中国亚热带-暖温带过渡区锐齿栎林净生态系统碳交换特征
2020-11-13牛晓栋孙鹏森刘晓静栾军伟刘世荣
牛晓栋,孙鹏森,刘晓静,栾军伟,刘世荣,*
1 中国林业科学研究院森林生态环境与保护研究所/国家林业和草原局森林生态环境重点实验室,北京 100091 2 河南宝天曼国家级自然保护区管理局,南阳 474350 3 国际竹藤中心竹藤资源与环境研究所,北京 100102
人类活动引起的大气CO2浓度升高是全球气候变化的主要驱动因素之一[1],森林生态系统在减缓全球变暖方面扮演重要角色,据统计全球森林1年能吸收14.9Pg大气中的CO2,这个量相当于全球化石燃料燃烧碳排放的一半[2]。因而,在区域和全球尺度上精准计算森林的碳吸收能力对于认识森林生态系统在减缓全球气候变化方面发挥的作用和潜力是非常重要的。涡度相关法是长期测算生态系统碳通量最可靠和切实可行的方法[3- 4]。许多国家已经开展了多年的各类型森林生态系统的碳通量的观测,积累了大量翔实可靠的数据,并取得了一些阶段性的成果[3,5]。中科院于2002年建立了中国陆地生态系统通量观测研究网络(Chinaflux),开始对森林水、碳通量进行观测,后来国内部分高校和科研单位也陆续开始采用涡度相关技术开展不同类型森林生态系统的碳通量研究。然而,目前为止在我国暖温带林区的森林碳汇的精准估算还存在较大的不确定性,因为这一区域缺乏长期的森林的通量观测站点。
锐齿栎(Quercusaliena)是暖温带落叶阔叶林的主要建群树种之一,以锐齿栎为优势种的暖温带落叶阔叶林是我国自然地理南北分界秦岭山脉的代表森林类型,在北、中亚热带的湖北、湖南、江苏、四川等省亦广泛分布;在河南伏牛山、太行山海拔1000—2000 m的山地常成纯林。陈存根[6]等报道秦岭锐齿栎林的生产量为14.33 t hm-2a-1,表明秦岭林区锐齿栎具有很高的生产力。然而,这种生物量测定法计算生产力时只考虑了植物的生物量碳,土壤碳的变化并没有进行考虑。森林土壤可能会积累碳[7],也可能会排放碳[8],因此,需要开展锐齿栎林生态系统尺度的碳通量研究,籍以准确评估该区域的森林碳收支。
本研究选取河南内乡宝天曼地区典型土壤森林-锐齿栎林为研究对象,采用国际通用的涡度相关技术对该地区的锐齿栎林的碳通量进行观测研究,旨在探明该地区锐齿栎林的碳交换的日变化和季节变化规律,深入了解在不同时间尺度下锐齿栎的碳吸收/排放变化特征及其环境驱动机制。本研究在一定程度上可填补我国在亚热带-暖温带气候过渡区天然次生林碳交换研究的空白,为进一步准确评估我国森林的碳汇功能区分布提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
本研究在国家林业和草原局河南宝天曼森林生态系统定位研究站进行。该站位于河南省内乡县宝天曼自然保护区(111°47′—112°04′E,33°20′—33°36′N),相对海拔高度600—1800 m,最高峰曼顶海拔1830 m。生态站的水文、土壤和植被具体情况可参见文献[9]。
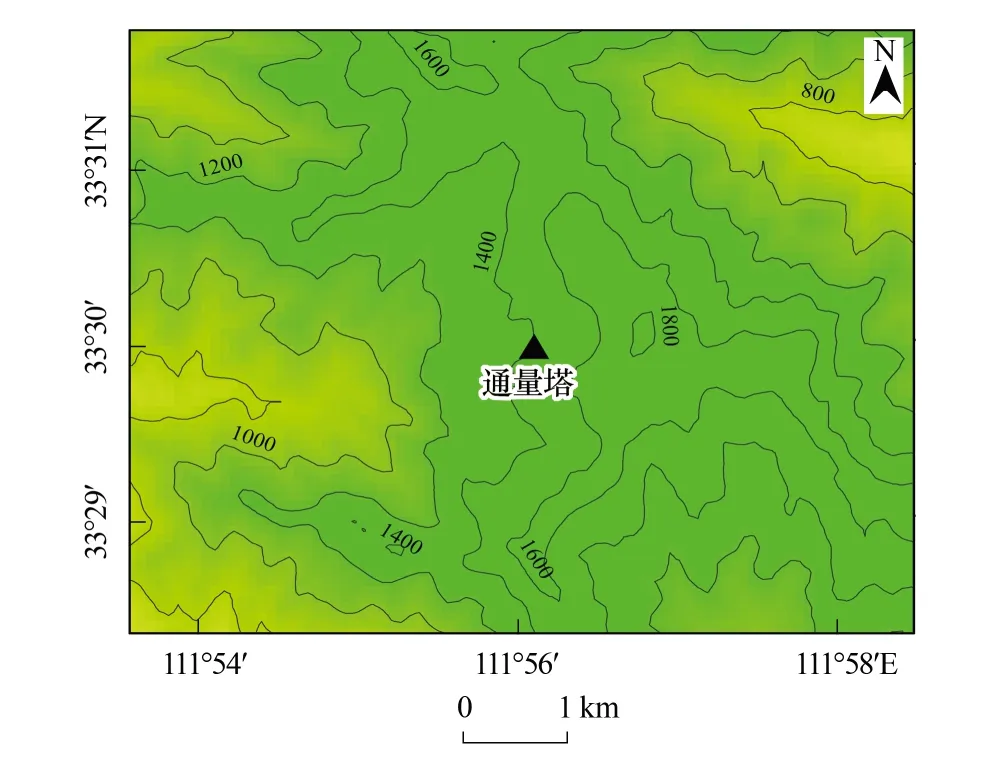
图1 宝天曼森林生态站通量观测点的地形图 Fig.1 The topographical map of flux observation site at Baotianman forest ecosystem research station
