小麦不同绿色器官对灌浆中期高温胁迫的响应
2020-11-13冯波刘开昌高春华曹芳王峥李升东张立顺
冯波,刘开昌,高春华,曹芳,王峥,李升东,张立顺
(1.山东省农业科学院作物研究所,山东济南 250100;2.山东棉花研究中心,山东 济南 250100;3.济南永丰种业有限公司,山东济南 250306)
小麦生育后期发生的高温胁迫因其通常伴随着籽粒灌浆而抑制籽粒产量[1-4]。近年来,我国黄淮海地区每年在小麦灌浆期都发生高温胁迫,小麦产量和品质下降。热胁迫已经成为限制小麦产量的最主要非生物胁迫之一。IPCC第五次会议报告指出,到2100年全球的平均温度将比1980—2000年的平均温度升高1.8~4.0℃,未来热胁迫带来的问题很可能加剧。研究小麦的耐高温机制并找到提高小麦耐高温胁迫性能科学可行的方法,缓解因胁迫造成的减产是当前小麦育种家和栽培学者迫切的科学任务。近些年来,小麦非叶光合器官(如穗、裸露的穗下节间等)因具有高效光合能力而引起人们的研究兴趣,尤其是逆境下叶片因干旱、干热风以及病虫害而衰老,而非叶光合器官的高效光合特性可以弥补叶片的光合不足[5-10],非叶光合器官在高温逆境下还具有光合抗逆优势[8,10,11]。因此,许多科学家认为提高穗、穗下节间等非叶光合器官在小麦群体中的比例,可以提高小麦对干旱和热胁迫的适应能力,从而提高作物的产量潜力[8,12]。
研究发现,逆境下植物通过关闭气孔、渗透调节、活性氧代谢等生理活动进行自我调节以适应环境变化[13]。灌浆期小麦叶片的抗氧化酶活性随着温度升高而降低,裸露的穗下节间具有较高的抗氧化酶活性,从而在生育后期环境温度较高的条件下较叶片衰老慢[9]。张英华等[14]研究表明,高温胁迫初期旗叶、穗的SOD活性和旗叶、旗叶鞘、籽粒的CAT活性以及旗叶、旗叶鞘、颖片的POD活性即诱导增强,而其它器官的抗氧化酶活性则在高温持续一段时间后诱导增强,高温解除后旗叶鞘、穗下节、颖片的SOD活性和旗叶、颖片、籽粒的POD活性有恢复迹象,高温对其它器官的SOD和POD活性以及所有器官的CAT活性造成不可逆影响。NPQ在光能转化与利用平衡关系中发挥着重要作用,以热能的形式无害地耗散吸收的多余光能,是植物不可或缺的光防御机制[15,16]。高温胁迫时,小麦叶片可以通过热耗散调节来缓解光合电子传递链的过度还原,保护光合机构,缓解高温对小麦旗叶的伤害[17,18]。Zhu等[19]研究也指出,超过35℃的高温显著增加水稻植株NPQ,叶黄素循环与热耗散在缓解高温引起的光抑制中起着重要作用。隋娜等[20]研究发现,高温胁迫下番茄反义株系的NPQ和(A+Z)/(V+A+Z)增加幅度较野生型大,说明高温胁迫下转基因植株的光保护系统能够更加有效地耗散过剩激发能,减轻对光系统的伤害程度。
