麋鹿鹿角脱落、群主更替、产仔的年节律及其环境影响因子
2020-11-12程志斌刘定震白加德钟震宇林润生田东晓王丽斌
程志斌, 刘定震, 白加德,*, 钟震宇, 林润生, 田东晓, 王丽斌, 刘 田
1 北京麋鹿生态实验中心, 北京 100076 2 北京师范大学生命科学学院, 教育部生物多样性与生态工程重点实验室, 北京 100875 3 北京市气象信息中心, 北京 100089
地球上的光、温度、湿度等生态因子具有明显的周期性,因而在生物的进化和发展中形成了昼夜节律、月节律、年节律等生物节律[1-3]。动物的年节律如繁殖、脱角、换羽(毛)、迁徙、冬眠等行为是一种复杂的生物学现象,它是动物对光照、温度、湿度、水等非生物条件,食物、种内关系和种间关系等生物条件的适应,每个物种均有各自的生物节律特点,来适应外界复杂而变动的外界环境条件[4-8]。因此,通过动物的生物节律来研究动物与环境之间的适应性关系始终是生态学家和物候学家的研究热点[9-12]。繁殖活动是哺乳动物生命中最为重要的组成部分,已有大量关于鸟类[13-15]和兽类[16-18]繁殖年节律会着温度、降水等环境变化而产生适应性变化的报道,有蹄类作为哺乳动物中的重要类群,其繁殖的年节律受光周期、气候、营养、纬度、种群密度等环境因子的影响[19-20]。然而结合具体的平均气温和降水量来探讨气候对哺乳动物繁殖特征年节律影响方面的报道较少。
各物种采取的繁殖策略是多种多样的,包括从高度季节性到完全无季节性的繁殖模式[21]。生活在高纬度地区的鹿科动物属于前者,雄鹿每年的繁殖周期包含发情期、身体恢复期、鹿角脱落期和长茸骨化期四个阶段,雌鹿每年的繁殖周期包含发情期、怀孕期、产仔期和哺乳期四个阶段[22]。首先,鹿角是鹿亚科(Cervinae)和空齿鹿亚科(Odocoileinae)动物特有的性二型特征[23],是繁殖期雄性争夺交配权和雌性选择雄性的重要指标[24-25],它是哺乳动物唯一可再生的器官[23],高纬度地区鹿科动物的鹿角每年周期性脱换[23,26]。其次,占群繁殖,是一雄多雌的哺乳动物雄性繁殖期的重要行为特征之一,如川金丝猴(Rhinopithecusroxellana)[27]、象海豹(MiroungaLeonina)[28]、藏羚羊(Pantholopshodgsonii)[29]、普氏野马(Equusprzewalskii)[30]等。第三,雌性鹿类动物的每次的发情持续时间较短,通常只有几个小时[31],不易于观察,因此更为直观的产仔周期成为研究雌鹿的繁殖年节律特征的首选[17,32]。
尽管关于鹿类动物年节律特征的研究有许多,但主要为单一或者两个繁殖特征,如马鹿[17, 33-34](Cervuselaphus)、狍[35](Capreoluspygargus)、驯鹿[36](Rangifertarandus)等广布物种的角生长、发情或者产仔的年节律,以及白尾鹿(Odocoileusvirginianus)血液中激素的年节律变化[37-38]。而同一群体的多个繁殖特征的变化差异、相互之间的关系及其环境影响因子等方面的研究甚少。麋鹿(Elaphurusdavidianus)为我国特有物种,国家一级保护动物,IUCN红皮书列为野外灭绝,属于典型的季节性繁殖动物,雄性具角,具有自身独特的脱角周期[23]。麋鹿的婚配制度为典型的一雄多雌的后宫制[39-40],雄性麋鹿采用序列占群的繁殖策略[40],通过顶角争斗获得群主地位和交配权[41-43],目前尚未见到麋鹿年节律研究的相关报道。本文通过研究麋鹿鹿角脱落、群主更替、产仔等繁殖特征的年节律及其环境影响因子,为深入了解哺乳动物在进化过程中对环境的适应性,以及为保护麋鹿这一珍稀物种提供科学依据。