通量塔位于海拔1410.7 m(图1),地理坐标33°29′59″N,111°5607″E。观测林分下垫面比较平缓。以通量塔为中心建立1个永久的1hm2样地,样地内优势种为锐齿栎(Quercusaliena),其他伴生乔木有三桠乌药(Lauraceae.obtusiloba)、垂枝条泡花树(Meliosmaflexuosa)、大椴(Tilianobilis)、华榛(Coryluschinensis)等。活立木密度为1314株/hm2。平均乔木树高18 m,胸径19.8±2.8 cm。灌木包括刚毛忍冬(Lonicerahispida)、桦叶荚蒾(Viburnumbetulifolium)、接骨木(Sambucuswilliamsii)、连翘(Forsythiasuspense)、毛花绣线菊(Spiraeadasyantha)和秦岭木姜子(Litseatsinlingensis)等,平均高度3.8 m。
1.2 通量观测
宝天曼通量站开路涡度协方差系统安装于塔29 m(1.5倍林冠高)高处的主风方向位置。涡度系统由红外CO2/H2O气体分析仪(LI- 7500,Li-Cor Inc.,USA)和GILL三维超声风速仪(Gill,UK)组成。湍流脉动信号采样频率为10 Hz,脉动数据通过CR3000(Campbell Scientific Inc.,USA)采集。
在通量塔22 m高处,采用1个光量子传感器(Model LI190SB,Li-cor,Inc.,USA)测量光合有效辐射(Photosynthetic effective radiation)。采用HMP- 45D空气温湿度仪(Vaisala,Finland)测量4层空气温度(Air temperature,Ta)、空气湿度(Air relative humidity,RH),高度分别为2,8,14和22 m。4层土壤温度(Soil temperature,Ts)采用传感器(107,Campbell Scientific Inc.,USA)进行测量,深度分别是5 cm、10 cm、20 cm和40 cm。4层土壤含水量采用1套EasyAG型土壤湿度梯度仪进行测量,深度分别是10 cm、20 cm、30 cm和50 cm。所有常规气象因子测量频率为0.5 Hz,通过CR3000数据采集器(Campbell Scientific Inc.,USA)每30 min自动记录其平均值。
1.3 涡度相关数据处理
采用具有自主知识产权的中国通量网(Chinaflux)通量观测数据处理系统[10]对观测到的30 min的CO2通量数据进行计算。数据处理的步骤包括二次坐标旋转[11]、谱校正、WPL校正[12]、冠层储存通量修正[13]等。本研究中,探头高度以下的大气CO2储存项Fs利用CO2/H2O分析仪测定的CO2浓度进行计算。
净生态系统碳交换(net ecosystem carbon exchange,NEE)由下面的公式计算出:
NEE=Fc+Fs
式中:Fc为CO2湍流通量,由涡度系统观测获得;Fs为涡度相关仪器观测高度以下大气CO2储存通量,可由下式进行估算:
Fs=ΔC/Δt×Δz
式中:ΔC为高度z处前后2次相邻时间测定的CO2浓度差(mg/m3),Δt为前后2次测定的时间间隔,为1800 s,Δz为CO2浓度观测高度[11],在本研究中为29 m。NEE为负时表示锐齿栎林净吸收大气CO2,反之表示净排放CO2。
