBC含量。20—60 cm土层的土壤MBC与植物多样性、盖度和优势种高度呈显著的正相关性,与枯落物层厚度的相关性比较弱。总体来说,水源区不同土地利用类型相同土层的土壤MBC与地上植被的生长状况具有较强的相互关系,随土层的加深植物对土壤MBC含量的影响在减弱。
(2)就不同土地利用类型层次来看
表3是水源区不同土地利用类型土壤微生物量碳与地上植被的相关系数,由表3可以看出,水源区乔木林、灌木林、荒草地3个层次的土壤MBC含量与其地上植被的各项因子无显著的相关性,说明在相同土地利用类型下不同土层的土壤MBC与地上植被的相互关系较弱。

表2 水源区不同土层土壤微生物量碳与地上植被的相关系数
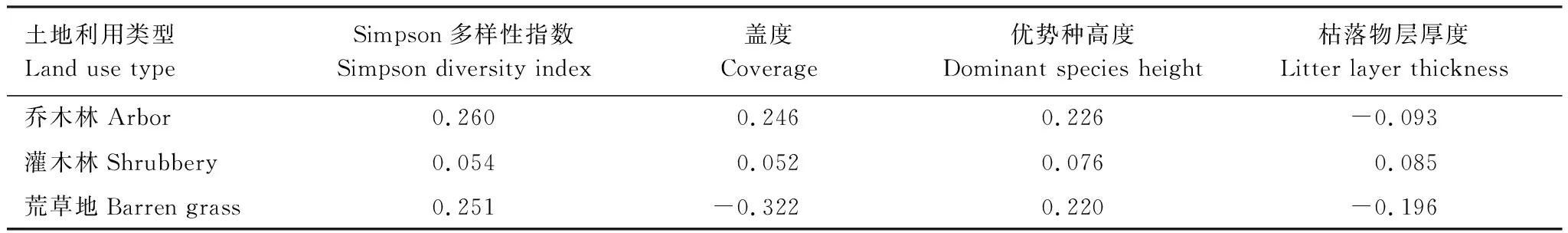
表3 水源区不同土地利用类型土壤微生物量碳与地上植被的相关系数
3.3.2梯田区地上植被对土壤MBC的影响
梯田区因受人为干扰强烈,其种植作物单一,植物多样性指数为0,枯落物层厚度也为0 cm,故在分析时不考虑这两个因子。
(1)就不同土壤层次来看
表4是梯田区不同土层土壤微生物量碳与地上植被的相关系数,由表4可以看出,梯田区0—20 cm土层的土壤MBC含量与地上植被盖度呈显著正相关性,优势种高度与其没有相关性,说明0—20 cm土层的土壤MBC与地上植被的盖度有很强的相互关系。40—60 cm土层的土壤MBC与地上植被的各项因子均无相关性。总体来说,梯田区种植水稻的盖度与土壤MBC有一定的关系。

表4 梯田区不同土层土壤微生物量碳与地上植被的相关系数
(2)就不同海拔层次来看
表5是水源区不同土地利用类型土壤微生物量碳与地上植被的相关系数,由表5可以看出,梯田顶部、梯田上部、梯田中部和梯田下部4个层次的土壤MBC含量与其地上植被的各项因子无显著的相关性,说明在相同海拔下不同土层的土壤MBC与地上植被的相互关系较弱。
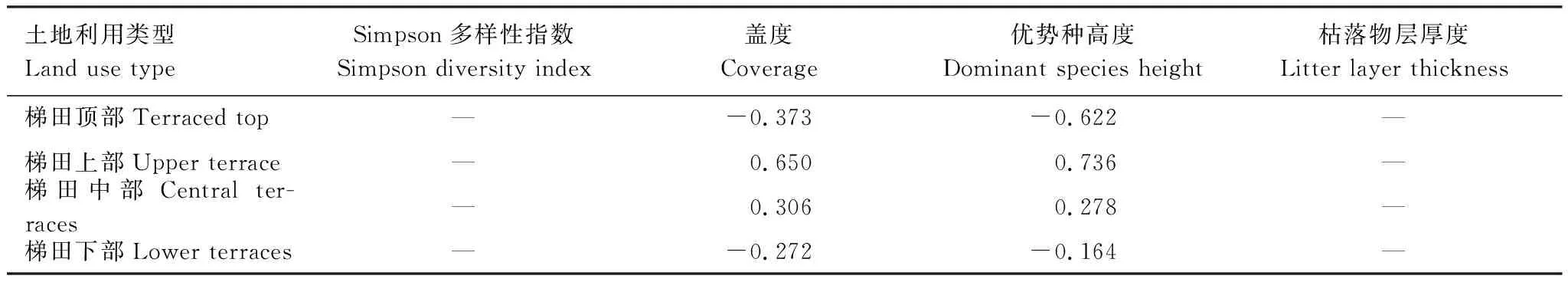
表5 梯田区不同土地利用类型土壤微生物量碳与地上植被的相关系数
3.4 土壤MBC含量与土壤理化性质的关系
对乔木林、灌木林、荒草地和梯田4种不同土地利用类型的土壤MBC含量与土壤理化性质进行Spearman相关分析(表6),结果显示:土壤MBC与土壤的有机碳和孔隙度呈显著正相关性,与土壤所在海拔也呈显著正相关性,而与土壤容重呈显著负相关性。土壤MBC与全氮、铵态氮、pH值、含水率均无显著的相关性。