1 材料与方法
1.1 研究地概况
研究地为北京南海子麋鹿苑(39°50′N, 116°30′E),位于北京城南10 km,占地60 hm2,麋鹿半散放活动的保护区面积约40 hm2,为古代元明清皇家苑囿南海子的一部分,是麋鹿模式种产地、我国最后灭绝地和我国第一处重引入地。苑内优势植物有牛筋草(Eleusineindica),大画眉草(Eragrostiscilianensis),马唐(Digitariasanguinalis)和狗尾草(Setariaviridis)等;年均气温为13.1℃,1月平均气温为-3.4℃,7月平均气温为26.4℃;年降雨量约为600 mm[44]。苑内无河流经过,为人工湖,水域靠降雨和抽取地下水补给。
北京南海子麋鹿种群是麋鹿在国内灭绝后建立的第一个半散放迁地种群。1985年和1987年北京南海子麋鹿苑从英国乌邦寺共引入38只麋鹿。回归35年来,北京南海子麋鹿苑分别向湖北石首、江西鄱阳湖、浙江慈溪、河北滦河上游等地输送536只麋鹿,建立了39个迁地保护种群,在长江中下游建立了麋鹿野生种群。北京南海子麋鹿种群维持在130—210只之间(雄雌性比约1∶1),采取半散放的饲养方式,由于夏秋两季地面草本植物生物量有限,一年四季均有人工补饲。
1.2 数据采集及处理
1.2.1鹿角脱落数据采集及处理
2014—2016年,每一年度掉角期12月—翌年2月,每天上午和下午各一次在麋鹿生活区域巡视,收集麋鹿自然脱落的角,且在角上标记脱落日期。选择鹿角脱落开始时间、鹿角脱落高峰期、鹿角脱落结束时间作为鹿角脱落年节律因子。
脱角数量达到半数为脱角中位值日期[45],由于脱角中位值日期处于脱角集中时间段,为了便于讨论,将其定义为鹿角脱落高峰期;由于2岁龄鹿角(称笔杆角,无分叉)在脱落后掉于草丛中难以收集,因此本文将2叉(3岁龄)及2叉以上的鹿角全部脱落完毕的日期定义为鹿角脱落结束时间。
鹿角回收率(%)=(鹿角回收数/2岁以上雄性数×2)×100%
1.2.2群主更替数据采集及处理
2015—2017年,每年发情期5—9月,一周观察4—7 d,每天观察0.5—2.5 h,使用双筒望远镜(SWAROVSKI 8×42WB)、相机(Canon550)记录群主的特征,通过鹿角形状、耳标、耳缺、项圈、体型等区分个体和年龄。群主为占有和控制发情雌鹿群(后宫)的等级序位高的雄鹿[41]。群主有两种方式控制发情雌鹿群:一是直接控制发情雌鹿群,二是控制发情场[40]。由于北京南海子麋鹿为半散放种群,发情场地有限,在水边泥滩的高大乔木下,占有发情场地是群主最喜欢的方式。本研究中群主为占有固定发情场的群主,而占有临时雌性群(指离开固定发情场外出觅食形成的临时雌性群,它们通常1 h内会被群主驱赶回固定发情场或者被群主控制)的临时群主不计算在内。