经过上述计算得到的CO2通量仍存在一些异常值,再对其进行一系列的质量控制,主要包括降水同期数据剔除、阈值剔除(-2—2 mg m-2s-1)、异常数据剔除(某一个数值与连续5点平均值之差的绝对值>5个点方差的2.5倍)以及摩擦风速U*筛选等。本文利用平均值检验法[14]计算得到的摩擦风速阈值为0.11 m/s。将夜间摩擦风速在0.11 m/s以下的数据进行剔除。对上述被去除的数据和由于仪器故障、停电等原因造成的数据用下列方法进行插补:(1)小于2 h的缺失数据用线性内插法插补;(2)大于2 h的缺失数据采用查表法[15]进行插补。具体做法为将温度与光合有效辐射作为主要环境因子,以相邻两月为一个时间段建立查找表。在一个时间段内,将光合有效辐射(Photosynthetically active radiation,PAR)作为主分隔因子,以100 μmolm-2s-1为间隔,在每个间隔内以温度作为次分隔因子,以2 ℃为间隔划分为若干级别,然后分别计算每个间隔内有效NEE平均值。不同月份的缺失值再根据缺失时的气象条件在相应查找表中查找相应有效NEE来插补。
1.4 生态系统呼吸和生态系统总生产力
利用涡度相关法仅能直接获取NEE,为了解生态系统呼吸(Ecosystem respiration,Re)和总生态系统生产力(Gross ecosystem productivity,GEP)的变化,需要将NEE进行拆分。由于夜间(PAR≤3 μmolm-2s-1)生态系统完全处于CO2排放状态,所以夜间生态系统呼吸值(Ecosystem respiration at night,REnight)等于夜间NEE值,而白天(PAR>3 μmol m-2s-1)生态系统呼吸(Ecosystem respiration at daytime,REday)则是依据REnight与土壤温度拟合的呼吸模型再结合白天土壤温度计算出。具体见下式:
Re=REnight+REday
GEE=NEE-Re
GEP=-GEE NEP=-NEE
生长季每个月,30min尺度上PAR与对应的白天(PAR>3 μmol m-2s-1)净生态系统碳交换量(Net ecosystem carbon exchange at daytime,NEEday)的关系可以用直角双曲线模型[16]表示。见下式:
NEEd=Amax×α×PAR/(Amax+α×PAR)-Rd
式中:NEEd为白天净生态系统碳交换量(mg CO2m-2s-1);α为生态系统的初始光能利用效率( μmol CO2μmol-1photon);Rd为生态系统白天的平均生态系统呼吸(mg CO2m-2s-1)。
夜间NEE与土壤5 cm深处的温度用下式[17]拟合:
NEEnight=R0×Q10Ts/10
NEEnight是夜间生态系统CO2交换量,R0是当土壤5 cm温度为0℃时的夜间生态系统呼吸速率,Q10是温度敏感性系数,Ts是深度为5 cm处的土壤温度。
1.5 统计分析
每日的昼夜温差是用每天白天的所有30 min的空气温度的平均值减去每天夜间的所有30 min的空气温度的平均值。采用回归分析对生态系统净碳交换量及其组分与环境变量的关系进行分析。通过残差分析研究白天净生态系统碳交换量NEEd与多个环境因子的关系[18]。30min尺度上,NEEd残差是指实测NEE值与由PAR得出的估算值的差值,正、负残差分别表示生态系统低于或高于由PAR决定的生态系统碳吸收能力。选取主要生长季6—9月白天的数据,以NEEd残差为因变量,温度、饱和水汽压差(Saturation vapor pressure deficit,VPD)和土壤含水量(Soil water content,SWC)为自变量进行相关分析,用Person相关系数及t检验得出的显著性评价不同变量间的相关关系。采用SPSS软件进行统计分析,采用origin软件制图。
本文中空气温度选取22 m高度处的数据,计算每日平均值;土壤温度选取深度为5 cm处的数据,计算每日平均值;降雨量和光合有效辐射计算每日累积值,光合有效辐射高度为22 m;VPD由22 m处空气温湿度根据下式计算得出:
计算每日平均值。
2 结果与分析
2.1 观测期间锐齿栎林中环境因子的季节变化
本研究观测期间锐齿栎林的空气温度(Air temperature,Ta)与土壤温度(Soil temperature,Ts)具有相似的季节变化趋势(图2),其日平均温度最高值均出现在7月27日(Ta=23.7℃,Ts=21.0℃),日平均空气温度最低值和土壤温度最低值分别出现在1月30日(Ta=-6.1℃)和2月12日(Ts=0.5℃),冬季的Ts明显高于Ta,并滞后于Ta。Ta和Ts的日平均变幅分别为-6.7—23.7℃和0.5—21.0℃,观测期间内Ta平均为9.9℃,Ts平均为9.6℃。
观测期间锐齿栎林内最大的日总光合有效辐射(PAR)(22.5 mol m-2d-1)出现在5月16日(DOY=136),月总辐射呈现双峰型(图2),5月和7月的总辐射最高,10月最低。同时受雨雪天气的影响,PAR存在较大的短期波动。
观测期间的总降雨量为1232.8 mm(图2),比多年平均值791.9 mm多出440.9 mm,多出近37.9%,生长季降雨占全年降雨的77.6%,其中9月降水量最高(235 mm),仅9月9日(DOY=252)1 d的降水量就高达73.2 mm。日均VPD的最大值达2.8 kPa出现在4月7日(DOY=97)(图2)。5月平均VPD最高,总体表现为春季高,夏季低。
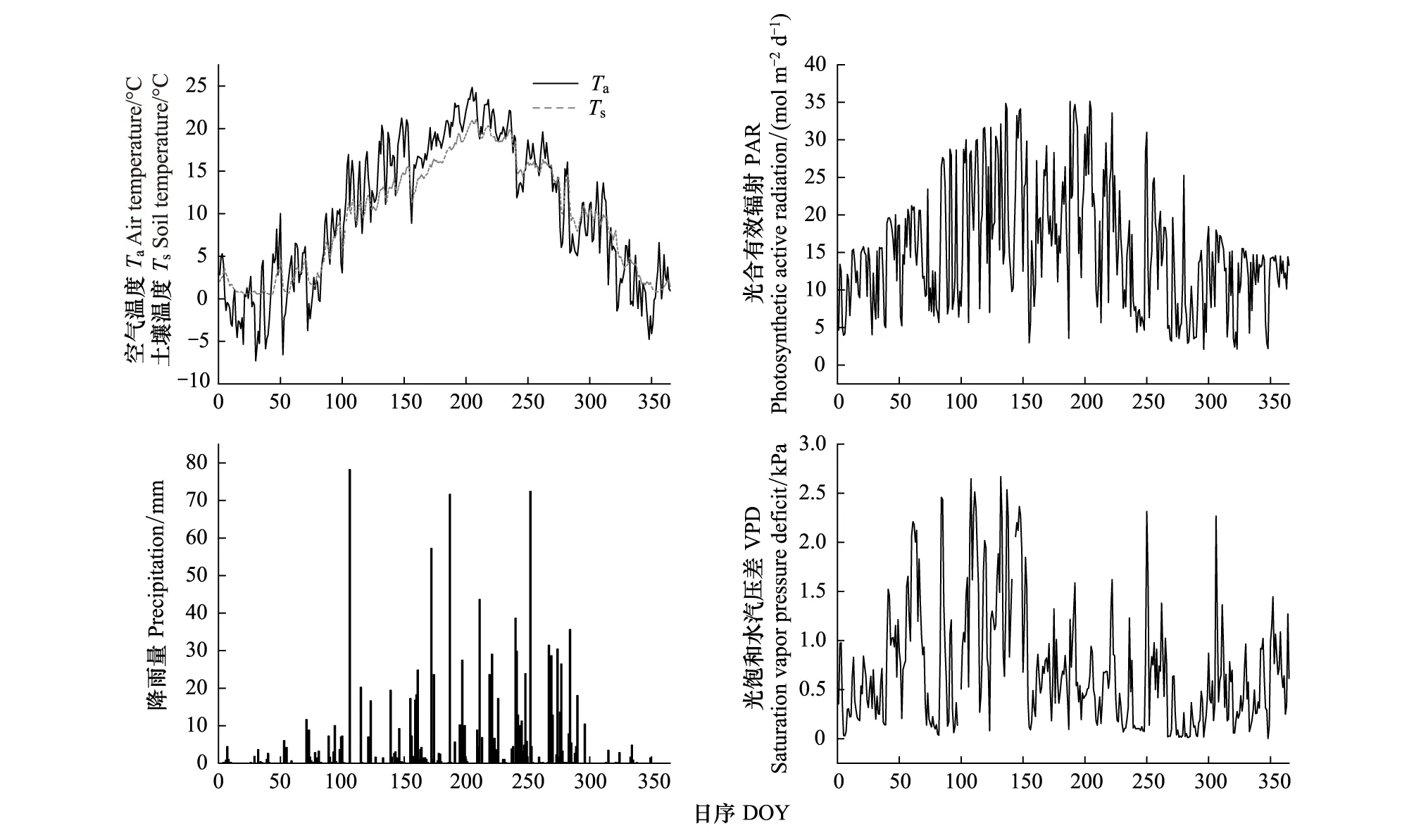
图2 2017年1月—2017年12月宝天曼锐齿栎林中环境因子的季节变化Fig.2 Seasonal variation of environmental factors at the Baotianman oak forest from January 2017 to December 2017Ta: 空气温度 Air temperature; Ts: 土壤温度 Soil temperature; PAR: 光合有效辐射 Photosynthetic active radiation; VPD: 饱和水汽压差 Saturation vapor pressure deficit
2.2 净生态系统碳交换的日变化
研究表明,生长季(5—10月)锐齿栎林NEE具有明显的日变化规律(图3)。日出后,随着空气温度和光合有效辐射的增加,光合作用逐渐增加,锐齿栎林由释放CO2转为吸收CO2,NEE由正变负。至11:30前后,锐齿栎林的CO2吸收量达到最大值。15:00以后,随着光合有效辐射和空气温度的下降,光合作用逐渐变弱,锐齿栎林的CO2吸收量也逐渐减少。日落前后,锐齿栎林由吸收CO2转为释放CO2,NEE由负变正。夜间锐齿栎林的CO2释放量波动较小,并明显小于白天CO2的吸收量。
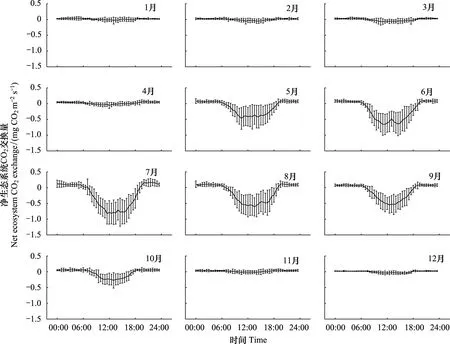
图3 天然锐齿栎林NEE月平均日变化Fig.3 Diurnal variations of monthly mean net ecosystem carbon exchange (NEE) in a natural oak forest
