山西铁桥山省级自然保护区华北豹及其猎物的时空动态研究
2020-11-13罗文慧高春雨李吉照李春实唐一鸣姜广顺
罗文慧,高春雨,李吉照,李春实,唐一鸣,王 姣,姜广顺,华 彦
1 东北林业大学 野生动物与自然保护地学院,哈尔滨 150040 2 广东省林业科学研究院森林培育与保护利用重点实验室,广州 510520
物种在群落中如何共存是群落生态学的关键问题,也是理解群落多样性的关键,其中种间关系(包括竞争、捕食等)是解释共存格局的因子之一[1]。种间竞争的物种通常通过生态位分化(时间、空间、食物等)达到共存,而食肉动物在猎物丰富时,通过与猎物在空间、时间生态位的重叠提高捕食率[2-3]。物种生态位分化的信息有助于理解同域分布物种的生存状况,为生物多样性管理与保护提供建议。放牧对同域分布的野生动物的影响是全球性保护问题[4],高密度的家畜不仅增加竞争排斥,导致同域分布的食草动物数量减少[5],并且对于放养的家畜与同域分布的食草动物而言,体型和觅食策略相似物种的相互作用可能会降低至少一个物种的适应度[6-7],而当天然猎物降低到一定阈值,会使大型猫科动物捕食家畜的概率上升[8]。野生动物捕食家畜,不仅让当地居民对野生动物容忍度降低[9],甚至可能会引起对其产生报复性猎杀。目前,同域物种的生态位分化差异主要集中于关于植物群落研究,生态相似种的共存机制研究,或者对野生动物的单一资源生态位(时间、空间或营养)研究,尚缺乏综合时间和空间资源情况时家畜与野生动物的相互干扰、竞争研究[10-12]。
华北豹(Pantherapardusjaponesis)是仅分布于中国华北地区的豹亚种[13],也是铁桥山省级自然保护区的主要保护对象。铁桥山自然保护区位于和顺县境内,该县20世纪70年代起就开始黄牛改良工作,至21世纪初,稳健增长的肉牛养殖业已成为发展农村经济的优势产业之一[14]。家畜与华北豹野外猎物在资源利用上形成竞争,而竞争关系的物种会从空间、时间和食物进行资源划分,进而分化生态位,这有利于缓解同域生存压力[2]。并且有研究表明,哺乳动物的资源选择利用情况会因人为干扰而改变[6,15-16]。目前,华北豹相关研究主要为种群数量与分布、肠道真菌多样性和圈养种群饲养繁殖,仍缺少华北豹与猎物的资源选择利用情况,以及放牧对华北豹及其猎物种群的动态影响[17-19]。
本研究基于红外触发相机捕捉数据分析放牧影响下华北豹及其猎物的空间共存和活动规律,期望到达以下目标:(1)通过占域模型分析不同时期物种之间空间共存关系;(2)再运用重叠系数综合分析不同物种的时空资源竞争、干扰情况;(3)通过核密度估计物种的日活动规律,探讨不同物种的时间分配。
1 研究地概况
山西铁桥山省级自然保护区地处太行山中段,地理坐标113°05′—113°35′E,37°13′—37°34′N,占地面积为38974 hm2。海拔高差约900 m(海拔900—1800 m)。保护区气候属暖温带大陆性季风气候,年平均气温为6.3 ℃,年降水量593 mm。保护动物包括华北豹在内17种国家重点保护野生动物,植被类型有油松林、辽东栎林、混交林,以及天然灌丛。
