联体线虫科(线虫动物门)系统分类研究进展
2020-11-06林施泉王小谷王春生
林施泉,王小谷,王春生*
(1.自然资源部 海洋生态系统动力学重点实验室,浙江 杭州 310012;2.自然资源部 第二海洋研究所,浙江 杭州 310012)
0 引言
联体线虫科(Comesomatidae Filipjev, 1918),隶属于线虫动物门(Nematoda Cobb, 1932),色矛纲(Chromadorea Inglis, 1983),疏毛目(Araeolaimida De Connick and Schuurmans Stekhoven, 1933)[1]。本科线虫物种多样性较高,目前有超过230个真实种[2]。从潮间带、浅海至超过6 000 m的深渊海底均可见其踪迹[3-9]。多项调查结果显示该科线虫密度所占比例可超过40%,是海洋沉积物中优势度较高的一类线虫[10-11]。
早在1979年,JENSEN[12]就对联体线虫科的分类体系做了详细的归纳总结,同时列出了各属的种类检索表,当时该科的分类体系中包含了13属约100个种类。25 a后在FONSECA和BEZERRA[1]的分类体系中,联体线虫科扩大到20个属[含阿卡塔线虫属(ActarjaniaHopper, 1967),但是根据JENSEN[12]1979年的分析,阿卡塔线虫属与萨巴线虫属(SabatieriaRouville, 1903)是同属异名],种类数增加到164种。目前该科包含19个真实属,另有4个疑似属,种类数增加至239种(截止至2019年10月)。2009年至今这10 a间该科已报道的新种数约占近10 a全世界所有新发现海洋线虫总数的1/11(根据Nemys网站统计数据[2]),由此可以看出联体线虫科是当前海洋线虫分类研究中最受关注的一个科。
目前,对联体线虫科最新的综述归纳见于2014年FONSECA和BEZERRA[1]对疏毛目的综述,仅归纳到属这一层次,对30 a来增加了1.3倍的种类并没有作更深入的总结。针对当前世界上对该科的研究现状,有必要对联体线虫科分类系统再次进行梳理,这有助于缩小我国与国外同类研究间的差距。
1 历史沿革
联体线虫科包含3个亚科,分别是联体线虫亚科(Comesomatinae)、萨巴线虫亚科(Sabatieriinae)和矛咽线虫亚科(Dorylaimopsinae)[1]。这3个亚科中最早被发现的是联体线虫属(ComesomaBastian, 1865),该属于1865年由英国著名生物学家BASTIAN[13]发现并建立,以C.vulgare为模式种。FILIPJEV[14]于1918年根据该属直伸型卵巢的特征将其归入他建立的联体线虫亚科 (Comesomini),从属于单宫线虫科(Monhysteridae)。当时的联体线虫亚科仅包含联体线虫属、萨巴线虫属和异萨巴线虫属(Parasabatieriade Man, 1907)3个属,实际上异萨巴线虫属与萨巴线虫属应为同属异名[15]。随后在1933年,DE CONINCK和SCHUURMANS STEKHOVEN[16]将联体线虫亚科提升为科,并将该科从单宫线虫总科(Monhysteroidea)移入到色矛线虫总科内(Chromadoroidea)。
萨巴线虫属于1934年被 FILIPJEV[17]定为模式属,归入新成立的萨巴线虫亚科,他将萨巴线虫亚科归入条线虫科(Linhomoeidae)。当时萨巴线虫亚科还包括克莱西亚线虫属(KreisiaAllgén, 1929)、 阿莱默线虫属(AlaimonemaCobb, 1920)、 皮索线虫属(PepsonemaCobb, 1920)、 中齿线虫属(MesonchiumCobb, 1920)、矛咽线虫属(DorylaimopsisDitlevsen, 1918)和费氏线虫属(FilipjevaDitlevsen, 1928)。这些属的分类地位后来多有变动,例如:COBB[18]认为皮索线虫属与矛咽线虫属应该是同属异名;JENSEN[12]认为中齿线虫属与矛咽线虫属也是同属异名;LORENZEN[19]分析指出克莱西亚线虫属为可疑属,阿莱默线虫属应归入链环线虫科(Desmodoridae),费氏线虫属应归入希阿利线虫科 (Xyalidae)。
