增温和刈割对高寒草甸土壤呼吸及其组分的影响
2020-11-12牛书丽常文静
蒙 程,牛书丽,常文静,全 权,曾 辉,*
1 北京大学深圳研究生院城市规划与设计学院, 深圳 518055 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 3 中国科学院大学资源与环境学院, 北京 100049
自工业革命以来,大气温室气体升高引起的全球变暖使得全球平均气温上升了约0.85 ℃,预计到21世纪末全球平均气温将升高1.5 ℃[1]。青藏高原是气候变化的敏感区域,预计变暖幅度远大于全球平均值[2-3]。越来越多的证据表明气候变暖可以改变高寒草地的群落结构和功能[4-5],而这些变暖引起的生态系统过程的变化将会显著地影响高寒生态系统碳通量[6-8]。刈割和放牧作为草地的主要利用方式,可以影响土壤小气候、植物生长和物种多样性等,并进一步影响生态系统碳循环[9-10]。高寒草地储藏着丰富的土壤有机碳,目前正在经历着快速的气候变暖和土地利用变化(如刈割和放牧)[11-12]。增温和刈割可能会影响根系呼吸和微生物呼吸,从而影响生态系统碳循环。目前,有关高寒草甸土壤呼吸对气候变暖和刈割响应的研究仍然有限,因而进行增温和刈割对土壤呼吸及其组分影响的研究,可以为有效应对未来气候变化背景下的生态系统管理提供科学依据。
土壤呼吸是指未扰动土壤中产生二氧化碳(CO2)的所有代谢作用,主要包括自养呼吸(根呼吸和根际微生物呼吸)和异养呼吸(土壤微生物和动物呼吸)两个组分[13-14]。土壤呼吸作为陆地生态系统和大气中规模仅次于植物光合作用的碳通量过程,是全球碳循环和气候变化的重要调控因素[9, 14]。它主要受土壤温度[15-17]、含水量[18-20]、养分有效性[21-22]等非生物因子和植物群落结构[22-23]、植物根系[24]、土壤微生物[25]等生物因子的共同影响。大量研究探讨了增温和刈割对土壤呼吸的影响,但结论仍然存在争议[9, 26]。整合分析结果表明,增温能够刺激土壤呼吸,尤其是在高寒地区[27]。基于单个站点的研究则发现增温可以导致土壤呼吸的持续增加[9, 23, 28],或者初期增加,然后在几年后减弱[17,29-31]。这些响应可归因于生物和非生物因素之间的相互作用,包括微生物碳利用效率[32]、底物供应[33]、温度和土壤水的可用性变化[29-30]等。增温引起土壤呼吸的升高并不一定意味着土壤有机碳(SOC)分解与大气二氧化碳浓度之间存在正反馈,因为土壤呼吸的增加可能是由于净初级生产力增加导致自养呼吸大幅度增加,使得更多的碳(C)分配到根[34]。因此单独测量土壤呼吸并不能为评估土壤在气候变暖下是碳源或碳汇提供确凿的证据[35],同时测量土壤呼吸和异养呼吸对于更好地理解气候变暖下土壤碳排放动态显得尤为必要。