研究高温胁迫对小麦生理特性的影响是小麦抗逆稳产栽培调控技术和品种培育工作的基础。前人关于高温对小麦生理特性的影响,研究对象多为叶片;虽然小麦颖壳、内外稃、芒、籽粒功能各不相同,但将穗作为一个有机整体作为研究对象的甚少。前人研究内容以叶片的抗氧化酶活性较多,而系统研究小麦叶片及穗、穗下节间和旗叶鞘等非叶绿色器官对热胁迫响应差异的报道较少。本试验从抗氧化酶活性、非光化学耗散以及叶黄素循环等多个角度系统研究不同绿色器官对热胁迫响应的生理基础,揭示高温胁迫对不同绿色器官的影响,比较研究不同绿色器官应对高温胁迫的防御差异,以期为小麦抗高温胁迫的栽培技术和育种调控提出理论支撑。
1 材料与方法
1.1 试验设计与田间管理
试验于2017—2019年在山东省农业科学院济南试验田进行。供试小麦品种为济麦22(JM22)和新麦 26(XM26)。小区面积 1.5 m×9 m。基本苗165株/m2,行距25 cm。完全随机区组排列,重复3次。两年度JM22均于10月6日播种,XM26均于10月9日播种,花期一致。花后12~14天通过搭建的塑料大棚高温处理(Str)3天。棚高1.7 m,塑料薄膜厚度70μm,透光率92.7%。大棚两侧距底部30 cm处留空,以保证棚内良好的通风状态。每日8时至18时进行高温处理,处理结束后移除大棚。在试验小区冠层上方10 cm处悬挂温湿度计,每30 min自动记录温湿度。以不搭棚的大田小区作对照(CK)。两年高温处理期间均无降水。两年处理期间的平均温度如图1所示,其中2018年的平均温度与最高温度分别是27.86℃和 35.86℃,2019年分别为33.04℃和39.85℃(图1)。
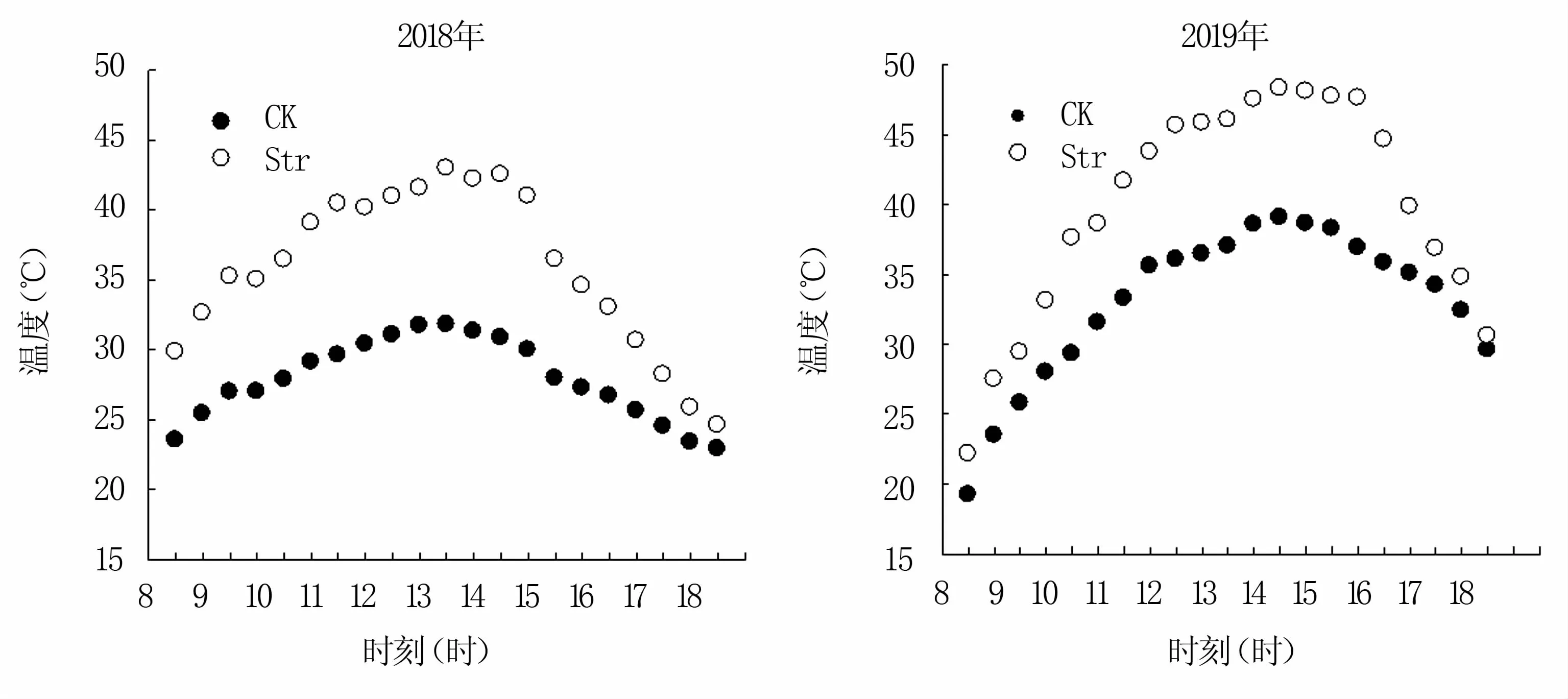
图1 试验期间高温棚内外的气温
1.2 测定项目与方法
1.2.1 样品标记与叶绿素总量测定 开花期对长势一致的单株进行标记,用以测定和取样分析。高温胁迫处理结束后0、5天和10天,取处理和对照标记植株旗叶、裸露的穗下节间、第三小穗以及旗叶鞘各0.5 g,剪成数段放入50 mL提取液(无水乙醇∶丙酮=1∶1)中,25℃黑暗条件下提取24 h,测定 663 nm和 645 nm处的吸光值,按照Arnon[21]的公式计算叶绿素总量。
1.2.2 丙二醛测定 胁迫处理前、结束后0天和5天,参考硫代巴比妥酸法[22]测定丙二醛(MDA)含量。
1.2.3 质膜透性测定 参照杨根平等[23]的方法。胁迫结束后0天,处理植株和对照每份材料均取15个直径0.5 cm的叶圆片,浸于试管中抽真空10 min,28℃保温30 min,其间摇动几次,测定电导率E0,此值作为背景值;然后28℃保温30 min,测定电导率(Ep);最后将试管置于沸水中15 min,冷却至室温,再测1次总电导率(Et)。
细胞质膜相对透性(%)=(Ep-E0)/(Et-E0)×100。