DE CONINCK[20]在1965年建立了联体线虫科的第3个亚科——矛咽线虫亚科,他将原属于鳗状线虫科(Anguillulidae)[21],后被归入条线虫科萨巴线虫亚科[17]的矛咽线虫属作为该亚科的模式属,同时还包含希线虫 属(XinemaCobb, 1920)。此时 DE CONINCK已将矛咽线虫亚科、联体线虫亚科和萨巴线虫亚科 3个亚科都归入联体线虫科中,他还指出矛咽线虫亚科与其他亚科的主要区别在于其口腔为锯齿状[20]。
GERLACH和RIEMANN[22]在1973年建立棘刺线虫亚科(Acantholaiminae),以棘刺线虫属(AcantholaimusAllgén, 1933)为模式属,并将该亚科并入 DE CONINCK建立的联体线虫科分类体系中[20]。然而 JENSEN[12]在1979年将棘刺线虫亚科移出联体线虫科,主要依据为前者种类具有复杂的口腔结构、食道有肌肉束结构、食道中缺乏侧管腔、交合器无引带、化感器呈半圆形以及表皮有很深的斑点等特征与联体线虫科有很大的差异。
联体线虫科的分类地位一直在不断变化中,在建立之初归属于单宫线虫总科[14],DE CONINCK 和 SCHUURMANS STEKHOVEN(1933)[16]、WIESER(1954)[23]、DE CONINCK (1965)[20]等都将联体线虫科归入色矛线虫亚目,色矛线虫总科中。DE CONINCK[20]在1965年描述的色矛目特征之一是具有反折的卵巢,包含了联体线虫科、微咽线虫科和色矛线虫科等。但是根据 JENSEN[12]的研究结果,联体线虫科多数种类为直伸型卵巢,仅霍帕线虫属 (HopperiaVitiello, 1969) 中有2个种类有反折的卵巢。为此,ANDRSSY[24]在色矛线虫亚目内新建立了一个新的总科——联体线虫总科 (Comesomatoidea),包含了联体线虫科(含联体线虫亚科和萨巴线虫亚科)和矛咽线虫科 (Dorylaimopsidae,包含矛咽线虫亚科),并将该总科归入杯咽线虫亚目中(Cyatholaimina),与该亚目中其他种类的最大区别在于雌性生殖系统、交合刺和食道结构特征。LORENZEN[19]赞同 FILIPJEV[14, 17]的观点,依据联体线虫科直伸型卵巢特征将其归入单宫目,而矛咽线虫科被降为亚科依旧并入联体线虫科内。PLATT[25]在1985年依据联体线虫科表皮斑点和多螺旋化感器的特征将其归入色矛目。MALAKHOV[26]根据联体线虫科化感器为多螺旋结构的特征将联体线虫科归入疏毛目。分子生物技术的发展也不断改变对该科的认识,如LITVAITIS et al[27]2000年根据 28S rDNA 测序结果认为联体线虫科应归入色矛目。SHARMA et al[28]根据 18S rDNA 测序和聚类分析结果,认为联体线虫科应归入单宫目。MELDAL et al[29]根据 SSU rDNA序列分析的结果也认为联体线虫科亲缘关系与单宫目线虫更近。FONSECA和 BEZERRA[1]认为目前联体线虫科的分类地位尚不明晰,仍有待深入研究,暂时先按 LORENZEN[19]在1981年建立的分类体系将其归入疏毛目轴线虫总科内。目前在 WORMS 网站线虫的分类体系中也是如此归属[30]。
2 形态特征
该科种类形态特征表现为表皮环化,多数种类的体环具有斑点,常有侧分化现象,有的种类体表有弱角质化的开孔。化感器多为螺旋结构。头部感觉毛通常排列呈6+6+4分布。口腔常分成2个部分,前部为圆形、杯形或狭窄;后部狭窄,弱角质化,或呈圆柱形,或呈圆锥形;两部分之间常见有齿状突起。食道有3个边缘管,其背侧食道腺开口于口腔底部。排泄孔位于神经环后面。雌性有2个直伸型的卵巢,具有纳精囊。雄性具有2个异向排列的精巢,交合刺弯曲,引带突朝背尾向。肛前附器通常呈乳突状。尾尖略微膨大,未角质化,具有3根尾刚毛。各部分结构的具体特征如下文所述。
2.1 体长、颜色及外观
目前已知的联体线虫科多数种类体长在1.5~3.5 mm之间,也有例外,如Sabatieriapisinna体长仅 0.7 mm,而德氏萨巴线虫 (S.demani)体长可达 8 mm。大部分种类虫体无颜色,少数种类如大化感器长颈线虫 (CervonemamacramphisJensen, 1979)虫体呈现黄绿色。该科线虫虫体通常呈圆柱形,两端逐渐变细,但长颈线虫属(CervonemaWieser, 1954)的颈部狭长;唇部通常如坐垫形状,但是长颈线虫属、雷曼线虫属(LaimellaCobb, 1920)和皮氏线虫属(PierrickiaVitiello, 1970)种类的唇部发育不完全;尾部呈锥柱形,尾尖,无角质化结构,常有 3根尾刚毛,尾孔突起且对称。