将第一只群主开始占有和控制雌鹿群,且占有固定发情场的日期定义为第一次发情期开始时间(即群体而言雄性发情开始时间)[17];第一次发情期间群主更替次数达到半数的日期为群主更替中位值日期[46],由于群主更替中位值日期处于群主更替集中时间段,为了便于讨论,将其定义为群主更替高峰期;对于整个种群而言,将当年第一次发情期的群主占群更替结束的日期定义为第一次发情期结束时间(通常群主全身为夏毛,未开始更换冬毛);将当年第二次发情期的群主开始占群的日期定义为第二次发情期开始时间(通常群主夏毛已经脱落完毕,正在长冬毛);将当年第二次发情期的群主占群更替结束的日期定义为第二次发情期结束时间(即群体而言雄性发情结束时间)。
选择第一次发情期开始时间、群主更替高峰期、第一次发情期结束时间、第二次发情期开始时间、第二次发情期结束时间作为群主更替年节律因子。
1.2.3产仔数据采集及处理
2016—2018年,每一年产仔期3—7月,每天使用望远镜观察麋鹿群,尤其是离群即将分娩或者正在分娩的母鹿,上午和下午各观察鹿群及其活动场所一次,做好新生仔鹿出生记录。选择产仔期开始时间、产仔高峰期、产仔期结束时间作为产仔的年节律因子。
产仔数量达到半数的日期为产仔中位值日期[47],由于产仔中位值日期处于产仔集中时间段,为了便于讨论,将其定义为产仔高峰期。
1.2.4环境数据采集及处理
2014年1月—2018年7月气象数据由北京市气象信息中心提供,站点为大兴站(站号54594,国家一般气象站),距离研究地点10.3 km;2018年7—12月的气象数据取自中国地面国际交换站气候资料日值数据集,站点为北京站(站号54511,国家基本气象站),距离研究地点3.7 km。选择气温(3—9月、12月的月平均气温,春、夏、秋、冬季4个季节的季平均气温,年平均气温)、降水(春季第一场雨、春季第一场透雨、雨季开始时间、雨季结束时间、年降雨量)作为气候因子。
春季第一场透雨参照姚佩珍和张强[48]北京春季第一场透雨指标的方法,由于2014年、2015年、2017年、2018年春季降雨量均超过70 mm,降水充沛,第一场透雨日的标准按照降雨量超过或者约等于当旬轻旱的平均标准值统计。雨季开始和结束时间参照于晓澄等[49]华北雨季开始早晚的方法。
由于麋鹿半散放区域的面积不变,以年末种群数量作为种群密度因子。
1.3 作图
按照实际日期制作每一年度麋鹿鹿角脱落、群主更替、产仔和日降水量情况变化图。日降水量存在的缺失值在图中不显示。
2 结果
2.1 麋鹿鹿角脱落、群主更替、产仔的年节律
2.1.1鹿角脱落的年节律
由表1可知,2014—2016年麋鹿角的回收率为81.37%—90.70%,说明可根据实际所收集到的麋鹿角脱落日期统计鹿角脱落高峰期。图1表明,整个种群而言,麋鹿角脱落的年节律存在差异,2014—2016年的鹿角脱落开始时间分别为12月16日、12月7日、12月3日,鹿角脱落高峰期分别为12月31日、12月26日、12月21日,鹿角脱落结束时间分别为2月7日、2月4日、1月17日。说明北京南海子麋鹿种群鹿角脱落时间开始于12月,结束于1月下旬或2月上旬;2015年、2016年的鹿角脱落开始时间、鹿角脱落高峰期、鹿角脱落结束时间与前一年比较均出现同步提前的现象。