历史原因,保护区内现有约9000户居民,居住地涉及2个乡(镇)47个村。保护区地处黄土高原地带,草坪面积较大,草料资源丰富,境内的和顺县也被列为“国家级黄牛改良基地县”,自21世纪初肉牛饲养量逐年增加[14],并且部分农户为了减少人力成本甚至散养家畜。

图1 红外相机架设与华北豹拍摄位点示意图Fig.1 Infrared camera erection and north China leopard shooting site diagram
2 研究方法
相机架设时间为2018年10月至2019年5月,红外相机型号为Ltl Acorn-6210MM,在地理信息系统(GIS)中用4 km×4 km的网格将保护区覆盖,以每个网格中2台相机的密度布设相机,每个样点在架设3个月时,更换SD 卡和电池,保证相机能够正常连续工作。为减少相邻位置之间潜在的空间自相关,每台相机位点之间距离大于300 m[20],相机架设地点(图1)一般选择山脊与水源附近,主要靠近动物活动路径的位置,所有相机陷阱均不放置任何诱饵。相机参数设置为照片(连拍3张)、时间间隔(30 s)、视频(10 s),为减少重复拍摄,设置灵敏度为低。
2.1 数据分析
2.1.1物种间单季节模型
依据村民放牧习惯,将研究时期的12月至翌年3月划分为圈养期,研究期的其它月份为散养期。运用Presence软件的双物种单季节占域模型(Two-species single season model)[21]分析比较不同时期物种间空间共存关系的差异。本研究通过将每个监测点的相机捕获时间以15 d构建探测历史,在有监测到动物出现的相机位点对应时间的单元格内填“1”,否则为“0”[22]。根据最优模型(ΔAIC<2)估计的6对物种分别在家畜的圈养期和散养期的物种互作因子(SIF,species interaction factor),可以反映出物种空间共存模式。其中,SIF=1 时表明物种间的空间分布独立发生(没有共存和回避);SIF>1 时表明物种对发生在同一地点的概率比预期的高,空间分布上趋向于共存;而SIF<1时表明物种对发生在同一地点的概率比预期的低,空间分布上趋向于回避[23]。
2.1.2日活动规律及时间重叠

3 研究结果
2018年10月—2019年5月,铁桥山自然保护区红外相机总捕获日为12190 d。62个相机位点中53个有效,其中19个位点拍摄到华北豹,网格占域率为0.36。
3.1 物种间空间共存关系
双物种单季节占域模型结果(表1)显示,在家畜散养期,华北豹-狍和华北豹-野猪的空间回避作用都比家畜圈养期弱,华北豹-牛空间回避作用最强。从家畜散放到圈养时期,狍-野猪的空间作用关系从共存转为空间回避。而牛-狍和牛-野猪的空间作用关系都为空间共存。
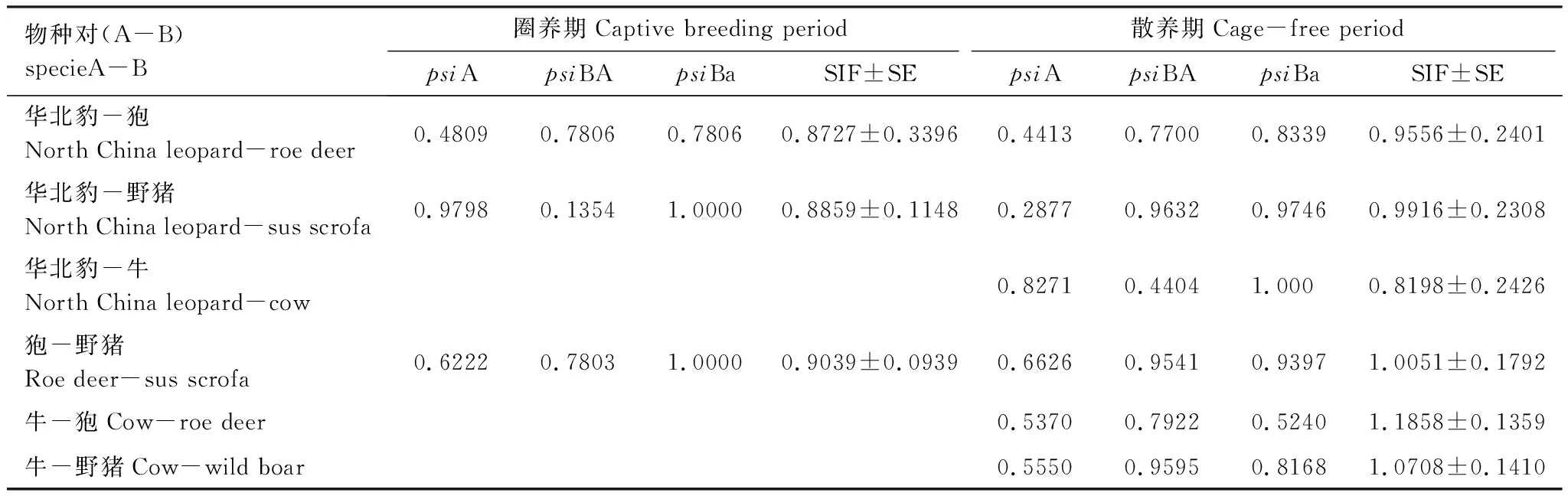
表1 不同时期物种间共存关系结果
3.2 日活动节律
华北豹、家畜、西伯利亚狍(Capreoluspygargus)和野猪(Susscrofa)在散养期的有效探测次数分别是21、628、236和138次;华北豹、西伯利亚狍和野猪在圈养期的有效探测次数分别是38、176和54次。