2.2 表皮
该科各属线虫表皮形态不尽相同,或有横纹,或覆有或大或小的斑点。有的种类表皮上这些圆点均匀分布,如管腔线虫属(VasostomaWieser, 1954)和少部分萨巴线虫属种类,其他大多数都有侧分化现象,矛咽线虫属和少部分萨巴线虫属种类的侧分化出现规则纵向排列的粗斑点。异毛联体线虫属和一些萨巴线虫属的表皮光滑无装饰物。长颈线虫属和雷曼线虫属表皮有横纹,如果放大数倍可发现其实是规则的圆点。在颈部和尾部侧分化现象更加明显。矛咽线虫属部分种类纵向线数量存在雌雄差异。在透明矛咽线虫(Dorylaimopsispellucida)、点矛咽线虫(D.punctata)、 比利时异中齿线虫和SabatieriamigransJensen & Gerlach, 1977种类的表皮上还发现有弱角质化的圆孔结构。
表皮侧分化是矛咽线虫亚科和萨巴线虫亚科区分属间差异的1个重要特征,也是区别与矛咽线虫属相近属的1个重要特征。
2.3 体刚毛
体刚毛通常排列成4个纵列,位于背侧和腹侧,一些种类(如矛咽线虫属和萨巴线虫属)的雄性尾部腹侧常有额外的纵列。位于颈部的体刚毛(颈刚毛)比其他体刚毛长(如部分萨巴线虫属种类),或者 2~4根刚毛聚成一簇(如一些长颈线虫属和萨巴线虫属种类)。体刚毛特征仅能用作种水平的鉴定。
2.4 化感器
联体线虫科化感器通常为螺旋状,一般有3~4圈,Sabatieriarota仅有 1圈,长颈线虫属种类最多的有 7圈。化感器直径通常为相应体宽的60%~70%,但对于某些萨巴线虫,这一比例可达到90%。大部分种类化感器前缘位于头刚毛附近或稍后一点,但长颈线虫属种类化感器大致位于2倍头直径的位置。
多螺旋形化感器是联体线虫科和杯咽线虫科特有的特征,色矛目的Spiliphera属和筛咽线虫属 (Ethmolaimusde Man, 1880)也有,但是不如前2个科的明显。
2.5 头部刚毛
联体线虫科头部刚毛通常排列成3环,6根内唇感觉毛呈乳突状,6根外唇感觉毛和4根头感觉毛呈刚毛状。6根外唇感觉毛通常为刚毛状,在侧面的两根显著大于其余4根,位于第2圈和第3圈感觉毛之间(例如雷曼线虫属、联体线虫属和筛咽线虫属的部分种类)。杯咽后联体线虫(Metacomesomacyatholaimoides)的第2圈和第3圈感觉毛并排成等长的1圈刚毛。第3圈感觉毛通常长于第2圈,但长颈线虫属、后联体线虫属(Metacomesoma)和皮氏线虫属的种类是等长的。Hopperiamassiliensis和Laimellaminuta两个种类所有感觉毛全部退化成乳突状。长颈线虫属种类在头部稍后位置还有1圈或多圈刚毛,但在幼体期没有发育。
2.6 口腔
口腔的分化程度是该科的一个重要特征。如果有出现分化,通常为圆形、杯形或者为狭长的腔室,有时口腔前后之间有凸出物。该科的后腔室角质化程度较弱,像折叠的管子(如萨巴线虫亚科)或为圆锥形(矛咽线虫亚科)。矛咽线虫亚科部分种类口腔形态类似条线虫科的种类,如Paracomesomainaequale的齿状结构和条线虫科的咽齿线虫(OdontophoraBütschli, 1874)的结构就很相似,但这可能只是一个趋同现象。
长颈线虫属口腔很小。皮氏线虫属部分种类口腔与长颈线虫属类似,部分种类口腔前部为球形未分化,该科其余线虫的口腔均分化为2部分,但形态和角质化程度不尽相同。萨巴线虫属、弧头短尾线虫属(ScholpaniellaSergeeva, 1972)属和后联体线虫属口腔前部为杯状,弱角质化;后部狭长,无角质化,像一个折叠的管子;有时在前后2个部分之间还有凸起物。雷曼线虫属口腔后面部分很短,高度角质化,变为食道最前端管腔。典型的萨巴线虫属和联体线虫属类似,但口腔前部分更小,2个部分之间的凸起物角质化程度明显。矛咽线虫亚科口腔后半部分宽大,有高度角质化的腔壁,或为圆锥形(如异中齿线虫属和管腔线虫属),或为圆柱形(如矛咽线虫属、霍帕线虫属和后萨巴线虫属)。该亚科口腔前后两部分边界有明显的角质化凸起,后萨巴线虫属除外。比利时异中齿线虫(ParamesonchiumbelgicumJensen, 1976)种类口腔前段靠后部分有角质化凸起,还分化成一排小齿。点矛咽线虫在前庭和短小的前部分口腔之间还分化为不规则边缘。异毛联体属(Paracomesoma)线虫口腔后半部分有高度角质化的腔壁(如同矛咽线虫属),伴有3个或者6个凸出物。