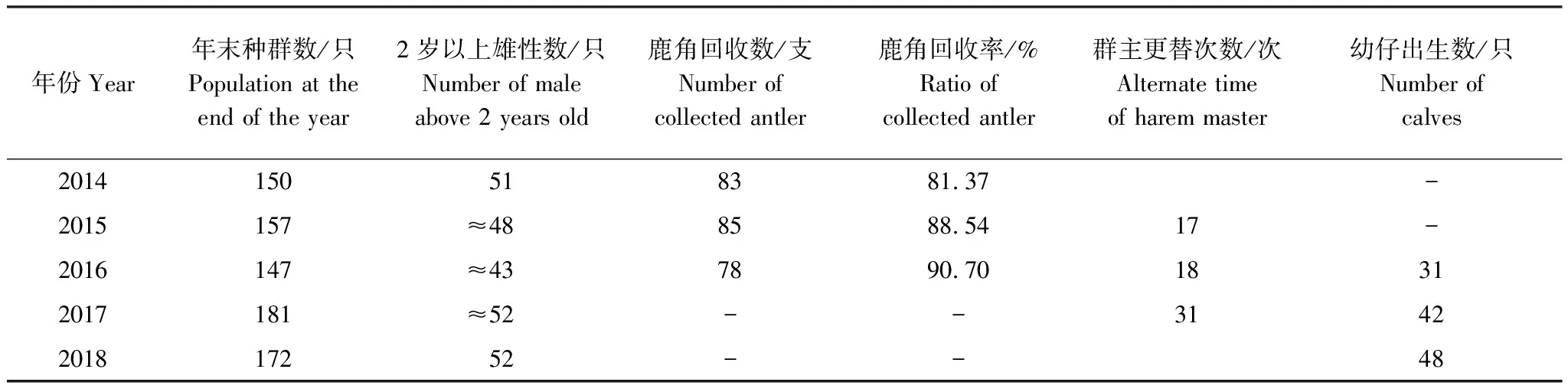
表1 北京南海子麋鹿苑2014—2018年麋鹿年末存栏、脱角、群主更替次数、产仔情况
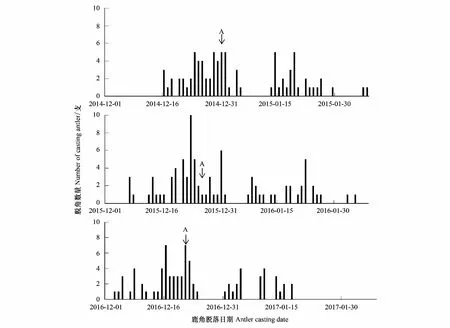
图1 北京南海子麋鹿苑2014—2016年度麋鹿角脱落情况
2.1.2群主更替的年节律
图2表明,雄性麋鹿存在第二次发情期的现象;2017年出现两个固定发情场所,其中第二个固定发情场6月7日出现,6月28日消失。整个种群而言,群主更替存在年节律的差异,2015—2017年的第一次发情期开始时间分别为6月9日、5月30日、5月17日,群主更替高峰期分别为7月22日、7月4日、6月28日,第一次发情期结束时间分别为9月1日、8月12日、8月15日,第二次发情期开始的时间分别为9月1日、8月15日、8月17日,第二次发情期结束时间分别为9月8日、9月9日、9月4日;2015—2017年发情期时间分别为91d、102d、110d。说明北京南海子麋鹿苑麋鹿发情期开始于5月下旬或6月上旬,结束于9月上旬;2016年和2017年的第一次发情期开始时间、群主更替高峰期与前一年比较均出现同步提前的现象;而第一次发情期结束时间、第二次发情期开始时间和第二次发情期结束时间不存在同步提前的现象;从2015年至2017年,整个鹿群发情的时间有延长的趋势。
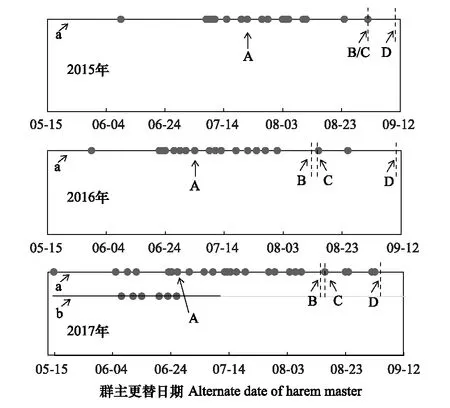
图2 北京南海子麋鹿苑2015—2017年麋鹿群主更替情况
2.1.3产仔的年节律
由表1和图3可知,整个种群而言,麋鹿产仔年节律存在差异,产仔期开始时间2016—2018年分别为4月12日、3月28日、3月16日;产仔高峰期分别为4月23日、4月17日、4月3日,产仔期结束时间分别为7月25日、7月15日、5月25日;产仔期分别长达104 d、109 d、70 d。2016年70.97%的幼仔集中在高峰期前后的13 d内出生,2017年64.29%的幼仔集中在高峰期前后的21 d内出生,2018年56.25%的幼仔集中在高峰期前后的19 d内出生。说明北京南海子麋鹿苑麋鹿产仔期开始于3月中旬至4月中旬,结束于5月下旬至7月下旬;2017年和2018年产仔期开始时间、产仔高峰期、产仔期结束时间与前一年比较均出现同步提前的现象;麋鹿产仔同期化变弱。
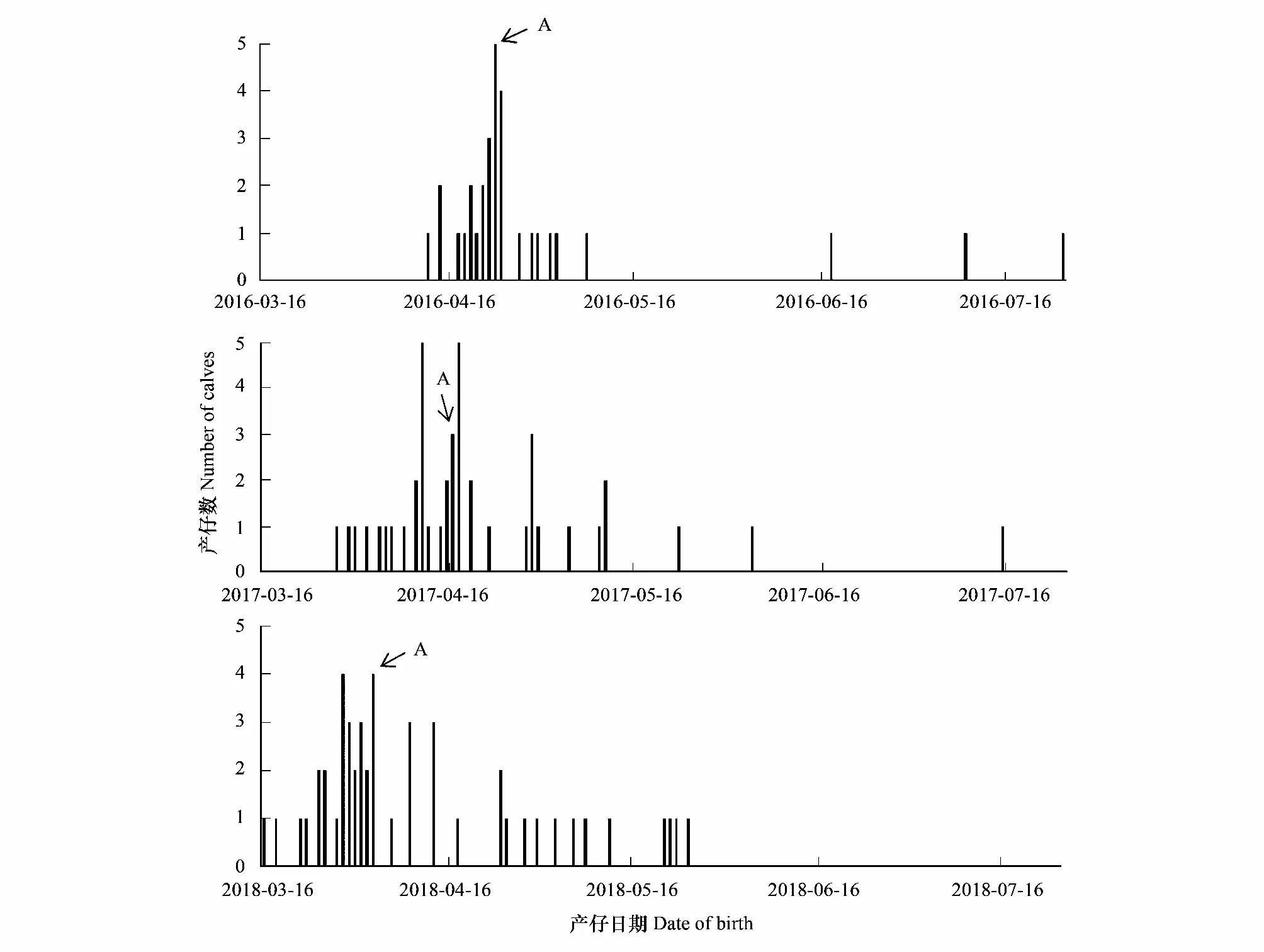
图3 北京南海子麋鹿苑2016—2018年度麋鹿产仔情况
2.2 环境因子
由表1可知,北京南海子麋鹿苑2014—2018年麋鹿年末种群数量分别为150只、157只、147只、181只、172只。
2014—2018年的月平均气温见图4;年平均气温分别13.88℃、13.62℃、13.36℃、13.65℃、13.34℃;春季平均气温分别为16.42℃、15.54℃、15.40℃、16.27℃、15.04℃,夏季平均气温分别为26.30℃、26.17℃、26.37℃、26.08℃、27.51℃,秋季平均气温分别为13.56℃、12.77℃、12.89℃、12.57℃、13.36℃。2014—2017年冬季(12月、翌年1—2月)平均气温分别为-0.13℃、-1.36℃、-0.11℃、-2.00 ℃。