1.2.4 抗氧化酶活性测定 胁迫处理结束后0天和5天,取处理和对照标记植株的穗、穗下节间、旗叶鞘、旗叶,用液氮速冻后置于-80℃超低温冰箱中保存。称取各不同绿色器官鲜样(穗取基部第三和第四小穗)0.5 g,剪碎放入预冷的研钵中,加入磷酸缓冲液1 mL,冰浴下研磨,加缓冲液定容至终体积5 mL,再4℃条件下10 000 r/min离心20 min,取上清液用于酶活性测定。超氧化物歧化酶(SOD)、过氧化物酶(POD)活性分别采用氮蓝四唑光化学还原法和愈创木酚法测定[24],过氧化氢酶(CAT)活性采用紫外线吸收法测定[22]。所有酶活性单位以样品鲜重为基准。
1.2.5 NPQ及过剩光能测定及计算 处理结束后0天,挖取处理和对照的标记植株,快速带回实验室后,分取旗叶、穗、穗下节间以及旗叶鞘,并放置于湿润的吸水纸上,参照Nedbal等[25]的方法,避光暗适应30 min后,采用开放式荧光成像系统(FluorCamFC800,Photon System Instruments Ltd,Brno,Czech Republic),于红光(690 nm)激发条件下分别测定不同绿色光合器官的叶绿素荧光感应曲线(Kautsky曲线)。叶绿素荧光发射瞬间变化由顶部CCD相机捕获,同时记录荧光生理参数NPQ和Qp。过剩光能=(1-Qp)/NPQ。
1.2.6 叶黄素循环组分测定 取胁迫结束0天处理和对照的不同绿色器官,按赵世杰等[26]的方法,于暗中色谱纯丙酮研磨提取,0.22μm微孔滤膜过滤。采用美国Waters公司生产的控制仪、W510型高压液相泵、2487型紫外检测仪,浙江大学智能信息工程研究所研制的N2000系统工作站,测定叶黄素循环各组分含量。组分物质之间的转化用(A+Z)/(A+Z+V)百分比表示。
1.3 数据处理
两年度试验结果趋势一致,采用其数据平均值进行分析。用Microsoft Excel 2010进行数据处理,DPS 17.10软件进行统计分析,单因素方差分析法(one-way ANOVA)和 Duncan’s法进行方差分析和多重比较,SigmaPlot 10.0软件作图。
2 结果与分析
2.1 高温胁迫对小麦不同绿色器官叶绿素总量下降百分比的影响
由表1可以看出,高温胁迫后,小麦不同绿色器官的叶绿素含量均较对照降低,但不同绿色器官间叶绿素总量降幅不一。胁迫结束0天,穗部叶绿素含量下降百分比最大,极显著大于旗叶,旗叶极显著大于穗下节和旗叶鞘,穗下节间和旗叶鞘间无显著差异。两个品种该指标趋势表现一致,但XM26旗叶、穗、穗下节间和旗叶鞘的叶绿素含量下降程度都大于JM22。
胁迫结束后5天,两个品种仍是穗部叶绿素含量下降百分比最大,极显著大于其它绿色器官;其次是旗叶鞘,极显著大于旗叶和穗下节间。两品种略有不同的是,JM22各绿色器官中叶绿素含量下降百分比最低的是穗下节间,而XM26则是旗叶,并且旗叶叶绿素下降百分比XM26高于JM22。