口腔底部常见弱角质化突出物,作用未明。JENSEN[12]通过比较该科6个种类以及单宫目2个属[线形线虫属(LinhomoeusBastian, 1865)和反轮线虫属(Anticyclus)]和RIEMANN et al[31]在1977年的研究结果,都认为这个结构是同源的,作用可能是连接食道腺的出口。
2.7 食道
该科种类的口腔被食道包绕,有时食道末端膨大,在长颈线虫属和Pierrickiaaequalis(Gerlach, 1956) Vitiello, 1970种类就有食道球,食道管腔类型为三射状食道管。
食道腺通常为颗粒状,其中侧腹面的食道腺开口于神经环前端;位于背侧的食道腺靠近角质化的口腔凸起物附近。由于棘刺线虫属和三齿线虫属(Tridentellia)的食道缺乏侧管腔,因此JENSEN[12]认为这2个属更偏向于杯咽线虫科,而非联体线虫科。
2.8 排泄系统
该科线虫排泄系统包含1个单核腺细胞(焰细胞),位于腹侧,在贲门后,有时焰细胞明显与体腔相连,它与壶腹通过管道相连,代谢物到壶腹后经过1条狭长的管道到达角质化开孔,排出体外。
2.9 生殖系统
该科雌体具有2个直伸的卵巢,大多雌孔位于虫体中部。具有颗粒状的阴门腺。阴道一般朝向背腹侧,但有的种类 (例如SabatieriamigransJensen & Gerlach, 1977)朝向前后侧。雌性生殖系统包含2个分支(双卵巢型),2个卵巢异向排列,直伸型,少数种类为反折型,具有纳精囊。地中海矛咽线虫(DorylaimopsismediterraneaGrimaldi-De Zio, 1968)种类具有双性特征,该种类雌性发育有雄性第1和第2性征。
雄性性腺一般为2个异向直伸型的精巢。
该科雄性典型的交合刺特征可参考萨巴线虫属:交合刺弯曲,近端膨大,远端尖锐,有时末端膨大或有凸出物;有的种类引带有侧分化和腹侧分化;引带突朝向背后方。长颈线虫属种类的交合刺角质化程度较低;矛咽线虫亚科种类的交合刺或为纤细略弯曲状或为弯曲状,矛咽线虫属部分种类的交合刺还分成前后两部分,中间通过关节相连。部分种类引带还有侧向分布的导片,在腹侧末尾弯曲。引带突朝向背尾端,与体轴平行。Scholpanialla属种类与萨巴线虫类似,区别在于引带突很小,甚至退化成筷子状朝向尾部。联体线虫亚科的一些种类交合刺伸长且角质化明显;引带小且高度角质化或似管状弱角质化;引带突单个,与交合刺平行。后联体属线虫无引带突。
该科很多种类具有5~40余个腹侧肛前附器,大多数类似乳突状或为较模糊不易观察的小孔。
3 分类系统
LORENZEN[19]1981年对自由生活海洋线虫的系统发育做了详细研究,在他构建的体系中,联体线虫科的单系特征仅有化感器螺旋数至少有2.5圈这一点,包含14个属。根据FONSECA和BEZERRA[1]在2014年的分类体系,联体线虫科包含20个属,但是阿卡塔线虫属与萨巴线虫属为同属异名[12],因此实际上联体线虫科包含19个真实属239个种类,另有4个疑似属。这19个属分别归属于3个亚科:联体线虫亚科、矛咽线虫亚科和萨巴线虫亚科,以联体线虫属为模式属。3个亚科的主要区别如表1所示。

表1 联体线虫科亚科检索表Tab.1 Key to the subfamily of Comesomatidae
目前,我国已报道的联体线虫科共有33种[4-6, 9, 32-43],隶属于3个亚科9个属。
3.1 联体线虫亚科(Comesomatinae Filipjev, 1918)
联体线虫亚科种类表皮有环纹或斑点;口腔后部呈圆柱形、膨大或塌陷形,前后口腔交界处有突起物;交合刺长;引带突呈管状,带有1个坚硬的中板或仅有中板,引带突纤细[1]。口腔结构和长交合刺的特征是本亚科种类区别于其它亚科的重要特征。该亚科以联体线虫属(ComesomaBastian, 1865)为模式属,目前包含4个属,31个种类。该亚科各属种类特征见图1,分属检索表见表2。

表2 联体线虫亚科分属检索表Tab.2 Key to the genera of Comesomatinae