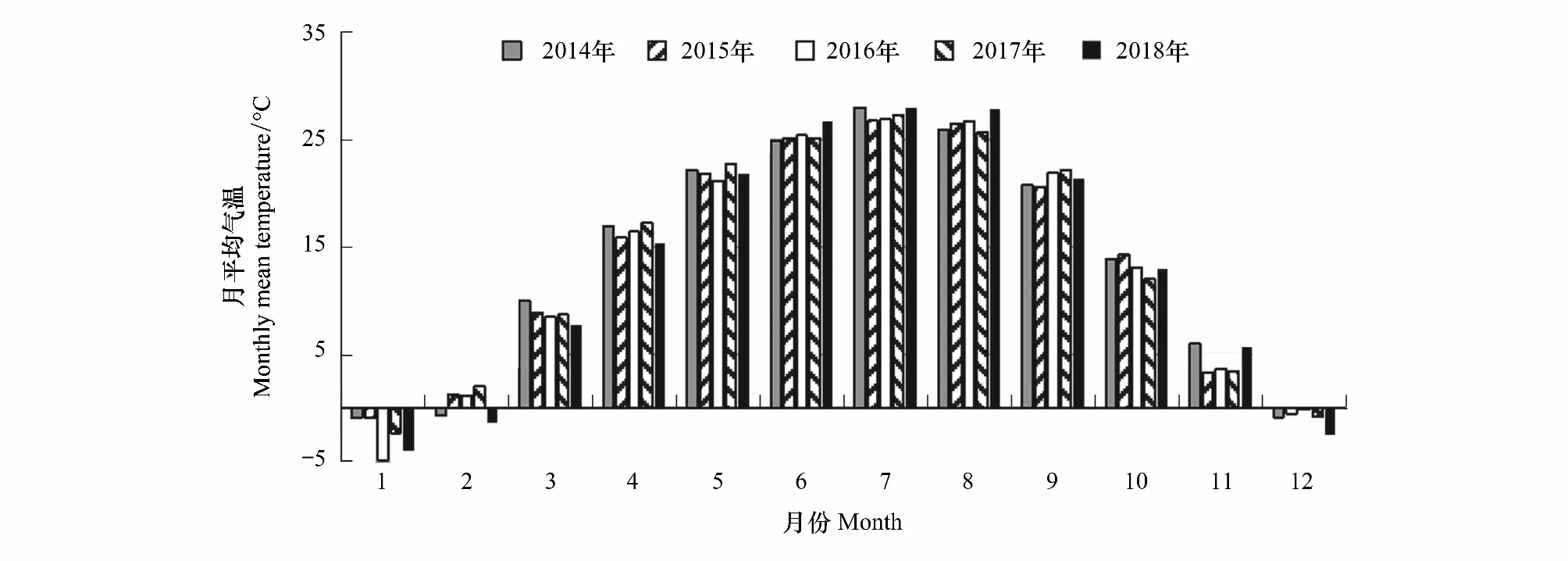
图4 北京南海子麋鹿苑2014—2018年月平均气温
2016年春季 3—5月共7场降雨,降雨量总和为30.7 mm,属于干旱,7场降雨均没有达到春季第一场透雨的标准。2014—2018年的年降雨量分别416.8 mm、505.8 mm、722.7 mm、682.9 mm、537.1 mm。由图5可知,2014—2018年的第一场春雨分别为3月28日、3月31日、4月2日、3月20日、3月17日;2014年、2015年、2017年、2018年的春季第一场透雨分别为4月17日、3月31日、3月23日、4月5日;2014—2018年的雨季开始时间分别为7月2日、7月17日、7月19日、7月4日、7月8日;2014—2018年的雨季结束时间分别8月13日、8月31日、7月28日、8月27日、8月13日。

图5 北京南海子麋鹿苑2014—2018年日降水量
2.3 鹿角脱落、群主更替、产仔的年节律之间的同步关系
由图1—3可知,当2014—2016年麋鹿鹿角脱落开始时间、鹿角脱落高峰期和鹿角脱落结束时间逐年提前时,2015—2017年第一次发情期开始时间、群主更替高峰期也随之逐年提前;2016—2018年的产仔期开始时间、产仔高峰期和产仔期结束时间也紧接着逐年提前。2015—2017年的第一次发情期开始时间至翌年产仔期开始时间间隔分别为308 d、302 d、303 d,平均间隔为304.3 d;2015—2017年的群主更替高峰期至翌年产仔高峰期间隔分别为276 d、287 d、279 d,平均间隔为280.7 d。说明2014—2016年的麋鹿鹿角脱落年节律因子(鹿角脱落开始时间、鹿角脱落高峰期和鹿角脱落结束期)、2015—2017年群主更替年节律因子(第一次发情期开始时间和群主更替高峰期)、2016—2018年的产仔年节律因子(产仔期开始时间、产仔高峰期和产仔期结束时间)存在明显的同步关系。
2.4 鹿角脱落、群主更替、产仔的年节律与环境因子之间的关系
图1—5可知,麋鹿鹿角脱落的年节律与秋季平均气温、冬季平均气温、年平均气温不存在随着平均气温升高或降低而提前或者延迟的现象;2015—2017年群主更替的年节律与2014—2016年的年平均气温和秋季平均气温,与本年度夏季平均气温不存在随着平均气温升高或降低而提前或者延迟的现象;2016—2018年产仔的年节律与2015—2017年秋季平均气温、冬季平均气温和年平均气温,与本年度春季平均气温不存随着平均气温升高或降低而提前或者延迟的现象。产仔期开始时间与3月平均气温,产仔期开始时间和产仔高峰期与4月平均气温,第一次发情期开始时间与5月平均气温,第一次发情期开始时间和群主更替高峰期与6月平均气温,群主更替高峰期与7月平均气温,第一次发情期结束时间和第二次发情期开始时间与8月平均气温,第二次发情期结束时间与9月平均气温均不存在随着平均气温升高或降低而提前或者延迟的现象。但鹿角脱落开始时间、鹿角脱落高峰期和鹿角脱落结束时间存在随着12月平均气温升高而提前的现象;产仔期开始时间和产仔高峰期存在随着前一年9月平均气温的升高而提前的现象。说明除群主更替年节律外,产仔年节律和鹿角脱落年节律表现出受气温因子影响的现象。