在植物生长季,不同月份锐齿栎林的NEE日变化规律大体类似,但也有不同之处。2017年6月份锐齿栎林有明显的光合“午休”现象,在13:00时NEE降低为-0.48 mg m-2s-1,而最高时为11点时的-0.69 mg m-2s-1;5月和7月的中午14点也有轻微的下降,但很不明显,应该不能称之为常见的光合“午休”现象。2017年月平均日最大CO2吸收量在7月最大,为-0.82 mg m-2s-1,10月最小为-0.28 mg m-2s-1。每日锐齿栎林吸收CO2开始时间以6,7月最早(约6:30),10月最晚(约8:00);每日锐齿栎林吸收CO2的结束时间6—7月在19:00,5和8月在18:30左右,9月和10月分别提前至18点和17点。每日锐齿栎林吸收CO2的持续时间以6,7月最大(13 h),10月最小(9.5 h)。
进入非生长季(11月至翌年4月),随着气温下降和光合有效辐射的降低,大部分锐齿栎的树叶枯黄凋落直至叶子完全掉光。这个阶段生态系统的光合作用和呼吸作用都很微弱,NEE日变化趋势变得十分平缓。但由于少量针叶树种的存在,白天的NEE可以看到有极微弱的碳吸收。
2.3 生态系统碳通量的季节变化
受温度、辐射和降水等因素的影响,锐齿栎林NEE的季节变化较大。2017年日累积NEE变化范围为-7.9—1.6 g C m-2d-1(图4)。4月开始气温回升快,空气温度从3月的3℃迅速增长到10℃。一些林下草本和灌木萌发早,树叶也开始萌发,使得锐齿栎林生态系统呼吸迅速增大,但由于冠层叶面积还很小,光合作用相比较3月份并无显著提高,使得4月份出现了一个碳排放的峰值。5月开始随着气温进一步回升,锐齿栎的叶子开始迅速生长,在一个月内即达到了叶面积的最大值。日平均CO2吸收量在这个月期间也逐步上升,并且在5月31日达到了第一个峰值-7.6 g C m-2d-1。由于5月末—6月初这段时间降雨较少,同较高的气温一起导致了5月24日—6月3日这一期间空气中较高的VPD,使得在5月31日第一次CO2吸收峰值之后有个短期的吸收低值阶段。随后充足的降水使得锐齿栎林在6月5日至8月上旬这一阶段的碳吸收能力保持在较高的水平,并且在7月12日达到了1年中日累积碳吸收最高值-7.9 g C m-2d-1。8月18日之后锐齿栎林吸收碳能力呈明显的下降趋势,并且在8月24日—9月5日有近半个月的持续的低的碳吸收阶段。9月6日之后,锐齿栎林的碳吸收能力又开始出现一个小幅度的持续的回升期,直至10月开始后,随着气温明显下降,树叶开始慢慢变黄,锐齿栎林的碳吸收能力才开始稳步地下降,在10月28日之后开始进入持续地碳排放阶段。8月25日—9月5日这一时期是连续的阴雨天气(图1b),连续阴雨导致的低温和低的光照使得锐齿栎林出现了连续的低的碳吸收(图4)。11月的月平均空气温度低于5℃,土壤5 cm温度也降到了10℃以下,林内锐齿栎的叶子都已落光,只有很少量针叶林存在,加之气温、辐射很低,生态系统的光合作用与呼吸作用都很微弱,总体表现为弱的碳排放。综合分析表明,锐齿栎林在生长季5—10月为碳汇,在非生长季11月至翌年4月为微弱的碳源(图5)。
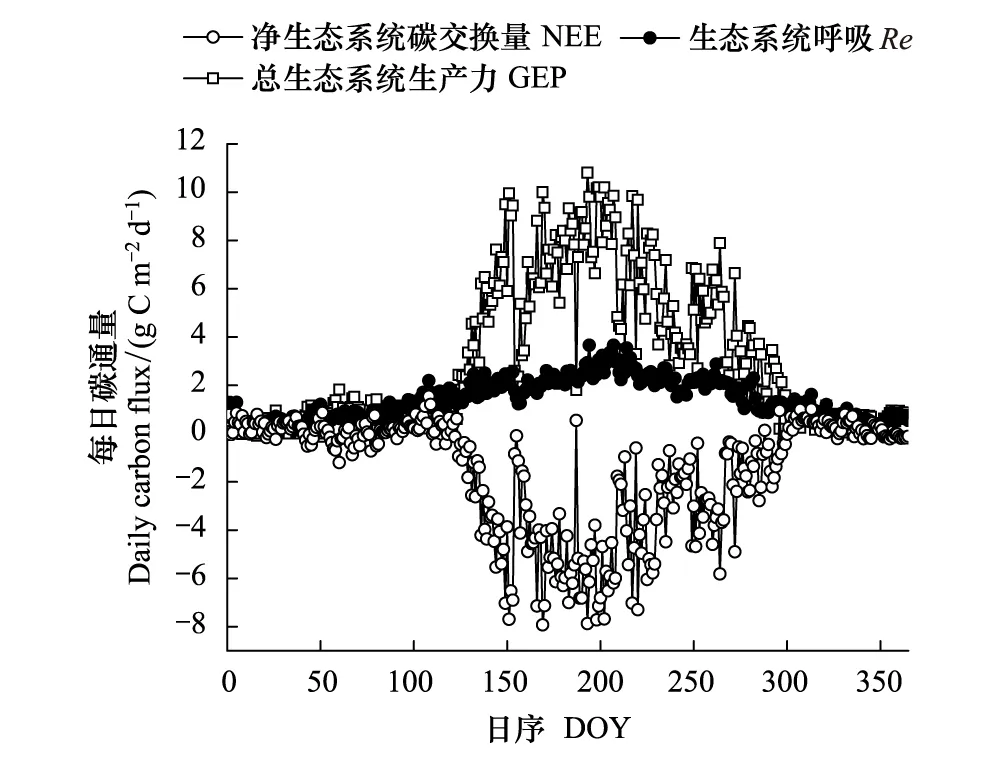
图4 天然锐齿栎林NEE,Re,GEP季节变化(2017年) Fig.4 Seasonal variation of NEE,Re,GEP in a natural oak forest during 2017NEE: 净生态系统碳交换量 Net ecosystem carbon exchange; Re: 生态系统呼吸 Ecosystem respiration; GEP: 总生态系统生产力 Gross ecosystem productivity
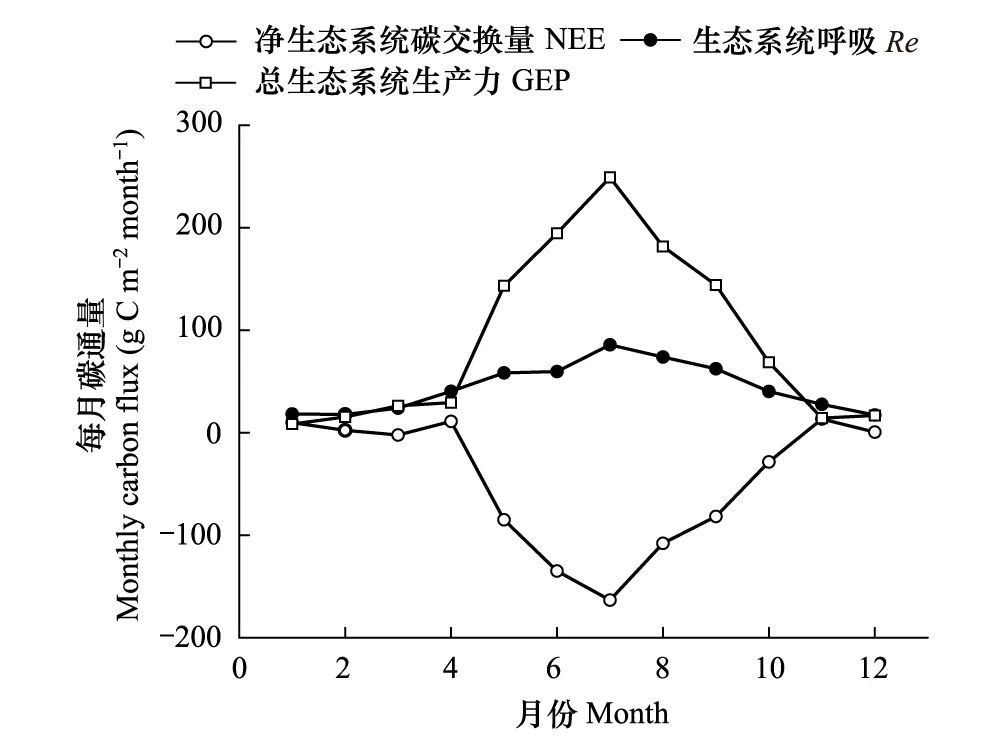
图5 全年各月累积净生态系统CO2交换量(NEE)、生态系统呼吸(Re)和总生态系统生产力(GEP)变化过程Fig.5 Change process of monthly cumulative NEE,Re,GEP