华北豹、狍都为昼间活动(图2),并且两者在家畜圈养期的夜间活动有所增加。牛的日活动节律也为昼行性(图2)。野猪在不同时期的日活动节律有较大变化,在家畜散养期,野猪活动节律为晨昏性,而家畜圈养期则为昼行性,并且夜间活动也较活跃(图2)。
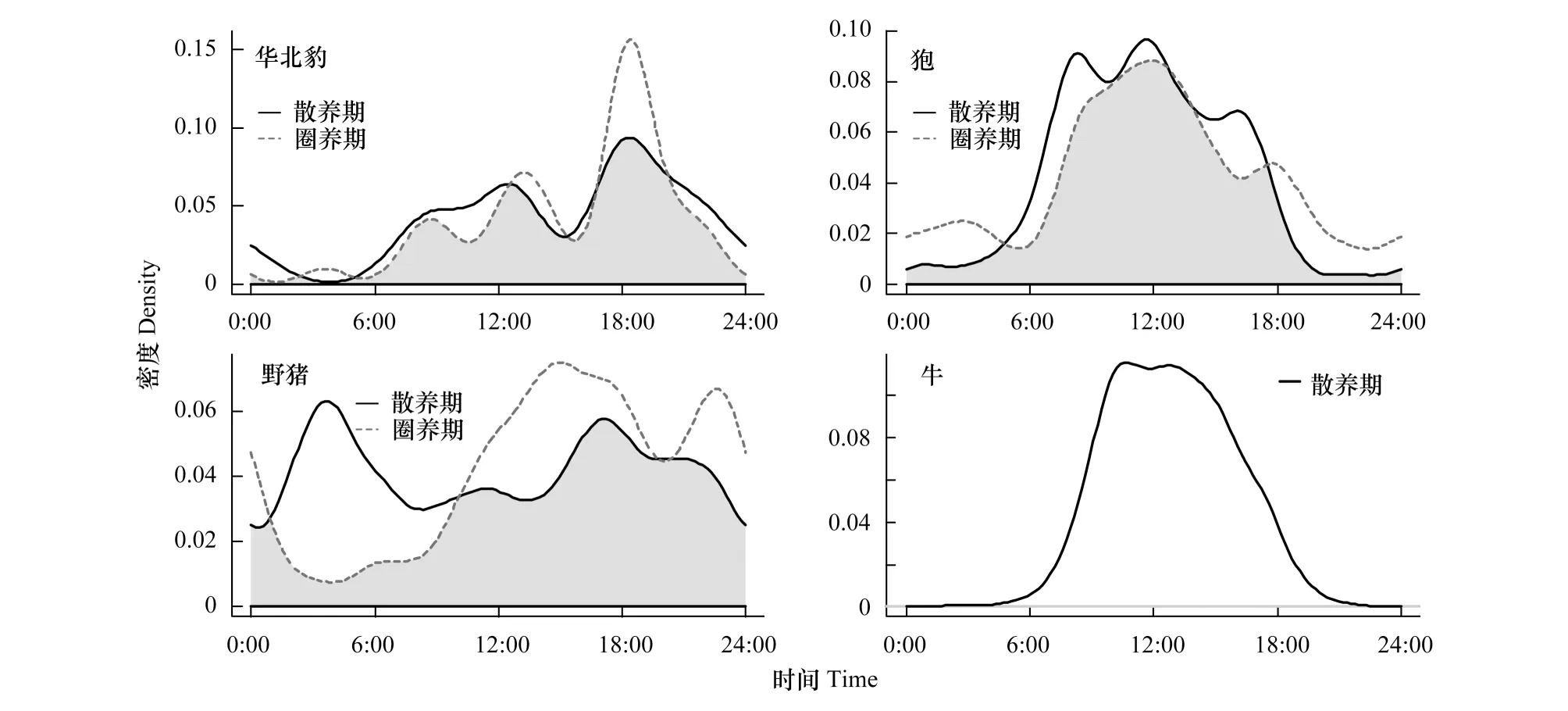
图2 4种哺乳动物日活动节律Fig.2 The daily activity patterns of four kinds of mammals
3.3 时间重叠
自家畜被散放到被圈养的两个时期里,三对物种间(无论是捕食还是竞争关系)日活动节律的重叠系数都变大,其中,狍和野猪变化最大(图3,图4,图5),散养期两者的重叠系数0.58(95%的置信区间为0.57—0.66)。圈养期两者重叠系数为0.70(95%的置信区间为0.56—0.82)。华北豹与野猪重叠系数由散养期0.69(95%的置信区间为0.58—0.85)变化至圈养期0.73(95%的置信区间为0.56—0.84)(图3,图4)。华北豹与狍的重叠系数不同时期变化最小(散养期0.62—圈养期0.65)(图3,图4,图5)。牛与狍的昼夜活动的重叠程度与其他物种间比都是最大的(重叠系数0.81(95%置信区间为0.74—0.86))(图3,图5)。捕食关系中,华北豹与牛日活动节律重叠最小(95%的置信区间为0.39—0.75))(图3,图5)。总体来看,牛与野猪的重叠系数最小,两者在散养期只有0.48(95%置信区间为0.39—0.55)(图3,图5)。

图3 4种哺乳动物在家畜散养期的日活动节律比较Fig.3 Comparison of daily activity patterns of four kinds of mammals in Cage-free period

图4 4种哺乳动物在家畜圈养期的日活动节律比较Fig.4 Comparison of daily activity patterns of four kinds of mammals in Captive breeding period

图5 不同时期物种间重叠系数Fig.5 The coefficient of overlap for two species in different periods
4 讨论
4.1 物种时空资源的利用动态
当同域分布的物种用同一方式利用同种资源时,会以相互补偿的机制反映在利用食物、空间、时间资源上的差异和竞争[25]。家畜在保护区散养时期,华北豹-狍、华北豹-野猪的空间回避作用都比无家畜存在时弱,可能是时间重叠程度较圈养期小的补偿机制。成年肉牛体型大,故豹会较多地捕食小牛(一般为5月出生)。而本研究调查期间是10月—翌年5月,故可能造成华北豹与牛的空间、时间资源重叠都不高。并且也有研究显示家畜对大型食肉动物的出现没有显著影响,在Siilkhem国家公园,家畜对雪豹的空间占域没有影响[26]。故家畜对大型食肉动物的影响仍待考究。