刈割作为草原的利用和管理方式之一,可以减少植被覆盖,进而影响土壤表面的蒸发和能量平衡,导致土壤温度升高和加剧土壤水分减少[36]。一些研究发现由于C基质供应减少,即使土壤温度升高,刈割也会使土壤呼吸减少19%—49%[21, 37]。Jia等[38]和Han[39]等人发现土壤呼吸对刈割没有响应是因为刈割未影响植物生长和土壤水分。另一些研究则发现刈割可以通过提高土壤温度[40]、改变地上植物群落结构[41]或刺激丛枝菌根真菌[42]来增强土壤呼吸。因此,刈割对土壤呼吸的影响仍存在争议,其与增温的交互作用研究更是缺乏。
本研究利用红外线辐射加热器(Infrared heater)模拟增温,研究增温和刈割对土壤呼吸和异养呼吸的影响。探讨全球气候变化背景下,模拟增温和刈割对高寒草甸土壤呼吸、自养呼吸和异养呼吸的影响及其与土壤温度、湿度变化之间的关系,以期为青藏高原高寒草甸生态系统的可持续管理提供基础数据和理论基础。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东南缘的四川省阿坝州红原县(32°48′N,102°33′E),海拔大约3500 m。该地区的气候类型属于大陆性高原温带季风气候,年平均气温1.1 ℃,最高气温11.1 ℃,最低气温-9.7 ℃;全年日照时数2000—2400 h;年平均降水量753 mm,降水主要集中在5—9月,占年降水量的80%。植被类型为高寒草甸草原,主要优势种包括:发草(Deschampsiacaespitosa)、溚草(Koeleriacristata)、华丽龙胆(Gentianasino-ornata)、鹅绒委陵菜(Potentillaanserine)、小花草玉梅(Anemonerivularis)。土壤类型为亚高寒草甸土。
1.2 试验设计
试验样地所在区域地势平坦,植物物种分布均匀。试验采用双因素裂区设计,主区为增温方式,亚区为刈割方式,共设置3 m×2 m的主区10个,主区随机设置为对照和增温处理,每个处理设置5个重复;每个主区又被裂分为两个1.5 m×2 m的亚区,其中一个亚区刈割,另一个亚区不刈割,共形成20个试验小区。相邻主区间隔3 m。整个试验共设置4个处理,分别为对照(CK),增温/无刈割(W),无增温/刈割(CL),增温/刈割(WCL),每种处理设置5个重复。
增温样地从2014年6月开始采用红外线辐射器(MSR-2420, Kalglo Electronics Inc., Bethlehem, Pennsylvania, USA)进行全年连续加热,辐射器长165 cm,宽15 cm,悬挂在样地中央,悬挂高度为1.5 m,辐射器输出功率为2000 W,对照样地悬挂相同尺寸的模拟辐射器来模拟增温处理样地所悬挂加热器带来的遮蔽效果。在每年的生长季末期进行一次刈割,留茬高度为1 cm。
1.3 研究方法
1.3.1土壤呼吸和异养呼吸的测定