2.4 夜间净碳交换和温度的关系
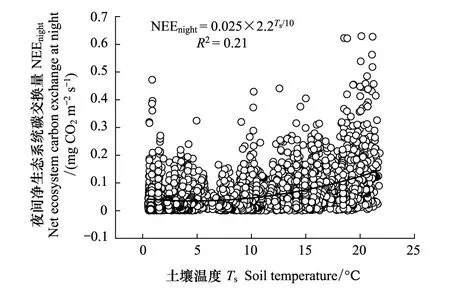
图6 强湍流条件下(U*>0.11)夜间生态系统呼吸(Re)与5 cm深度土壤温度(Ts)的关系Fig.6 Relationship between nighttime ecosystem respiration (Re) and soil temperature measured at 5 cm depth (Ts) under high turbulence(U*>0.11)NEEnight: 夜间净生态系统碳交换量 Net ecosystem carbon exchange at night; Ts: 土壤温度 Soil temperature
30min尺度上5 cm土壤温度能很好地解释夜间锐齿栎林生态系统的净CO2交换量的变异,计算得出生态系统呼吸的温度敏感性(Q10)的值为2.2,R0为0.025 mg CO2m-2s-1(图6)。
2.5 白天净生态系统碳交换与PAR的关系
30min尺度上生长季各月的白天NEE随着PAR的增大而逐渐增强,NEE负值也随之增大,随着生态系统对CO2的吸收量增大的趋势,系统的碳汇能力也逐渐增大,白天最大光合速率和平均白天生态系统呼吸速率均在7月最大(表1),与光合有效辐射表现出相同的季节变化趋势,但当辐射增加到光饱和点(PAR=1000 μmol m-2s-1)(图7)时,生态系统的碳吸收量接近饱和。生长旺盛期6—8月的生态系统初始光能利用效率为0.07—0.09 μmol CO2μmol-1photon,平均值为0.08。最大光合速率在7月份最大为1.47 mg CO2m-2s-1,在10月份最小为0.44 mg CO2m-2s-1,白天呼吸速率最小在10月份,为0.07 mg CO2m-2s-1,最大在7月份为0.19 mg CO2m-2s-1(表1)。
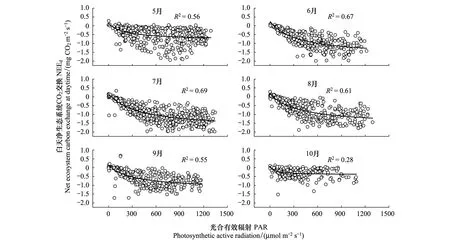
图7 2017年5—10月白天生态系统净碳交换(NEEd)与光合有效辐射(PAR)的关系Fig.7 Relationship between daytime net ecosystem CO2 exchange (NEEd) and PAR from May to October,2017NEEd: 白天净生态系统碳交换量 Net ecosystem carbon exchange at daytime
2.6 环境因子对NEE的影响
6月和9月的NEEd残差与10 cm深度土壤含水量(Soil water content at depths of 10 cm,SWC)极显著相关,但6月是负相关,9月是正相关。其他月份NEEd残差与VPD极显著正相关(表2)。
2.7 昼夜温差及日平均气温对碳通量的影响
图8是生长季(6—9月份)气温日较差和日均温对生长季每日NEE值的影响结果。在生长季,随着昼夜温差的增大,CO2净吸收量增加,从R2来看,相比较日均温,NEE与日较差具有更好的相关性(R2=0.5114)。NEE与日较差二者之间呈二次多项式关系,存在一个阈值为5.03℃。
3 讨论
3.1 锐齿栎林生态系统碳交换的环境影响因子