胁迫结束后10天,两个品种叶绿素含量较对照下降百分比最大的为旗叶,极显著大于其余绿色器官;其次是旗叶鞘,再次是穗和穗下节间。JM22的穗下节间叶绿素下降百分比极显著低于穗,而XM26两者间无显著差异。
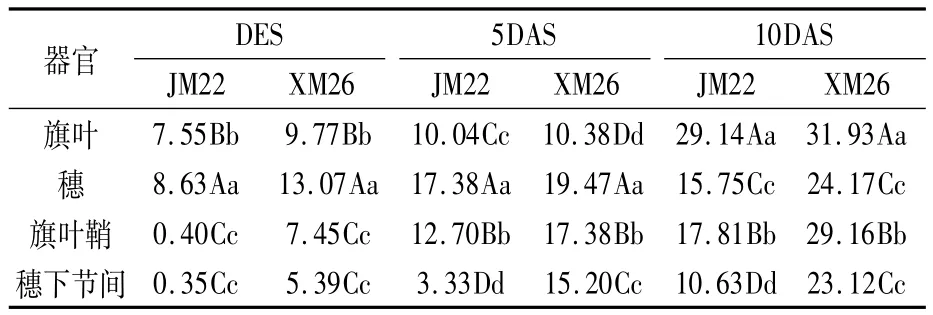
表1 高温胁迫后不同绿色器官叶绿素总量较对照下降的百分比 (%)
2.2 高温胁迫对小麦不同绿色器官丙二醛含量的影响
图2表明,高温胁迫开始发生到胁迫结束后5天,旗叶、穗、旗叶鞘及穗下节间的MDA含量都呈逐渐增加趋势,且高温胁迫加速MDA的积累,两品种表现一致。胁迫结束0天,两品种MDA含量由高到低均依次为旗叶>穗>穗下节间>旗叶鞘。JM22和XM26旗叶、穗、穗下节间和旗叶鞘MDA含量分别比对照增加19.58%和27.22%、19.11%和 15.17%、9.61%和 12.39%、9.89%和14.98%。
胁迫结束后5天,两品种不同绿色器官处理与对照MDA含量由高到低均依次为穗>旗叶>旗叶鞘>穗下节间,JM22和XM26旗叶、穗、穗下节间和旗叶鞘MDA含量分别比对照增加28.82%和 31.42%、26.38% 和 29.13%、20.61%和19.70%、17.83%和 18.81%。

图2 高温胁迫对小麦不同绿色器官MDA含量的影响
2.3 高温胁迫对小麦不同绿色器官质膜透性的影响
由表2可知,高温胁迫增大小麦不同绿色器官的电导率,各绿色器官质膜透性增大。与对照相比,质膜透性增加百分比最大的是旗叶,其次是穗的外稃,变化最小的是穗下节间,旗叶鞘、护颖及芒居于中间。两品种变化趋势一致。质膜透性的增加使得细胞内源物质外渗程度加大,但XM26处理之前和处理之后的各个绿色器官质膜透性均大于JM22对应各绿色器官,不仅如此,胁迫后其各绿色器官电导率增加百分比也都大于JM22,说明热胁迫对XM26各绿色器官质膜透性的影响大于对JM22各绿色器官的影响。

表2 高温胁迫对小麦不同绿色器官细胞质膜透性的影响
2.4 高温胁迫对小麦不同绿色器官SOD活性的影响
图3显示,高温胁迫结束0天,JM22旗叶SOD活性较对照升高15.55%,XM26较对照升高22.60%;穗部SOD活性降低,JM22较对照降低12.67%,XM26降低 23.00%;JM22穗下节间SOD活性比对照降低1.79%,XM26降低7.95%;JM22旗叶鞘的 SOD活性较对照降低0.70%,XM26降低1.90%。
胁迫结束后5天,各绿色器官SOD活性继续降低。胁迫处理旗叶的SOD活性显著低于对照,JM22降低58.24%,XM26降低32.27%;JM22穗部SOD活性比对照降低11.08%,XM26降低23.28%;穗下节间SOD活性显著低于各自对照,JM22降低43.23%,XM26降低59.90%;旗叶鞘SOD活性显著低于对照,JM22减少69.08%,XM26减少71.03%。高温胁迫发生前至胁迫结束后0天和5天,两品种旗叶、穗、穗下节间以及旗叶鞘的SOD活性变化表现一致。