表6 土壤微生物量碳含量与理化性质相关性分析
4 讨论
4.1 不同土地利用类型土壤MBC含量变化
不同森林类型、不同耕作措施都对土壤MBC有影响,土地的利用方式、人工干扰强度及土壤的理化性质都对土壤MBC有影响[20-24]。本研究中,水源区3种土地利用类型土壤MBC平均含量的范围在62.17—434.42 mg/kg,低于其他森林的结果[21-22],产生差异的原因可能是土壤理化性质和气候条件不同造成的;梯田土壤MBC平均含量范围在53.35—115.18 mg/kg,远远小于天然湿地[25-26],与垦殖的湿地差别不大[27],产生的原因可能是哈尼梯田属于人工湿地,但由于长期的耕作导致土壤MBC减少。此外,本研究中不同土地利用类型的土壤MBC含量随土层的增加而降低,这与在其他地区的研究相符[21]。
本研究区的水源区人为干扰较弱,梯田区有很强的人为干扰。水源区土壤MBC含量最高的为乔木林,最低的为荒草地,3种不同土地利用类型受其林地组分、土壤养分影响而成规律性变化。梯田区4个不同部位土壤MBC含量基本一致,含量普遍较低,0—20 cm土层土壤含量是乔木林的0.14—0.26倍,灌木林的0.25—0.47倍,荒草地的0.50—0.92倍,说明梯田土壤MBC含量低于未受人工干扰的其他土地利用类型。荒草地因土壤肥力较弱,导致其土壤MBC含量较低,梯田因种植水稻在一年之中被翻土、耕种、长期浸水而使其土壤MBC含量偏低。
4.2 不同土地利用类型土壤MBC季节变化
本研究中水源区3种不同土地利用类型的土壤MBC具有明显的季节变化规律,夏季偏高冬季偏低,与前“夏高东低”的“单峰型”结论有所差异[21,28],产生的原因可能是研究区因其特殊的地理结构,气温季节变化不大,夏季与冬季相差9.5℃[29],其土壤MBC具有明显的季节变化规律,但不如其他地方的变化明显。梯田区的土壤MBC随季节变化规律为“夏秋高冬春低”,产生的原因可能跟梯田种植作物相关,梯田每年5月插秧10月收割,夏、秋季梯田内种植有作物,其MBC含量较春、冬季高。
水源区夏季MBC含量是冬季的1.02—2.78倍,梯田区夏季MBC含量是冬季的1.10—2.45倍,由图2和图3可以看出,梯田土壤MBC季节变化比水源区明显,变化幅度较大。梯田土壤在种植作物生长期间(夏季、秋季)受到的干扰较小,在种植作物之前需要整田,收割作物时需要把梯田内水排出,并在收割结束之后要翻土并继续淹水,在无作物种植期间(春季、冬季)其人为干扰很强,使其MBC含量大大降低。
4.3 地上植被对土壤MBC的影响
土壤微生物与地上植被关系密切,Chen Chen[30]通过106篇文献进行全球性的meta分析,得出对于森林而言10%植物丰富度的下降会导致微生物量下降5%。地上植被多样性对土壤MBC也有着不同程度的影响,不同林分、多样性、植物生长情况都对土壤MBC有着不同程度的影响[31-33]。本研究区,不同土地利用类型的植被生长情况不同,乔木林植被覆盖度高、植物种类多、植被高度较高;灌木林植被覆盖度高但植物种类少;荒草地植被覆盖度高但植被少而低矮,导致枯落物很少。
在本研究中,水源区就不同土壤层次来看,不同立地类型的每一层土壤MBC与植物多样性指数、盖度、优势树种高度、枯落物层厚度都有很强的相关性,说明地上植被的生长情况与地下土壤MBC有很强的相关关系;就不同土地利用类型层次来看,在相同土地利用类型不同土层的土壤微生物量情况与地上植物的相关性不显著,产生这样的情况可能的原因是本研究相同土地利用类型的采样点相隔近,采样点所处大环境与气候条件一致,导致相同土地利用类型的地上植被各项因子与地下土壤MBC关系不显著。梯田区就不同土壤层次来看,仅梯田区0—20 cm土层的土壤MBC含量与地上植被盖度呈显著正相关性;就不同海拔层次来看,相同海拔下不同土层的土壤MBC与地上植被的相互关系较弱。产生这样的原因可能是梯田区因受人为干扰强烈,种植、收割、翻耕、长期淹水等影响,其地上植被单一,仅为种植的水稻,无枯落物层覆盖,其地上植被各项因子几乎没有差异,且采样点所处大环境与气候条件一致。
4.4 土壤MBC含量与土壤理化性质的关系
影响土壤MBC的因素有很多[34],本研究从土壤有机碳、土壤全氮、土壤铵态氮、土壤硝态氮、土壤pH值、土壤含水率、土壤容重、土壤孔隙度和土壤所在的海拔来进行分析。本研究中土壤MBC与土壤的有机碳有显著的正相关关系,与前人所做研究结论一致[21,35],说明本研究区土壤有机碳与土壤MBC含量具有紧密联系。此外土壤MBC与孔隙度呈显著正相关关系,不同土地利用类型和耕作措施会影响土壤的孔隙度进而影响土壤营养状况[36-37],本研究结果与前人研究结论一致[38],说明本研究区土壤孔隙度对土壤MBC含量影响较大。4种不同土地利用类型土壤含水率与土壤MBC没有显著相关性,与前人研究结论相同[34,39-40],说明了在本研究区域内土壤含水率并不是影响4种土地利用类型土壤MBC的重要因素,可能原因是研究区雨水比较丰富、各季节水源区和梯田区土壤的含水率相差不大有关。
综上所述,在哈尼梯田生态系统中,不同的土地利用类型的土壤MBC含量差异显著,人为干扰对土壤MBC的影响很大。因此,在水源区应保护好森林,尽量减少人类活动,特别是对土壤的保护,不应再挖毁水源区土壤,避免因不恰当的人类活动减少土壤微生物量从而破坏整个生态系统,这对哈尼梯田的可持续发展具有重大意义。