试验设置前期(2014年初)在每个小区内永久埋入两种类型的PVC土壤呼吸环。其中一个土壤呼吸环(内径10.5 cm,高5 cm)埋入土中2—3 cm,用于总土壤呼吸的测定。将另外一个高度为40 cm的PVC环(内径10.5 cm)埋入土中36—38 cm。由于超过90%的植物根系分布在表土(0—20 cm)中[43],因此40 cm深的PVC环可以切断旧植物的根部,并防止新的根系在环内生长。该实验于2014年初进行,本文所用数据自2018年5月开始测量,从而留出足够的时间使PVC环内的植物根死亡[23]。因此,深PVC环中的CO2通量可以代表土壤异养呼吸。此方法与Wan等[44],Zhou等[9]和Wang等[45]使用的方法相同。对于浅PVC环,我们每隔两周检查、清除一次植物。用手将环内植物地上部分完全去除以阻断其对根系的碳供应。土壤自养呼吸由土壤呼吸减土壤异养呼吸得到。
在2018年生长季期间(5月—9月),使用Li-6400便携式光合仪(LI-COR Inc., Lincoln, NE, USA)的土壤呼吸室测定土壤总呼吸和异养呼吸,频率为每月两次,其中2018年7月上旬因持续降雨缺少一次测量。土壤呼吸速率和异养呼吸速率的测定选在阳光明媚的一天同时进行且在9:00—11:00完成。
1.3.2土壤温度和湿度的测定
在测量土壤呼吸速率的同时,测量土壤温度和湿度。使用LI-6400自带的温度探针测量10 cm处土层的土壤温度。使用手持式湿度探针(TDR100, Spectrum Technologies, USA)在靠近呼吸环的3个点处测量10 cm处土层的土壤湿度,测量结果为体积含水量。
1.4 数据分析
采用Microsoft Excel 2016进行数据初步整理,采用R[46]对数据进行统计分析,显著性水平设置为0.05。利用单因素方差分析方法比较不同处理样地之间土壤呼吸速率、自养呼吸速率和异养呼吸速率的差异。利用重复测量的方差分析比较不同增温和刈割处理下土壤呼吸及其组分的差异,以分析增温、刈割和测定时间及三者的交互作用对土壤呼吸速率及其组分的影响。利用一般线性模型(General linear model, GLM)分析土壤呼吸速率及其组分与土壤温度、湿度的关系,分别拟合一元线性回归模型、二元回归模型和指数回归模型,比较三种模型的AIC值,选择AIC值最小的模型;若两个模型的AIC差值不超过2,则选择较简单的模型[47]。使用OriginPro 2018C(OriginLab Corporation, Northampton, MA, USA)对分析结果作图。
2 结果
2.1 土壤温度和湿度对增温和刈割的响应
通过整个生长季的测量,增温和增温+刈割显著提高了0—10 cm土层的土壤温度(图1),增温小区和增温+刈割小区平均温度与对照相比分别提高了2.53 ℃和2.45 ℃(P<0.05)。与对照相比,增温处理下土壤湿度显著降低3.21%;增温+刈割处理下土壤湿度显著降低4.87%(图1)。