图1 联体线虫亚科代表种头部和尾部示意Fig.1 Anterior end and tail structure of representativespecies of subfamily Comesomatinae(a—Comesoma arenae[44];b—德氏似联体线虫[45];c—Metacomesoma cyatholaimoides[23];d—微小异毛联体线虫[46]。)(a—Comesoma arenae[44];b—Comesomoides deconincki[45];c—Metacomesoma cyatholaimoides[23];d—Paracomesoma minor[46].)
3.1.1 联体线虫属(ComesomaBastian, 1865)
头部感觉毛排列成 3圈,外唇刚毛比头刚毛长(图1a),颈部刚毛排列成 1圈或多圈,每1圈 4根刚毛;口腔后部塌陷,弱角质化,在前部和后部交界处有 3个很小且钝的凸起;交合刺细长;引带突盘状或不明显[1]。联体线虫属是联体线虫科中最早建立的1个属,以C.vulgareBastian, 1865为模式种,目前记录有11个种,种类检索表见表3。

表3 联体线虫属种类检索表Tab.3 Key to the species of Comesoma
3.1.2 似联体线虫属(ComesomoidesGourbault, 1980)
体表有斑点,侧分化点不规则排列;头部感觉毛排列成3圈,头刚毛最长(图1b);化感器环绕2圈;口腔前部呈杯形,后部弱角质化呈管状;交合刺延长;无背部引带突;具有肛前附器。似联体线虫属目前仅有1个种类:德氏似联体线虫(ComesomoidesdeconinckiGourbault, 1980)。
3.1.3 后联体线虫属(MetacomesomaWieser, 1954)
头部化感器排列成2圈;外唇刚毛与头刚毛等长(图1c);口腔后部塌陷,极弱角质化;交合刺长;引带呈管状,包绕住交合刺后部,一般无引带突[34]。本属与模式属(联体线虫属)的主要区别在于外唇刚毛和头刚毛排列方式(前者排列呈1圈,后者分成2圈)以及有无颈部刚毛(前者无颈部刚毛)。
目前该属有2个种类,除了1954年WIESER[23]发现模式种外,2018年HUANG和 HUANG[34]在我国东海发现大化感器后联体线虫(M.macramphida),它与模式种的区别主要为前者头刚毛较短(1.5 μm),化感器圈数较多(4.5圈),交合刺长度为肛径的4.7倍。大化感器后联体线虫与德氏似联体线虫形态也很相似,主要区别在于后者表皮有侧分化,有引带突和肛前附器等特征,这些大化感器后联体线虫都不具备。
3.1.4 异毛联体线虫属(ParacomesomaSchuurmans Stekhoven, 1950)
异毛联体线虫属是联体线虫亚科种类最多的1个属,由SCHUURMANS STEKHOVEN[47]在1950年建立,当时描述的种类Paracomesomacoronatum事实上是另一个种Paracomesomadubium的同种异名[23],而Paracomesomadubium原名为Comesomadubia,SCHUURMANS STEKHOVEN[47]认为该种与Comesoma其他种类差异明显,口腔有3颗显著的牙齿(图1d),因此新建立异毛联体线虫属。HOPPER[48]认为该属无模式种不是有效属,但是又将雷曼线虫属的2个种类Laimellahexasetosa和Laimellaquadrisetosa归入本属中,这2个种类的雄性样品形态与雷曼线虫属特征不吻合;同时他也将Vasostomalongispiculum归入异毛联体线虫属。随后,HOPE和MURPHY[49]在1972年将Paracomesomadubium(Filipjev, 1918)认定为该属的模式种。该属主要特征如下:头部感觉毛排列呈3个分开的圈,有时外唇刚毛的侧边感觉毛比其它4根感觉毛长,并且位于第2圈和第3圈之间;口腔后部角质化,呈圆柱形,口腔内有3个或6个齿状凸起。异毛联体线虫属与联体线虫属的主要区别在于没有颈部刚毛。目前异毛联体线虫属已纪录有17个种类,种类检索表见表4。
3.2 矛咽线虫亚科(Dorylaimopsinae de Coninck, 1965)
该亚科虫体后部表皮通常有侧分化现象。体前感觉毛显著排列成 3圈。口腔高度角质化,前部狭窄,有时在口腔前庭有凸起物;口腔后部膨大,呈圆锥形或圆柱形,腔壁高度角质化,有3个棘状突起[1]。口腔结构是该亚科种类区别于其他2个亚科种类的最重要特征。目前该亚科包含 8个属,59个种类,以矛咽线虫属为模式属,该亚科各属种类特征见图2,分属检索表见表5。

表4 异毛联体线虫属种类检索表Tab.4 Key to the species of Paracomesoma