2.5 高温胁迫对小麦不同绿色器官POD活性的影响
自高温胁迫发生到胁迫结束后第5天,旗叶、穗、穗下节间和旗叶鞘的POD活性加速下降,各器官POD活性都显著低于各自对照(图4)。胁迫结束后0天,JM22和XM26旗叶、穗、穗下节间和旗叶鞘的POD活性分别较对照降低21.41%和28.73%、34.97%和 47.04%、36.17%和 46.89%、40.29%和50.25%。胁迫结束后5天,两品种旗叶、穗、穗下节间和旗叶鞘的POD活性比各自对照分别降低 46.45%和 66.67%、42.63%和68.60%、32.66% 和 54.12%、40.34% 和51.53%。

图3 高温胁迫对小麦不同绿色器官SOD活性的影响

图4 高温胁迫对小麦不同绿色器官POD活性的影响
2.6 高温胁迫对小麦不同绿色器官CAT活性的影响
由图5可知,高温胁迫结束后0天,JM22旗叶CAT活性比对照增加13.00%,XM26增加34.40%;而JM22穗、旗叶鞘和穗下节间的 CAT活性分别较对照降低 30.09%、1.75%、16.20%,XM26较对照分别降低 58.96%、37.76%、40.06%。
胁迫结束后第5天,各绿色器官处理的CAT活性显著低于对照,JM22和XM26旗叶、穗、旗叶鞘、穗下节间分别较对照降低 65.77%和81.04%、44.82%和 56.36%、30.67%和 40.37%以及 43.96%和 57.68%。
2.7 高温胁迫对小麦不同绿色器官NPQ和过剩光能的影响
如图6所示,不同绿色器官耗散非光化学能量(NPQ)的能力不同。高温胁迫结束后0天,各绿色器官的NPQ值较对照都升高,JM22旗叶、穗、穗下节间和旗叶鞘的NPQ分别较对照增加2.94%、34.38%、34.38%和 11.76%,XM26相应增加 10.34%、20.00%、16.67%和 14.29%。
胁迫结束后5天,JM22旗叶、穗、穗下节间和旗叶鞘的 NPQ值分别较对照减少 9.38%、15.39%、30.56%和 27.59%,XM26相应减少11.54%、19.36%、38.24%和 11.54%。

图5 高温胁迫对小麦不同绿色器官CAT活性的影响

图6 高温胁迫对小麦不同绿色器官NPQ的影响
高温胁迫结束后0天,两品种不同绿色器官的过剩光能与各自对照相比均显著降低,较对照减少百分比由大到小依次为穗>旗叶>穗下节间>旗叶鞘,JM22和XM26穗、旗叶、穗下节间、旗叶鞘过剩光能减少百分比分别为43.76%和47.27%、39.09%和 40.88%、34.30%和 38.76%、25.58%和28.17%(图7)。

图7 高温胁迫对小麦不同绿色器官过剩光能的影响
2.8 高温胁迫后冬小麦不同绿色器官叶黄素循环的变化