图1 不同处理下0—10 cm土层中土壤温度、土壤湿度的变化(平均值±标准误差)
2.2 增温和刈割对土壤呼吸速率及其组分的影响
在2018年整个生长季中,土壤呼吸速率(soil respiration rate,Rs)、土壤自养呼吸速率(soil autotrophic respiration rate,Ra)和土壤异养呼吸速率(soil heterotrophic respiration,Rh)季节动态的变化趋势基本一致(图2),并呈单峰曲线。从生长季开始到6月下旬,对照、刈割、增温、增温+刈割的Rs、Ra和Rh呈整体上升趋势,并在6月底达到峰值,土壤呼吸速率的最高值分别为8.18、10.9、10.73、13.38 μmol m-2s-1;土壤自养呼吸速率的最高值分别为1.34、2.87、2.50、4.15 μmol m-2s-1;土壤异养呼吸速率的最高值分别为6.83、8.04、7.08、7.41 μmol m-2s-1。6月下旬到8月下旬,土壤呼吸速率及其组分表现出下降趋势,8月下旬到生长季结束,各处理的土壤呼吸速率及其组分整体变化较平缓。
重复测量的方差分析表明增温、刈割和测定时间对土壤呼吸速率和异养呼吸速率均表现出显著影响(P<0.05),增温和刈割之间的交互作用对土壤呼吸和异养呼吸没有显著影响(P>0.05),但是对自养呼吸的影响是显著的(P<0.05)(表1)。
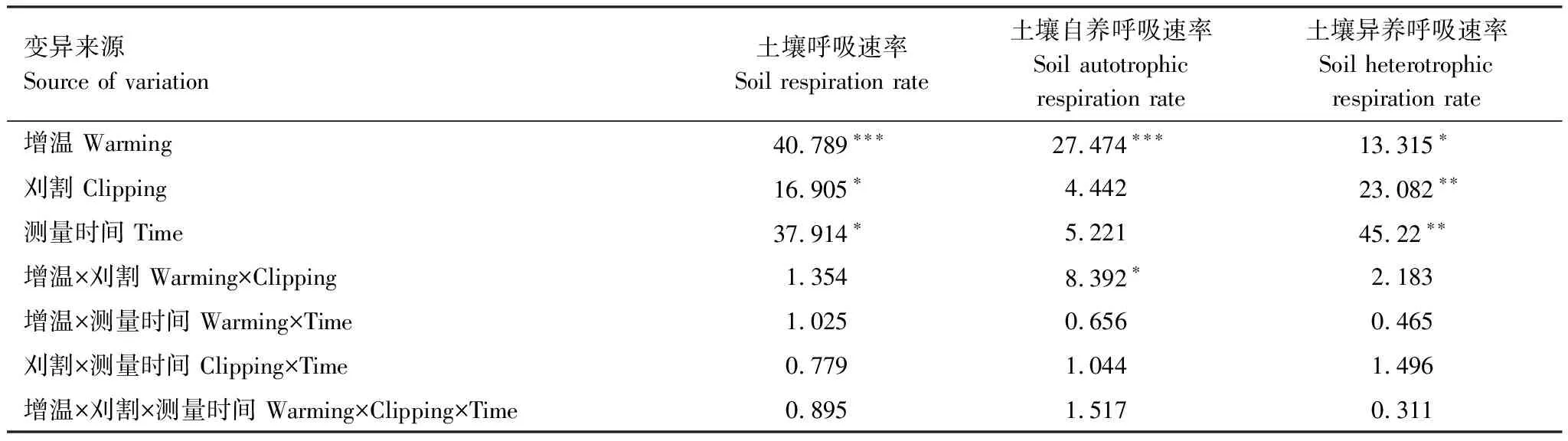
表1 不同处理下土壤呼吸速率、异养呼吸速率的重复测量方差分析结果(F-values)
在2018年生长季,与对照相比,单独增温使土壤呼吸速率和异养呼吸速率分别显著提高了31.65%、27.12%(P<0.01,P<0.01),单独增温对土壤自养呼吸速率没有显著影响;单独刈割使土壤呼吸速率提高了17.51%,但差异未达到显著性水平(P>0.05),单独刈割使异养呼吸速率显著提高了32.54%(P<0.01),单独刈割使自养呼吸速率降低了12.28%,但差异不显著(P>0.05)。与对照相比,同时增温和刈割促进了土壤呼吸、异养呼吸和自养呼吸(图2)。

图2 模拟增温和刈割对土壤呼吸速率、异养呼吸速率、自养呼吸速率的影响(平均值±标准误差)
2.3 土壤呼吸及其组分与土壤温度、土壤水分的关系
回归分析表明,土壤呼吸速率和异养呼吸速率与土壤温度存在显著的指数关系,土壤自养呼吸速率与土壤温度几乎没有显著的关系。增温和刈割对土壤呼吸速率、自养呼吸速率和异养呼吸速率的温度敏感性具有显著的负效应,在增温和刈割条件下,土壤呼吸速率及其组分的表观Q10值降低(表2)。此外,水分显著的影响了土壤呼吸速率及其组分(图3)。
