柳杉半同胞子代测定林树高性状遗传变异研究
2020-11-09陈秋玲
陈秋玲
(宁德市森林资源监测站,福建 宁德 350000)
柳杉(Cryptomeriafortunei)属于杉科(Taxodiaceae)柳杉属(Cryptomeria)[1]。中国柳杉天然林分布于东南部的天目山、武夷山等地[2],江苏、安徽、湖南、湖北、四川等地有人工林栽培[3]。柳杉是我国亚热带地区的优良速生用材林造林树种之一,较适合高海拔或高湿度的沿海山地地区种植[4]。关于柳杉遗传参数的估计已有很多报道[5-9]。黄利斌等[10]通过对包含31个家系的柳杉优树半同胞子代测定林林龄1~3年的树高、胸径性状的研究表明,半同胞子代有明显的性状变异,筛选出3个最优家系,树高、胸径性状遗传增益分别为12.06%和68.81%。李上前[11]分析了一片5年生柳杉一代种子园半同胞子代测定林树高、胸径、材积和冠幅等性状数据,发现除冠幅外各性状家系间差异均极显著,4个性状家系遗传力分别为0.41、0.16、0.36、0.16,单株遗传力分别为0.78、0.60、0.76、0.33。随着柳杉遗传改良工作的不断推进,新一轮育种循环已经开始。本文以营建于福建省霞浦国有林场马洋工区2年生柳杉二代种子园半同胞子代测定林为对象,进行最新世代柳杉幼龄林树高性状遗传潜力及变异幅度的研究,旨在深入了解柳杉遗传变异规律,为进一步的遗传改良及营林生产工作提供可靠的遗传评估信息。
1 材料与方法
1.1 试验地概况
试验地位于福建省霞浦国有林场马洋工区2林班6a小班,地理坐标为26°49′34″ N,119°57′39″ E,属亚热带海洋性季风气候,海拔492~543 m,年平均气温19.5 ℃,年平均降雨量1 411.3 mm。造林地前茬属杉木、柳杉混交林地,土壤为花岗岩发育的山地红壤,土层厚度80~100 cm,立地等级Ⅱ级,坡向东南,坡度15°~26°。
1.2 材料及方法
试验用苗木由霞浦国有林场自育,种子采自本场二代种子园,2016年秋采种,2017年春育苗。共有53个家系及2个对照(原霞浦杨梅岭林场普通苗木和原霞浦水门国有林场普通苗木)。
田间试验采用完全随机区组设计,8次重复,4株小区顺坡排列。造林前经过炼山、烧杂后,进行水平带状整地,带宽1 m。2018年春造林,造林株行距2 m×2.5 m,试验林面积为2 hm2。2019年3月进行补植,数据预处理时剔除了补植单株及补植单株较多的家系,共保留44个家系进行林龄1年和2年树高性状数据的统计分析。
本研究使用R语言进行数据的正态性检验、统计分析及绘图,使用似然比检验单侧检验进行遗传方差组分的显著性检验[12]。统计分析模型为:
y=Xb+Zplotplot+Zaa+e
(1)

通过泰勒展开式估计遗传力、遗传相关的标准误[13]。 单株狭义遗传力:
(2)
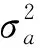
遗传增益计算公式为:
(3)
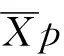
2 结果与分析
2.1 树高性状表型变异
林龄1~2年时树高性状表型表现见表1。可见,林龄1~2年时表型变异系数均较大,林龄1年时的表型变异系数大于林龄2年。图1列出了各家系在林龄1~2年时树高性状的最大值、最小值、中位数、均值、1/4分位数和3/4分位数,可见家系内单株生长量差异很大,两林龄家系间的生长量差异变化也较大。
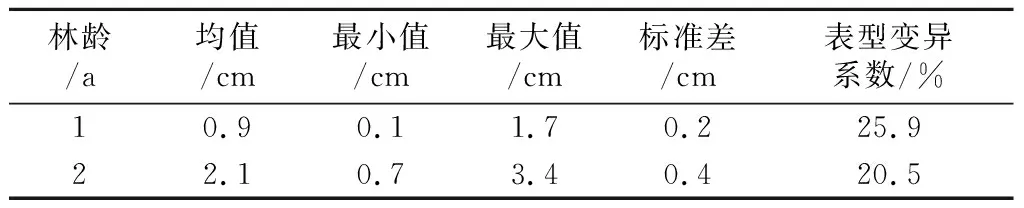
表1 树高性状表型变异

图1各家系树高性状表型表现
2.2 树高性状遗传变异
表2列出了两林龄树高性状加性遗传方差分量、小区方差、单株狭义遗传力、加性遗传变异系数的估计值。两林龄树高性状加性遗传方差、小区方差、单株狭义遗传力均显著。两林龄树高性状单株狭义遗传力均较低,林龄2年时遗传力高于林龄1年时。两林龄树高性状加性遗传变异系数低于表型变异系数,林龄2年时遗传变异系数高于林龄1年时。

表2 树高性状方差分析及遗传参数估算
2.3 各家系遗传增益
林龄1~2年时树高性状各家系遗传增益见图2。林龄2年时各家系遗传增益的最大值、最小值的绝对值均高于林龄1年时;两林龄各家系遗传增益排名相差很大;林龄2年时遗传增益接近0的家系数少于林龄1年时。

图2 各家系遗传增益
3 结论
3.1 柳杉林龄1~2年时树高有较高的生长量和较大的表型变异系数,表型变异系数分别为25.9%和20.5%。
3.2 柳杉林龄1~2年时树高性状加性遗传变异系数较低,分别为3.1%和3.7%。
3.3 柳杉林龄1~2年时树高性状单株狭义遗传力较低,分别为0.067±0.042和0.128±0.058。
3.4 柳杉林龄1~2年时树高性状各家系遗传增益绝对值和排名变化很大。
4 讨论
两林龄树高性状生长量及表型变异系数与国内报道的柳杉种子园子代测定林结果相一致[10, 14-17]。较高的生长量及较大的表型变异为柳杉速生用材林优良家系的筛选提供了选择基础,较大的家系内差异为优良单株的筛选提供了便利。林龄2年时树高性状单株狭义遗传力高于林龄1年时,与Balocchi[18]、Foster[19]等关于火炬松等其他速生用材树种的报道一致。可能是因为随着树木的生长,遗传效应对表型表现的影响逐渐增大。树高性状两林龄单株狭义遗传力分别为0.067±0.042和0.128±0.058,低于国内多数报道的柳杉子代测定林单株狭义遗传力[6, 11, 20, 21]。林龄1~2年时遗传变异系数远低于表型变异系数,可能导致林龄1~2年时进行树高性状表型早期选择的可靠性难以保证。较低的遗传力与加性遗传变异系数,使得林龄1~2年时进行遗传早期选择的价值降低。林龄2年时单株狭义遗传力和加性遗传变异系数估计值分别比林龄1年生时高出1倍和1/5,以及两林龄各家系较大的遗传增益绝对值与排名的变化,可能是因为苗木初植,进行根系修复,表型表现主要受水、肥等环境因素影响,随着林龄的增长,固有的遗传潜力才开始逐渐得到表达。