图8表明,高温胁迫条件下,耐热性不同的小麦品种JM22和XM26旗叶、穗、穗下节间和旗叶鞘的脱环氧化状态(A+Z)/(A+Z+V)的变化趋势一致,各绿色器官的(A+Z)/(A+Z+V)都显著增加,穗下节间增加最多,JM22和XM26分别增加19.80%和21.02%;其次是旗叶鞘,两品种分别增加18.37%和12.89%;再次是穗,分别增加16.95%和7.14%;旗叶变化最小,分别增加6.24%和 2.32%。

图8 高温胁迫对各绿色器官(A+Z)/(A+Z+V)的影响
3 讨论
3.1 高温胁迫对小麦不同绿色器官叶绿素含量的影响
作物叶绿素含量影响其光合作用的强度,持绿时间决定光合作用的持续期[27,28]。灌浆期高温胁迫后,小麦不同绿色器官黄化失绿是小麦衰老的形态表现,穗、旗叶、穗下节间和旗叶鞘的叶绿素含量均较对照降低,但各自降幅不同。胁迫结束0天,与对照相比,叶绿素含量下降百分比由大到小依次为穗、旗叶、旗叶鞘和穗下节间,穗下节间和旗叶鞘间无显著差异。两个品种表现一致。
胁迫结束后5天,JM22不同绿色器官叶绿素含量下降百分比由大到小依次为穗、旗叶鞘、旗叶和穗下节间,XM26为穗、旗叶鞘、穗下节间和旗叶。
胁迫结束后10天,不同绿色器官叶绿素含量下降百分比由大到小依次为旗叶、旗叶鞘、穗和穗下节间。旗叶的叶绿素含量较对照下降百分比最大,可能是因为此时对照旗叶的叶绿素含量最高,而冠层其它绿色器官各自对照的叶绿素含量较低。
小麦穗、旗叶、穗下节间和旗叶鞘是构成小麦冠层的主体,也是小麦进行光合作用的主体。高温后小麦不同绿色器官叶绿素含量的下降是造成小麦产量降低的直接因素。因此,如何通过栽培或者化控技术保持高温胁迫后冠层不同绿色器官叶绿素含量是小麦抗逆栽培的一个重要研究方向,也是后续开展研究的重点之一。
3.2 高温胁迫对小麦不同绿色器官MDA含量和质膜透性的影响
通常用丙二醛(MDA)含量来衡量逆境引起的膜脂过氧化程度[29]。高温会损伤细胞膜,引起离子渗漏,使细胞代谢和功能发生紊乱,这些结果的出现与活性氧积累和发生膜脂过氧化有关[30]。Cao等[31]研究高温对不同耐热性水稻的影响发现,高温胁迫显著增加耐热品种叶片的抗氧化酶活性,对热敏感品种影响不大;高温处理增加耐热和不耐热品种的丙二醛含量。
本试验结果表明,高温胁迫加速小麦各绿色器官MDA含量的积累。胁迫结束0天,两品种都表现为旗叶的 MDA含量增加较多,分别为19.58%和 27.22%;其次为穗,分别为 19.11%和15.17%;旗叶鞘和穗下节间增加相对较少,穗下节间为 9.61%和 12.39%,旗叶为 9.89%%和14.98%。胁迫结束后5天,穗下节间和穗的MDA含量较胁迫处理前分别增加92.44%和89.22%,旗叶鞘和旗叶则分别增加41.99%和29.62%。此时,由于环境气温较高,各对照绿色器官的MDA含量增加,处理较对照增加显著。