在夜间,一个简单的指数方程经常被用来模拟净生态系统碳交换量和温度之间的关系[19]。本研究的结果也证实了这一现象,虽然R2方值比较低,但是二者的相关性是极显著的。温度可以选择不同深度的土壤温度或不同高度的空气温度[14]。本研究选取的5cm深处的土壤温度拟合效果最好。由于夜间森林土壤呼吸占生态系统呼吸量的绝大部分[20],而土壤温度是控制土壤呼吸的主要环境因子[9],因此,以土壤温度作为驱动变量更为合理。这与北京八达岭人工林[18]和长白山阔叶红松林[21]的研究结果相似。另一方面,很多研究表明土壤水分对生态系统呼吸也有显著的影响,由于本研究选取的2017年是一个湿润年,降雨量为近几年最高值,因而未探讨土壤水分对生态系统呼吸的影响,下一步我们会重点关注土壤水分的影响。
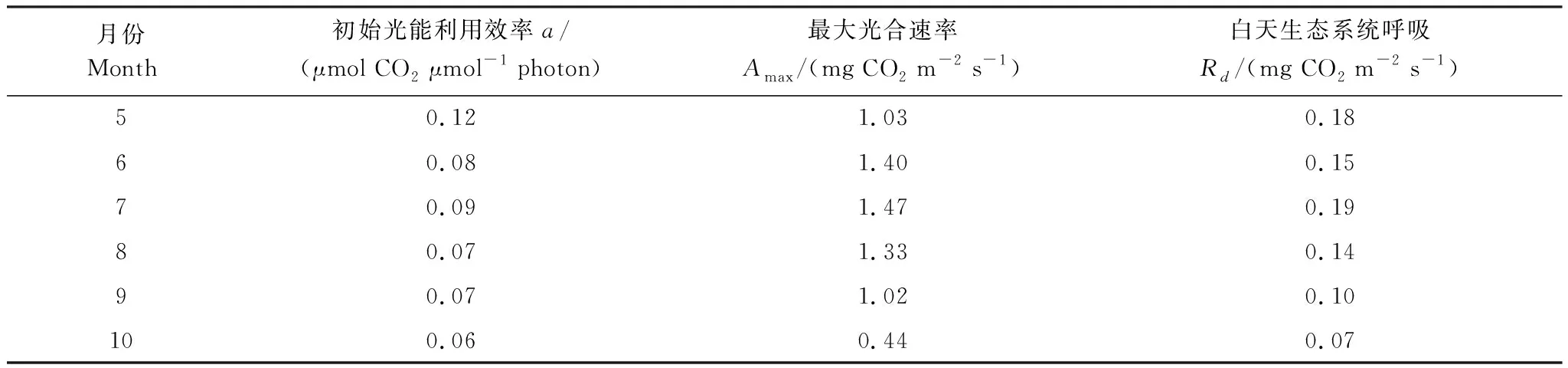
表1 白天生态系统净碳交换(NEE)与光合有效辐射(PAR)之间的相关参数比较
表2 2017年6—9月NEEd残差与主要环境因子在30min尺度的相关性
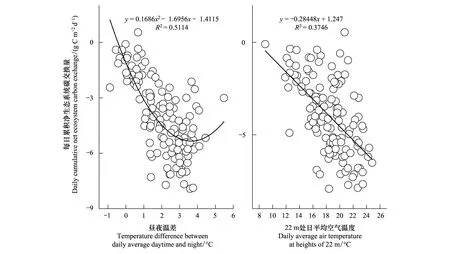
图8 生长季气温日较差和日均温对生态系统净碳交换日总量的影响Fig.8 Effects of daily air temperature difference and daily average temperature on daily amount NEE during growing season
在生长季,白天净生态系统碳交换量除了主要受PAR调控外,其他的环境因子如温度[21]、土壤含水量SWC[22]和VPD[23]也会对白天NEE的值产生影响。在生长季初期,土壤含水量对生态系统的碳通量有重要影响,如同小娟[23]发现黄河小浪底的人工林碳通量的年际变异与全年的降水、土壤含水量和VPD无关,但与春季尤其是5月份的SWC显著相关,春季低的土壤含水量会导致全年的碳吸收量都变低。而在生长季中期,VPD比SWC对净生态系统生产力(Net ecosystem productivity,NEP)的影响更大,如谢静等[24]对北京奥林匹克森林公园人工林的研究表明,夏季(6—9月)的NEP除了与PAR相关性最高之外,VPD的影响最大。唐详[18]在北京八达岭林场的人工林观测也证实了这一点,这与我们的研究结果一致。这可能是因为这一阶段植物的叶面积较大,对空气中的湿度更为敏感,而土壤含水量较低时,根系可以从较深的土壤中吸水[25]而不影响碳吸收。我们也观测到9月份的土壤含水量升高会对碳吸收有抑制作用,这可能是因为2017年9月份整体降水量与往年相比偏高[26],而土壤中含水量变高会对根系呼吸产生抑制,进而影响植物叶片吸收碳的能力。昼夜温差大有利于碳积累这一现象在青藏高原东缘阿柔高寒草甸[27]与海北高寒草甸[28]都有发现,但是这一现象在森林生态系统的研究中还未见报道。本研究发现在宝天曼地区主要生长季期间每日的昼夜温差与每日的累积NEE有明显的负相关关系,但有一个阈值为5.03℃,说明了昼夜温差对本地区碳吸收的重要作用。
3.2 观测期内锐齿栎林生态系统的碳收支
利用白天的30min尺度的NEE和PAR的关系可以计算出每月的初始光能利用效率,最大光合速率等参数。锐齿栎林生长季的月平均初始光能利用效率0.08 μmol CO2μmol-1photon比千烟洲人工林的0.032[29]和黄河小浪底人工林[23]的0.02高,也明显高于Harvard森林[30]的0.044和鼎湖山常绿林[31]的0.034。与长白山阔叶红松林6—8月的0.09[29]和哀牢山老龄林[32]的0.07—0.11接近。表明该锐齿栎林生态系统具有较高的初始光能利用效率,一方面可能是因为拥有短暂生长季的落叶林比常绿林的光合利用效率高[33];另一方面可能因为观测对象所处的高海拔山区多云天气较多,导致散射辐射的比例较大,本研究结果与很多研究者认为的光合利用效率的强度在散射辐射条件中是直射辐射2倍的结论[34]相吻合。