鹿角脱落开始时间、鹿角脱落高峰期和鹿角脱落结束时间均与年降雨量存在随着年降雨量的增加而提前的现象。2015—2017年第一次发情期开始时间、群主更替高峰期的年节律存在随着2014—2016年年降雨量的增加而提前的现象。2016—2018年产仔期开始时间和产仔高峰期不存在随着2015—2017年年降雨量的增加或减少而提前或延迟的现象。产仔期开始时间、产仔高峰期和第一次发情期开始时间均不存在随着春季第一场雨、春季第一场透雨的提前或者延迟而提前或者延迟的现象。群主更替高峰期不存在随着雨季开始的提前或者延迟而提前或者延迟的现象;第一次发情期结束时间、第二次发情期开始时间和第二次发情期结束时间均不存在随着雨季开始时间、雨季结束时间的提前或者延迟而提前或者延迟的现象。2015年的群主更替高峰期出现在雨季期间,2016年和2017年的群主更替高峰期出现在雨季开始之前。说明,产仔年节律没有表现出与降水因子的变化而出现相应的同步变化,鹿角脱落年节律表现出随着年降雨量的增多而提前的现象;第一次发情期开始时间、群主更替高峰期的年节律表现出随着前一年度年降雨量的增多而提前的现象。
由图1—3和表1可知,麋鹿鹿角脱落年的节律、群主更替的年节律、产仔的年节律均不存在随着种群密度升高或降低而提前或者延迟的现象。
3 讨论
3.1 麋鹿鹿角脱落、群主更替、产仔的年节律
高纬度地区的有蹄类,均具有各自的繁殖节律特征来适应高度季节性的生境。本研究中,北京南海子麋鹿种群鹿角脱落时间从12月开始,1月下旬或2月上旬结束,这与极地物种驯鹿、欧亚大陆高纬度地区的广布物种狍的鹿角脱落时间相一致,而马鹿、梅花鹿(Cervusnippon)、黇鹿(Damadama)等大部分鹿科动物为春夏之际开始脱角[50]。有蹄类喜欢选择在春夏之际气候和食物条件最适的时候集中产仔,研究显示麋鹿产仔期为3—7月,集中于4—5月,这与温带地区和寒带地区的其他鹿类相一致[50]。本研究表明,麋鹿发情期为5月下旬或6月上旬开始,9月上旬结束,麋鹿存在二次发情期现象;这与李春旺等[42]研究结果相同。本研究发现群主更替高峰期与翌年产仔高峰期间隔平均为280.7 d,而人工圈养的麋鹿妊娠期平均为280 d[51],因此可以根据前一年的群主更替高峰期推测第二年的产仔高峰期。有研究表明,黑尾鹿(Odocoileushemionus)发情高峰期可以预测翌年产仔时间的情况[52];可通过马鹿鹿角脱落年节律来预测紧接着的雄性发情年节律[17]。
高纬度地区鹿类动物的鹿角脱落、发情、产仔等繁殖周期具有同步性[53]。雄鹿为了提高等级序位,增加自身繁殖后代的贡献率,可通过使角提前骨化而先进入发情占群期[46]。南非一些反刍动物在反捕食策略上表现为产仔行为和交配行为同步[54]。本研究发现麋鹿鹿角脱落、群主更替、产仔的年节律存在明显的同步关系,其中鹿角脱落开始时间、鹿角脱落高峰期、鹿角脱落结束时间、第一次发情期开始时间、群主更替高峰期、产仔期开始时间、产仔高峰期、产仔期结束时间与前一年度比较均出现同步提前的现象。这与马鹿繁殖特征的年节律相似[33,55],Clements等[46]对野生马鹿长达28年的研究表明,马鹿群体鹿角脱落开始时间、鹿角脱落结束时间、雄性发情开始时间、雄性发情高峰期、雄性发情结束时间、雌性发情开始时间和产仔期开始时间等七个繁殖特征之间均呈显著正相关关系,并呈线性逐年同步提前现象[17]。