后期高温使小麦叶片造成质膜相对透性增大[31,32]。本试验表明,高温胁迫改变了小麦不同绿色器官的电导率。质膜透性增加百分比最大的是旗叶,其次是外稃,变化最小的是穗下节间,旗叶鞘、护颖以及芒居于中间。两个品种变化趋势一致。质膜透性的增加使得细胞内源物质外渗程度加大,但XM26处理前后各绿色器官的质膜透性均大于JM22,不仅如此,胁迫后其各绿色器官电导率增加百分比也都大于JM22,说明热胁迫对XM26各绿色器官质膜透性的影响大于对JM22各绿色器官的影响。胁迫结束0天和5天,XM26旗叶、穗下节间、旗叶鞘和穗部的MDA含量较对照增幅基本高于JM22对应各器官的增幅,也高于同期 JM22对应各器官的 MDA含量,说明XM26耐热性差、衰老快,在同等高温条件及高温持续期胁迫下膜脂过氧化程度更加严重。
3.3 高温胁迫对小麦不同绿色器官活性氧清除酶系统的影响
正常条件下,活性氧(ROS)的产生和清除保持平衡,植物细胞中没有活性氧的净积累。逆境下这种平衡会被破坏,造成活性氧的积累。而SOD、POD和CAT对防止膜脂过氧化、减轻逆境造成的膜伤害和延缓植物衰老有重要作用。张黎萍等[33]研究得出高温使小麦旗叶MDA含量升高,SOD活性下降,植株衰老加速。姜春明等[32]认为灌浆早期的高温处理使旗叶内活性氧防卫系统能更有效的启动,而灌浆中期的高温处理SOD活性未见上升,甚至有所下降,CAT活性虽然高于对照,却未能有效减轻膜脂过氧化程度。刘萍等[34]研究认为,高温胁迫可诱导活性氧清除酶系统(SOD、POD和CAT)活性下降,植株体内保护酶系统趋于衰弱或崩溃,高温导致膜脂过氧化程度(MDA)加剧,衰老速度加快。高温胁迫条件下,随着温度的升高,SOD、POD和CAT活性呈下降趋势。
本研究发现,灌浆中期遭受3天高温胁迫后,旗叶SOD和CAT活性都显著升高,而穗、穗下节间和旗叶鞘的SOD活性和CAT活性降低。高温胁迫使两品种旗叶、穗、穗下节间和旗叶鞘的POD活性不论是在胁迫结束后0天还是胁迫结束后5天都显著低于对照,POD清除H2O2的能力大大降低。胁迫结束后0天,XM26旗叶、穗、穗下节间和旗叶鞘的POD活性低于JM22,各绿色器官POD活性较对照下降的百分比也高于JM22。通过比对前人的研究结果,我们认为高温胁迫对活性氧清除酶系统的影响受多方因素的影响。酶促系统启动还是活性功能下降不仅受高温发生的时期、温度条件、高温持续时间的影响,也因绿色器官的不同、品种不同等而存在差异。
3.4 高温胁迫对不同绿色器官叶黄素循环的影响
叶黄素循环在多种自然条件下都有保护光合机构正常运转的功能。叶黄素循环的脱环氧化状态(A+Z)/(A+Z+V)与热耗散能力直接相关。植物遭受胁迫时,光合机构通过叶黄素循环处理过剩光能的能力增强[35]。吴长艾等[36]研究表明,依赖叶黄素循环的热耗散和PSⅡ反应中心的可逆失活在浸水处理过程中对小麦叶片光合机构避免强光高温的损伤方面起保护作用。小麦在开花灌浆期经常会遭受高温强光胁迫,过剩的激发能可能对小麦叶片造成伤害[36,37]。我们通过研究发现,高温胁迫后小麦不同绿色器官依赖叶黄素的热耗散能力均增强。通过比较不同绿色器官叶黄素循环(A+Z)/(A+Z+V)的变化可以看出,依赖叶黄素循环的热耗散能力依次为穗下节间>旗叶鞘>穗>旗叶。这说明高温胁迫条件下,小麦不同绿色器官的热耗散能力均增强。品种间比较发现,JM22依赖叶黄素循环的热耗散能力大于XM26。