图2 矛咽线虫亚科代表种头部和尾部示意Fig.2 Anterior end and tail structure of representativespecies of subfamily Dorylaimopsinae(a—越南歪头线虫[50]; b—Dorylaimopsis gerardi[11];c— Expressonema grandulata[51]; d—六齿霍帕线虫[52];e— Kenyanema monorchis[11]; f:—Metasabatieria primigena[53];g:—Paramesonchium mombasi[11]; h—长刺管咽线虫[37]。)(a—Asymmelaimus vietnamicus[50]; b—Dorylaimopsis gerardi[11];c— Expressonema grandulata[51]; d—Hopperia hexadentata[52];e— Kenyanema monorchis[11]; f—Metasabatieria primigena[53];g—Paramesonchium mombasi[11]; h—Vasostoma longispicula[37].)

表5 矛咽线虫亚科分属检索表Tab.5 Key to the genera of Dorylaimopsinae
3.2.1 歪头线虫属(AsymmelaimusTu, Vu Thanh, Smol & Vanreusel, 2008)
虫体前部及口腔不对称。表皮点纹纵向排列,有侧向分化,中间1列或2列大的点纹,侧边还有2列点纹位于食道区域,1列位于虫体中部。化感器螺旋形,2圈,很大。口腔迁移至背面,前部呈杯型,带3个小齿;后部呈管状,弱角质化。交合刺大且弯曲,引带突朝向尾部。目前该属仅有1个种,发现于越南哈龙湾,因发现地而被命名为越南歪头线虫 (AsymmelaimusvietnamicusTu, Vu Thanh, Smol & Vanreusel, 2008)。
3.2.2 矛咽线虫属(DorylaimopsisDitlevsen, 1918)
表皮侧分化为纵向排列的粗大斑点,斑点在体前部为3纵列,在后部变为2纵列,到肛前时又变为3个粗大的纵列,斑点列与列之间的距离也随之变化;6根唇部感觉毛短,6根头部感觉毛较短,4根头部感觉毛较长;口呈圆柱形,前端有3个齿;化感器螺旋状,接近3圈;食道逐渐膨大,无食道球,心形食道与肠连接;排泄细胞和排泄管开口明显;雄性精巢2个,对向分布,直伸型;弓形交合刺2根,对称;引带突朝向背尾侧;肛前附器为14个由前向后相互之间距离缩小的乳突;雌性卵巢2个,为直伸型;尾部呈锥柱形。该属以点矛咽线虫(DorylaimopsispunctataDitlevsen, 1918)为模式种。根据WORMS网站显示矛咽线虫属包含25个种类[30],但HUANG et al[35]2018年在我国胶州湾发现的D.heteroapophysis尚未被收录,实际目前应该有26种。本属种类检索表见表6。

表6 矛咽线虫属种类检索表Tab.6 Key to the species of Dorylaimopsis
3.2.3 艾斯普里索线虫属(ExpressonemaSmolyanko & Belogurov, 1991)
表皮有横向排列的点纹,无侧边分化;口腔呈圆锥型,有9颗小刺齿;交合刺细长,末端略膨大;引带退化,引带突小,位于背面,与交合刺平行。该属目前仅有1个种类:ExpressonemagranulateSmolyanko & Belogurov, 1991。该种雌体和雄体腺细胞数量不同,雄性精子有二形性[51]。
3.2.4 霍帕线虫属(HopperiaVitiello, 1969)
霍帕线虫属是由VITIELLO[54]在1969年建立,以HopperiamassiliensisVitiello, 1969为模式种。霍帕线虫属表皮具有斑点,侧边分化为不规则排列大点;口腔后部呈圆柱形,与前部交接处有3个刺状凸起;卵巢部分种类为折叠型;交合刺呈弯曲状,短;引带突朝向尾部或为背尾向。霍帕线虫属目前有17个种类,种类检索表见表7。

表7 霍帕线虫属种类检索表Tab.7 Key to the species of Hopperia