有蹄类动物在冬季极端天气影响下因食物资源匮乏,食性有较高重叠,而这种种间竞争就会干扰物种对其他资源的利用[27]。故在食物匮乏的家畜圈养期,狍-野猪的空间互作呈现回避。而在食物资源丰富的家畜散养期,狍-野猪空间利用趋于重叠。根据最优取食理论分析,动物在食物量资源丰富时选择最优食谱,并且已有研究表明狍与野猪都偏好针阔混交林[28-29],这就促使两者在空间资源利用上的重叠,然而狍与野猪却又以时间生态位的分化巧妙地缓解竞争。这一机制使同域分布的有蹄类动物实现共存。
家畜与狍的时间生态位有较高重叠,然而空间的共存愈发加重了种间竞争。高斯假说提出在资源有限时没有两个物种能在同一生态位共存,并且生态位重叠越多,竞争越激烈[30]。相反,野猪-牛空间资源利用上虽有重叠,但野猪与狍的时间分化,进而使野猪与牛的日活动节律大致错开了活动高峰期,为两者的共存提供可能。但在高强度放牧下植被退化对野生动物的影响仍不容忽视[31],研究表明体型大的家畜可减少灌木草本层的植物生物量,进而降低有蹄动物的生境质量[32]。生境质量的优劣也会影响野生动物资源利用方式,对预测保护物种生存现状和未来潜在生存环境负荷量有重要意义。
4.2 物种日活动规律
本研究对华北豹的日活动节律进行分析,虽结果的稳健性仍待探究,但是也有研究显示豹为昼间活动,与虎的活动节律形成时间活动上的分离[33],而历史上山西地区也有虎分布[34]。而如今虽难觅虎的踪迹,但每一物种在长期的自然选择中都形成了利于自己的最适活动节律[25],故华北豹一直保持着昼间活动。野猪的活动节律在家畜散养期表现为晨昏性,而在圈养期为昼行性,可能是圈养期正值冷季,夜间气温低,为减少不必要的能量散失,这结果与东北虎豹国家公园、秦岭观音山地区的研究结果相近[35-36]。但同时研究也显示野猪在夜间有较高活动性,可能是人为因素干扰引起的改变[31]。研究结果狍为昼间活动,而国内一些结果显示狍为晨昏性[37-38],分析认为可能主要受种间竞争驱使,以生态位分化降低对资源的竞争。如贺兰山两种有蹄类动物在日活动节律的差异实现时间活动的分化,减少竞争[3];植物通过物候期差异达到时间生态位分化实现共存[11]。
狍与野猪在家畜圈养期的夜间活动都有所增加,可能在气候恶劣、食物资源缺乏等不利条件下,野生动物通过加大时间生态位宽度提高适温范围,得以增强资源利用能力和环境适应力[39]。猫科动物活动模式的主要驱动力之一是猎物的可获得性,当猎物丰富时,捕食者和猎物在空间和时间维度上的同步性可以提高捕食概率[40]。故华北豹在家畜圈养期的日活动节律也随猎物在夜间的活动有所增加。
综上所述,放牧作为最主要的人为干扰之一,对同域分布物种在资源上的竞争和干扰将会威胁野生动物的生存。生态位重叠反映了同域分布物种在资源利用上的相似性,重叠程度越大,竞争越激烈。而生态位分化通过减少对时间、空间等资源的竞争,使生态相似的物种得以共存。生态位理论的研究内容不仅为理解野生动物种间关系、了解野生动物种群现状提供依据,并且在群落结构和功能、生物多样性和保护区管理中有重要意义。
致谢:感谢国家林业和草原局猫科动物研究中心、山西铁桥山省级自然保护区和山西华北豹栖息地保护院士工作站给予的帮助。