3.5 高温胁迫下不同绿色器官的非光化学耗散机制
NPQ,非光化学淬灭,是指因非光化学反应引起的荧光产量的降低,一般用来表示热耗散的多少。较强的热耗散能力可以促进过剩光能以热能形式耗散,降低光合系统内激发能压力,减少活性氧的产生和光抑制的发生频率,是重要的光合防御机制之一。热胁迫下能量耗散可以减少激发能在供体侧的积累,从而对PSⅡ具有较强的保护能力[38,39]。番茄幼苗叶片在一定高温胁迫后并没有诱导过量的活性氧(ROS)产生,这与非光化学淬灭(NPQ)有效淬灭过剩光能有关,而在40℃高温胁迫下番茄幼苗叶片NPQ显著降低,导致叶片中过剩光能(1-qP)/NPQ和 ROS大量积累[40]。李传宗等[41]在棉花上的研究表明,高温胁迫后棉花叶片通过NPQ增高提高其耐热性。本试验表明,不同的非叶光合器官耗散非光化学能量的能力不同,以穗和穗下节间的耗散能力较强,而旗叶和旗叶鞘的热耗散能力较弱。高温胁迫使各器官的NPQ都显著升高,说明在籽粒灌浆过程中随着光合器官的衰老和光合能力的减弱,光合器官俘获的过剩光能增多,而以热的形式耗散掉的光能在逐渐增加,从而减少过剩光能对光合器官的伤害。高温胁迫后各绿色器官的过剩光能与对照相比都显著降低,降幅由多到少依次为穗>旗叶>穗下节间>旗叶鞘。过剩光能较对照显著减少的原因有二,一是植物自身应对胁迫的保护机制,即吸收的光能减少;二是植物对吸收光能的利用相对增多,从而减少过剩光能对植物的损伤。
综上所述,小麦不同绿色器官对高温胁迫的响应不同,不同绿色器官通过有效启动活性氧清除酶促系统、增加非光化学耗散能力、通过叶黄素循环散热等途径进行避热耐热自我调节。旗叶中清除活性氧保护性酶活性显著升高,而穗、穗下节间和旗叶鞘等非叶光合器官的保护性酶活性则因高温而降低;不同绿色器官非光化学热耗散能力则以穗和穗下节间最强,旗叶和旗叶鞘较弱;依赖叶黄素循环的热耗散能力增加幅度依次为穗下节间>旗叶鞘>穗>旗叶。
4 结论
4.1 不同绿色器官对高温防御的生理机制存在差异。高温后旗叶中清除活性氧保护性酶活性显著升高,而穗、穗下节间和旗叶鞘的保护性酶活性则因高温而降低;非叶光合器官通过非光化学耗散和依赖叶黄素循环的热耗散能力要优于旗叶。
4.2 不同基因型品种耐热性不同。JM22光合性能受高温胁迫的抑制较XM26小;JM22在高温下能够更加有效地启动保护酶系来应对高温热害造成的活性氧积累和膜脂过氧化,并且其通过叶黄素循环和非光化学耗散过剩光能的能力也优于XM26。