3.2.5 肯尼亚线虫属(KenyanemaMuthumbi, Soeta-ert & Vincx, 1997)
该属以发现地肯尼亚为属名[11],归入该亚科主要是由于其交合刺特征与本亚科吻合。肯尼亚属线虫表皮有密集的斑点,有时可见环带,有侧分化,斑点不规则分布;化感器有 1.5~2个螺旋;口腔呈管状,没有齿;单卵巢,直伸型;雄性单精巢,伸向前部;交合刺巨大,弯曲,中间带有薄板;引带有细长的引带突;尾短,呈圆锥或圆柱形,末端膨大。肯尼亚属种类头部感觉毛与矛咽线虫亚科其它属差异显著,也是联体线虫科内唯一一个生殖腺为单精巢型的属。目前该属仅有1个种,即模式种KenyanemamonorchisMuthumbi, Soetaert & Vincx, 1997。
3.2.6 后萨巴线虫属(MetasabatieriaTimm, 1961)
表皮无侧分化;口腔后部呈圆柱形,无角质化;交合刺弯曲;引带突朝向背尾部。后萨巴线虫属原本归属于萨巴线虫亚科,JENSEN[12]依据其口腔特征将其转入矛咽线虫亚科。目前后萨巴线虫属仅记录有1个种类,即模式种MetasabatieriaprimigeraTimm, 1961。
3.2.7 异中齿线虫属(ParamesonchiumHopper, 1967)
HOPPER[48]根据 WIESER[23]在 1954 年描述的1个疑问种Laimellaserialis特征建立本属,并将该种定为模式种。本属线虫表皮侧分化,斑点横向排列或纵向排列至全身;口腔后部呈圆锥形,有 3个隆起部分,每个隆起部分都有尖凸起;交合刺弯曲;引带突朝向背尾部。本属与后来都并入矛咽线虫属的阿卡塔线虫属和中齿线虫属形态最相似,主要区别在于前者咽部为圆锥形[48]。本属目前发现有3个种,种类检索表见表8。

表8 异中齿线虫属种类检索表Tab.8 Key to the species of Paramesonchium
3.2.8 管腔线虫属(VasostomaWieser, 1954)
管腔线虫属由WIESER[23]在1954年建立,以螺旋管腔线虫 (V.spiratumWieser, 1954)为模式种。管腔线虫属表皮无侧分化现象;口腔后部呈圆锥形,有 3个细小的尖突起;交合刺弯曲;引带突朝向背尾部。管腔线虫属与矛咽线虫属最相似,主要区别在于后者表皮侧分化为纵向排列的粗斑点。目前管腔线虫属已记录有 8种,最新纪录为 ROSLI et al[55]在新西兰希古朗基南部海底峡谷中发现的六齿管腔线虫(V.hexodontium)。本属种类检索表见表9。

表9 管腔线虫属种类检索表Tab.9 Key to the species of Vasostoma
3.3 萨巴线虫亚科(Sabatieriinae Filipjev, 1934)
萨巴线虫亚科包含7个属,以萨巴线虫属为该亚科内模式属。PLATT[25]认为该亚科的种类无共通的特征。除了具有该科一般特征外,萨巴线虫亚科种类口腔一般弱角质化,口腔后部为折叠的管状;交合刺末端膨大;引带突成对或单个。雷曼线虫属、长颈线虫属和毛萨巴线虫属(Setosabatieria) 3个属种类表皮有环纹;该亚科大多种类表皮具有小斑点,但雷曼线虫属种类很少有小斑点。萨巴线虫亚科目前已发现7个属,该亚科各属种类特征见图3,分属检索表见表10。
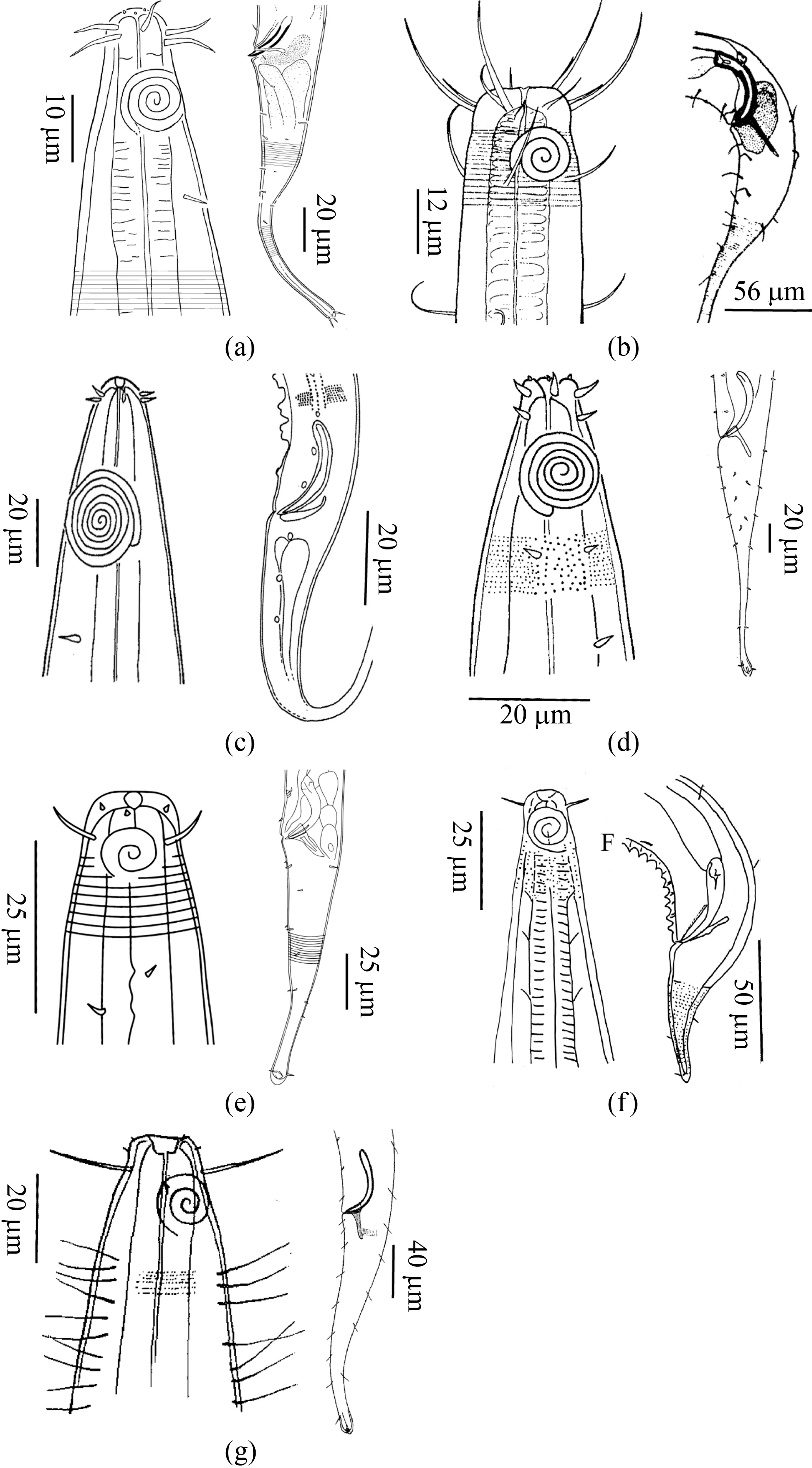
图3 萨巴线虫亚科代表种头部和尾部示意Fig.3 Anterior end and tail structure of representativespecies of subfamily Sabatieriinae(a—东海长颈线虫[56]; b—长尾雷曼线虫[57]; c—林奈微咽线虫[58];d—Pierrickia decasetosa[58]; e—Sabatieria annulata[59];f—Scholpanialla pontica[60]; g—Setosabatieria chitwoodi[23]。)(a—Cervonema donghaensis[56]; b—Laimella longicauda[57];c—Minolaimus lineatus[58]; d—Pierrickia decasetosa[58];e—Sabatieria annulata[59]; f—Scholpanialla pontica[60];g—Setosabatieria chitwoodi[23].)