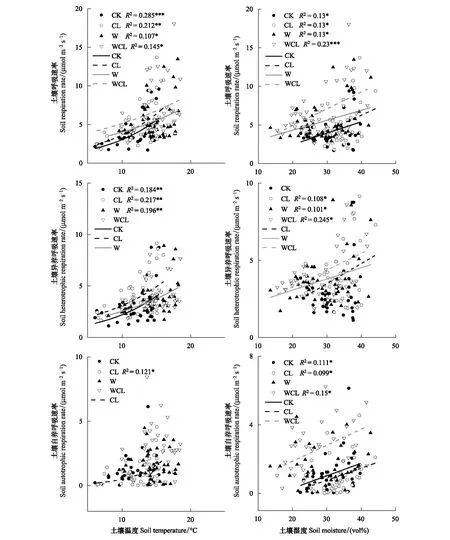
图3 土壤呼吸、异养呼吸、自养呼吸与土壤温度、土壤湿度之间的关系
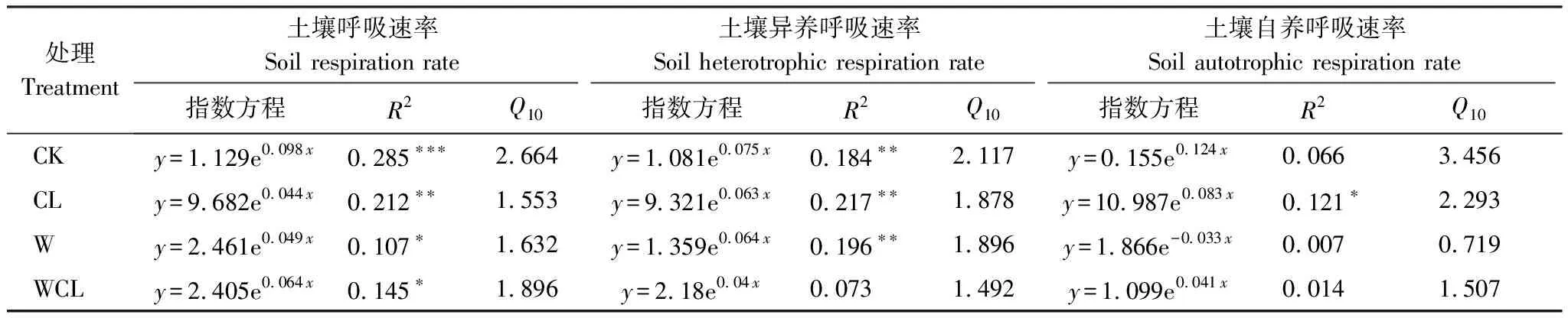
表2 不同处理下的土壤呼吸速率及其组分与土壤温度的关系
3 讨论
3.1 增温对土壤呼吸速率及其组分的影响
模拟增温试验的第5年,青藏高原高寒草甸土壤温度升高2.53 ℃,土壤呼吸速率显著提高31.65%(图2),高于全球尺度上的整合分析结果:温度升高2 ℃使得土壤呼吸平均提高12%[27],说明高寒草甸土壤呼吸的温度敏感性比全球平均水平要高。同时,我们的研究发现增温2.53 ℃使异养呼吸提高27.12%(图2),与最近的一项全球整合分析中升温2.0 ℃的情况下,异养呼吸平均增加21%的结果基本一致[27]。而我们的研究中增温对土壤自养呼吸没有显著影响的结果也与全球整合分析的结果一致[27]。在全球气候变化的背景下,有大量的野外控制实验探讨增温对陆地生态系统碳循环的影响,但是,有关土壤呼吸及其组分对增温响应的研究结果仍存在争议[19,48]。例如,对西藏半干旱高山草原[50]、内蒙古半干旱草原[29]等生态系统的研究结果显示升温诱导的水分胁迫产生的负效应抵消了高温对土壤呼吸和异养呼吸的正效应,进而导致增温抑制土壤呼吸和异养呼吸;与之相反,对针叶林[49],亚北极荒地[50]、永冻土区高寒草甸[51]等生态系统的研究结果表明温度升高能够提高土壤微生物和酶的活性,刺激土壤异养呼吸,从而促进土壤CO2的排放。我们的研究中连续4年增温后土壤呼吸速率显著提高可能是由于温度升高使得土壤酶活性提高,而土壤酶活性的提高与增温条件下土壤微生物数量的增加和微生物活性的提高有关。土壤微生物数量增加和土壤微生物活性的增强刺激了微生物的代谢活性,加速土壤碳矿化,促进了土壤异养呼吸,进而促进土壤碳排放[52-53]。我们并未发现增温对自养呼吸的显著影响,可能与实验中增温并未改变细根生产和死亡的年平均值有关[54]。
3.2 刈割对土壤呼吸速率及其组分的影响
刈割可以通过改变土壤微气候以及根系和土壤微生物的C底物可利用性来抑制土壤呼吸[21, 37]。然而我们的研究中刈割对土壤呼吸影响不显著,这可能是由于异养呼吸和自养呼吸对刈割的相反的响应趋势造成的。土壤微生物呼吸很大程度上受到土壤温度、土壤水分以及微生物生物量和微生物呼吸的影响[55]。刈割引起的土壤温度的增加能够提高土壤微生物活性和其对土壤有机质的矿化,导致微生物呼吸速率的提高[56]。本研究结果中刈割在一定程度上提高了土壤温度和土壤水分,微生物生物量也有所提高,最终表现为刈割刺激异养呼吸。自养呼吸主要依靠光合同化产物的供给[57]。刈割能够改变植物的分配策略,将储存在根系中的C运输到地上部分用于枝叶的再生,植物的地下C分配将会减少[58-59]。本研究中刈割导致地下生物量减少,进而抑制自养呼吸。异养呼吸的增加被自养呼吸的减少部分地抵消掉,导致土壤呼吸没有受到刈割的显著影响。