3.2 环境影响因子
3.2.1气候因子
光周期具有恒定性和规律性,随着地理纬度和季节而很有规律地变动,它是引起动物繁殖活动年节律变化的直接原因[3, 19, 56],通过神经——垂体——性腺轴控制动物繁殖期[42, 57]。根据动物繁殖与光照长短的关系,可将动物分为长日照动物和短日照动物,羚羊类和鹿类只有在白昼逐步缩短的秋冬之际才开始性腺活动,它们属于短日照动物[3],然而麋鹿是鹿类中唯一的夏季发情的短日照发情动物[42, 50, 53],这与本研究的结果相一致,麋鹿发情期为5月下旬—9月上旬,发情高峰期集中在光照时长最长的6—7月。通过适应光周期的变化,温带地区的鹿类动物产生繁殖年节律,它们可以优化调整产仔时间,使其与最适的环境气候因子(如食物)相同步[58]。本研究表明,麋鹿的鹿角脱落、群主更替、产仔等繁殖特征存在明显的年节律,恰好说明它们受光周期的影响,形成周期性变化的年节律。有研究表明,将南半球的普度鹿(Pudupudu)迁移至北半球,它的繁殖期会很快与当地的光周期同步[58]。
光在决定动物繁殖年节律中起着重要作用,但并不是说它是唯一的信号,温度、湿度、降水等气候原因对动物繁殖年节律也有重要的影响[3]。非洲的有蹄类如斑马(Equusquagga)的产仔年节律同环境的气温、降水量的变动有关[59]。地中海马鹿雌性发情开始时间在更加干旱和炎热的春季会推迟;在更加凉爽和降雨量多的秋季会提前[60]。我们的研究中,麋鹿的鹿角脱落、群主更替、产仔等繁殖特征连续3个年度的年节律不是固定不变的,而是存在年际差异,说明它们受到其他因子的影响。
温度可以影响动物的生活史特点,如使狍的产仔年节律提前[35]。气候变暖伴随着有效积温的逐年升高,使得苏格兰Rum岛植物的生长量增加,给当地马鹿带来更多的食物,导致鹿角脱落开始时间、鹿角脱落结束时间、产仔期开始时间、雄性发情开始时间、雄性发情结束时间呈逐年提前的趋势[17, 46]。而且马鹿鹿角开始脱落的时间与前一年秋季温度有关,因为秋冬季发情结束之后的食物多少与秋季温度密切相关[61],而且前一年9月和当年4月平均气温升高会使马鹿鹿角脱落年节律提前[62]。我们的研究结果也显示产仔期开始时间和产仔高峰期存在随着前一年9月平均气温的升高而提前的现象,鹿角脱落年节律存在随着每年12月平均气温升高而提前的现象。
降水是陆生动物的重要环境因子,降水会改变哺乳动物繁殖的年节律,本研究表明,麋鹿第一次发情期开始时间、群主更替高峰期的年节律表现出随着前一年度年降雨量的增多而提前的现象。雌性驯鹿在4月降水量大的年份,表现出产仔期开始时间推迟[18];但是本研究中,产仔年节律未表现出随着降水因子的变化而发生相应的同步变化。有研究认为,当冬季降雪量增多时,第二年马鹿鹿角脱落开始时间会推迟[62-63],本研究中麋鹿鹿角脱落年节律表现出随着年降雨量的增多而提前的现象。
3.2.2营养和种群密度因子