2017年观测点总生态系统生产力、生态系统呼吸和净生态系统生产力的年总量分别为1099.3、529.9和569.4 g C/m2。方精云[35]曾指出与样地实测数据相比较,涡度相关法所获得的NEP值可能偏高。一般来说,使用涡度相关法观测的夜间的CO2通量总体是偏低的[14],使得对Re估算可能偏低,由此导致对碳汇强度的估算可能偏高[31]。中国区域报道的森林生态系统的NEP范围为-256—904 g C m-2a-1[18,32]。本研究得到的569.4 g C/m2在这一范围内,这一值高于纬度相近的黄河小浪底人工林[23]的年碳吸收量(355 g C/m2)。表明宝天曼地区的天然锐齿栎林具有较高的碳吸收能力。研究区夜间温度明显低于白天温度,昼夜温差大,从而有利于植被碳同化作用,同时海拔较高温度较低,尤其夜间和非生长季的低温抑制了土壤微生物和植物的呼吸作用[36];生长季期间充足的水热,林龄又促使生态系统高的碳吸收量。上述原因叠加导致了宝天曼地区的锐齿栎林是一个较大的碳汇。
3.3 通量观测结果的不确定性
本研究利用国际流行的涡度相关法第一次定量计算出了宝天曼锐齿栎林生态系统的年碳吸收量,生态系统呼吸和总生态系统生产力。虽然之前的研究表明本站点的通量数据的能量闭合度为67%,说明观测获取的数据是比较可靠的[37],但仍然需要指出涡度相关法在测量夜间NEE时存在不确定性。王兴昌[38]指出涡度相关法和生物量测定法,箱式法对比,会高估净生态系统生产力25%,低估生态系统呼吸10%,低估总生态系统生产力3%。目前大部分站点比较流行使用摩擦风速阈值剔除法来进行夜间NEE数据的筛选,但一些研究[39- 40]发现在一些站点并不存在摩擦风速的阈值,这种方法仍然会低估生态系统呼吸值,使用夜间最大呼吸法可以更准确地计算出真实的生态系统呼吸量。本文也初步探讨了当摩擦风速阈值为0.15和0.2 m/s时年通量的大小,结果表明当摩擦风速阈值取0.15 m/s时,净生态系统生产力,生态系统呼吸和总生态系统生产力分别为566.4、525.9和1092.3 g C m-2a-1;而当阈值取0.2 m/s时,净生态系统生产力,生态系统呼吸和总生态系统生产力分别为570.1、526.7和1096.8g C m-2a-1。表明对于本站点而言,摩擦风速阈值在0.2 m/s以下对年通量的影响不是很大,但仍然要注意使用恰当的方法确定摩擦风速的阈值[14]。本文计算得出的生态系统呼吸值为529.9 g C m-2a-1,陆海波[41]在宝天曼的另外3块锐齿栎为优势种的样地测得的2014和2015年的土壤呼吸值分别为497±22和603±21 g C m-2a-1,可以看出在本站点使用涡度相关法测定生态系统呼吸时存在低估现象,低估的可能原因是忽略平流作用和储存通量低估,下一步要结合廓线数据使用夜间最大呼吸法[39- 40]更准确地计算夜间生态系统呼吸量。
4 结论
气候过渡带宝天曼地区的天然锐齿栎林在生长季(5—10)月份为碳汇,在非生长季各月为微弱的碳源,净碳释放和碳吸收量分别在4月和7月份达到最大。生态系统呼吸随5 cm土壤温度的升高而增加,Q10值为2.2。该锐齿栎林仍处于快速生长时期,具有较高的碳吸收能力,年碳吸收量达到569.4 g C/m2;研究区生长季辐射强,雨热同期,有利于植物的碳累积,夜间低温和非生长季的低温也抑制了碳排放。在生长季,去除光合有效辐射的影响后,夏季7、8月份,较高的VPD对于锐齿栎林的碳吸收有一定的抑制作用;生长季末期9月份高的土壤含水量对碳吸收也产生了抑制,表明生长末期降水过多对于植物的碳吸收也有负面作用。