3.3 土壤温度和土壤水分对土壤呼吸及其组分的影响
Q10被定义为温度每增加10 ℃,土壤呼吸增加的倍数,是用于评估土壤呼吸及其组分温度敏感性的重要参数。增温和刈割降低了土壤呼吸及其组分的温度敏感性(表2),这与之前的许多研究发现土壤呼吸的温度敏感性随着土壤温度的升高[9, 60-61]或刈割[21,62]而降低的结果相一致。增温导致自养呼吸和异养呼吸温度敏感性的降低可能是植物呼吸或微生物活动对变暖的直接生理适应[63-64],植物和微生物的生理适应最终导致土壤呼吸温度敏感性的降低。另外,增温可能导致土壤微生物群落的变化,并因此导致温度敏感性的变化[48]。刈割降低土壤呼吸及其组分的温度敏感性主要有以下3个原因。首先,刈割导致光合同化C产物向根系的分配减少[9]。其次,刈割引起的土壤温度的升高使土壤呼吸及其组分产生温度适应性[63]。最后,刈割引起微生物群落结构的改变,微生物更加适应环境,导致温度敏感性降低[65]。
土壤呼吸及其组分对增温的响应受土壤水分可用性的调节[19]。土壤含水量过低会降低细胞外酶的扩散速度以及微生物的移动,降低呼吸底物与微生物的接触机会[66],进而影响土壤呼吸及其组分。土壤含水量的降低会影响土壤通气性,进而影响微生物和根的呼吸。目前对于水热因子对土壤呼吸的影响还没有一致的结论,主要的观点有3种:(1)土壤温度和水分作为主要的环境因子,共同影响着土壤呼吸速率[67-68];(2)土壤水分对土壤呼吸的影响要强于土壤温度[69];(3)土壤温度对土壤呼吸的影响强于土壤水分[70]。土壤温度和水分对土壤呼吸及其组分影响的强弱主要是由于研究地点的限制因子不同导致的[71]。本研究发现土壤温度和水分可以共同解释土壤呼吸、自养呼吸、异养呼吸变化的38%、14%、35%,高于土壤温度或者水分单一因子对土壤呼吸及其组分变化的解释,表明在该高寒草甸,土壤温度和水分共同驱动土壤呼吸及其组分的变化。这是由于高寒草甸具有水分充足温度低的特点,温度为限制因子,温度升高能够刺激土壤呼吸;另一方面,尽管该地区水分充足,温度升高引起的土壤水分的降低对生态系统碳通量的影响显著[72],导致该地区土壤温度和水分共同调节土壤呼吸及其组分。
4 结论
基于5年的模拟增温和刈割野外控制实验,我们发现增温显著提高青藏高原高寒草甸土壤呼吸速率和异养呼吸速率,而增温对自养呼吸没有显著影响;刈割对土壤呼吸的刺激作用不显著,但是促进了异养呼吸,在一定程度上抑制了自养呼吸;增温和刈割之间的交互作用不显著。土壤呼吸速率及其组分与土壤温度和湿度之间存在显著的正相关。因此,在该高寒草甸生态系统,土壤水分、温度共同调节土壤呼吸和异养呼吸。本研究表明青藏高原东缘高寒草甸土壤碳排放与全球变化如增温和刈割之间存在正反馈。