营养(或食物)也是动物繁殖年节律变化的重要原因[59]。植物提前生长或者增加生物量,可为食草动物在关键的繁殖期提供更多的食物[64-65]。增加或者提前的春季食物可使雌鹿妊娠期和雄鹿鹿角脱落时间提前;植物生长时间延长,可提高哺乳期和长茸期食物的质量,让它们提前进入秋季的发情期[17]。动物能够根据产仔的营养需要,调整产仔时间,将产仔时间与环境气候最适时间(如食物供应高峰期)同步[35, 52, 63],提高幼崽存活率[32]。本研究中,2015—2017年麋鹿鹿群发情时间有延长的趋势,2016—2018年麋鹿产仔同期化变弱,分析原因,可能与北京南海子麋鹿种群为半散放状态,食物营养充分,食物压力较小有关。圈养动物与野生动物的产仔年节律不同,受气候和食物等因子的影响,圈养的马麝(Moschuschrysogaster)产仔同期化比野生种群更弱[66]。白尾鹿的动物园种群产仔期开始时间为2月,而野生种群4月才开始产仔[25]。南美草原鹿(Ozotocerosbezoarticus),阿根廷干旱草原生境中的种群全年产仔,产仔80%集中在10月至翌年4月出生;而同纬度相邻的乌拉圭半干旱大陆性气候生境中的种群为季节性产仔,产仔同期化更加集中,高峰期推迟至夏季末或秋季初食物充沛的时期;而热带环境下的巴西种群,由于明显的旱季和雨季,食物变化更为显著,产仔同期化最显著,产仔高峰期为8—9月,与当地的雨季完全同步[32]。但是,有学者认为,这一同步现象不是立即发生的[67-68],有时会存在时间误差,对于狍而言它的胚胎滞育策略就显得尤为重要[35]。
野外条件下食物和生存空间有限,种群密度大,意味着个体之间的竞争力大。地中海马鹿雌性发情开始时间在种群密度大的年份会推迟;在种群密度小的年份会提前[60]。苏格兰Rum岛马鹿种群也存在高密度种群会推迟角脱落开始时间和角脱落结束时间的现象[62]。本研究表明,麋鹿鹿角脱落、群主更替、产仔的年节律均不存在随着种群密度升高或降低而提前或延迟的现象,也许和半散放状态下,每天人工补饲,不存在食物压力有关。
3.2.3纬度
随着纬度的变化鹿类动物繁殖年节律会随之变化[52]。江苏大丰麋鹿脱角开始时间和结束时间早于北京南海子种群[23]。本研究中,北京南海子麋鹿群主更替高峰期为6—7月,而低纬度地区的江苏大丰麋鹿种群雄性发情高峰为提前为5—6月[42]。这与其他鹿类动物相似,墨西哥的白尾鹿发情开始时间为7月,而美国和加拿大地区为10月[25],高纬度和低纬度地区马鹿发情和产仔年节律研究的结果也表现出这一特征[33]。然而,本研究发现麋鹿鹿角脱落开始时间、发情开始时间、产仔期开始时间逐年提前,而江苏大丰麋鹿种群2004—2010年鹿角脱落开始时间、发情开始时间、产仔期开始时间出现了延后的现象[69],与本研究结果相反,也许与江苏大丰为滨海湿地生境,而北京南海子为内陆湿地生境有关。
种群繁殖特征年节律是一个复杂的规律,受气候、营养、种群密度等诸多因子影响,甚至种群内个体间的相互作用及各饲养区的管理模式也会对它造成影响[17, 60, 66]。本研究的数据为3个年度3种繁殖特征的年节律,时间相对较短,对麋鹿繁殖特征与环境因子之间的关系还需要更长时间的观察研究。
致谢:数据收集中得到北京麋鹿生态实验中心王建昌、包海元的帮助,气象数据分析中得到国家气候中心中国气象局气候研究开放实验室赵俊虎的帮助,特此致谢。