1口腔前部呈球形或杯型2…………………………………………………………………………………………………………………………口腔前部小4………………………………………………………………………………………………………………………………………2无颈刚毛或很短3…………………………………………………………………………………………………………………………………有颈刚毛 Setosabateria…………………………………………………………………………………………………………………………3引带突明显,朝向尾部 Sabateira………………………………………………………………………………………………………………引带突小,朝向背部 Scholpanialla………………………………………………………………………………………………………………4化感器小于4圈;外唇刚毛比头刚毛长Laimella………………………………………………………………………………………………化感器大于 4圈;外唇刚毛与头刚毛等长5……………………………………………………………………………………………………5食道呈圆柱形;化感器位置较靠前Pierrickia…………………………………………………………………………………………………食道末端膨大;化感器位置靠后6………………………………………………………………………………………………………………6表皮无侧分化Cervonema………………………………………………………………………………………………………………………表皮纵向侧分化Minolaimus……………………………………………………………………………………………………………………
3.3.1 长颈线虫属(CervonemaWieser, 1954)
长颈线虫属由WIESER[23]在1954年建立,以CervonemaallemetricumWieser, 1954为模式种。虫体细长,表皮弱环化;颈部狭长;内唇刚毛不明显,外唇刚毛和头刚毛等长;化感器4~7圈,螺圈狭窄,位置相对远离前部;口腔小,咽部呈圆柱形,食道后部形成肌肉束和食道腺,通常膨大,但没有形成食道球;交合刺弱角质化,短,直或略弯,长度约等于肛径;引带发育不完全或不可见,某些种类有引带突朝背部或背尾向;肛前附器小乳突状或无;尾锥呈柱形。目前该属纪录有19个种类(WORMS尚未收录CervonemalongispiculaHuang, Jia & Huang, 2018)。本属种类检索表见表11。
3.3.2 雷曼线虫属(LaimellaCobb, 1920)
雷曼线虫属由COBB在1920年建立[61]。雷曼线虫属虫体主要特征包括表皮有条纹或者密集的横向排列的斑点;外唇刚毛和头刚毛靠近;口腔发育不完全;食道前部明显角质化;尾锥呈柱形,末端为鞭毛状。雷曼线虫属目前报道有11个种类,以长尾雷曼线虫(LaimellalongicaudaCobb, 1920)为模式种。本属种类检索表见表12。

表12 雷曼线虫属种类检索表Tab.12 Key to the species of Laimella
3.3.3 微咽线虫属(MinolaimusVitiello, 1970)
微咽线虫属是VITIELLO[58]在1970年建立的,以M.cervoide为模式种。该属种类虫体前端呈圆锥形;表皮侧分化,有3列纵向排列的斑点;化感器大,多螺旋结构;口腔非常小,且无齿;尾纤细呈丝状。到目前为止,属内只有2个已知种,分别是M.lineatusVitiello, 1970和M.cervoidesVitiello, 1970,其中M.cervoides种只描述了雌体。M.cervoides与M.lineatus的主要区别在于前者尾相对较小,化感器相对较大。黄冕[62]的硕士论文中还描述了在我国海南岛发现的1个新种:多辅器微咽线虫 (M.multisupplementus)。该种与其他2个种的区别特征在于其角质化的表皮侧面有3排点状侧装饰;螺旋状化感器有7圈;咽呈柱状,无咽球;交合刺具有中心薄板;引带与交合刺平行,无引带突;有29个杯状肛前辅器;尾部有较长的丝状部分。
3.3.4 皮氏线虫属(PierrickiaVitiello, 1970)
皮氏线虫属由VITIELLO[58]在1970年建立,以PierrickiadecasetosaVitiello, 1970为模式种,VITIELLO还将原属于后联体线虫属的MetacomesomaaequaleGerlach, 1956归入皮氏线虫属中,加上GOURBAULT[45]1980年在大西洋发现的新种P.vitielloi,目前本属共有3个种类。本属种类表皮有斑点;化感器大于4圈,位于虫体前部;外唇刚毛和头刚毛等长;口腔很小;若有引带突则朝向背尾向(在P.aequalis种类中引带和引带突缺失)。本属种类检索表见表13。

表13 皮氏线虫属种类检索表Tab.13 Key to the species of Pierrickia
3.3.5 萨巴线虫属(SabatieriaRouville, 1903)
萨巴线虫属是联体线虫科中种类最多的1个属,它由ROUVILLE[63]在1903建立,以SabatieriacettensisRouville, 1903为模式种。虽然WORMS网站目前统计有105个种类,但是根据ROSLI et al[55]2014年报道,本属有69个真实种;LEDUC[64]支持ROSLI et al[55]的观点,并在新西兰陆架区发现1个新种S.megadenaLeduc, 2017;随后GUO et al[5]2018年在福建红树林湿地中发现S.conicosetaGuo, Chang & Yang, 2018;FU et al[3]2019年也在新西兰陆架区发现另一种萨巴线虫S.articulataFu, Leduc & Zhao, 2019;ZHAI et al[43]在南海发现新种S.minorZhai, Wang & Huang, 2019。综上,目前该属应该有73个真实种。
萨巴线虫属虫体表皮有环纹或者有横向排列的斑点,可能有侧向分化;头部化感器排列成3圈;头刚毛长于外唇刚毛;口腔前部呈球状或杯状,后部缩窄成塌陷状管,弱角质化,有时在前后交接处有小凸起;雄性排泄系统有2个额外的位于侧腹部的单核腺体,远离贲门;交合刺近端通常膨大;引带突通常直接朝向尾部或背尾向。
萨巴线虫属也是联体线虫科中分类最复杂的1个属。根据 PLATT[25]的观点,萨巴线虫属种类可以分为以下 5组:
(1) praedatrix组:此类群中大多种类表皮具侧分化现象,斑点大且排列稀疏;化感器多为2.5~3.0圈,偶见2或4圈;肛前附器呈管状或者孔状;引带突呈笔直型。目前 praedatrix类萨巴线虫共有46种。
(2)armata组:与 praedatrix类似,除了以下几点:头刚毛和颈刚毛更长(大于1倍头直径);虫体通常纤细(体长与体宽比值大于65%);化感器通常为3圈,偶见4圈;肛前附器常为管状。目前armata类萨巴线虫共有6种。
(3)pulchra组:颈刚毛短而粗,成对排列;化感器多为2.5~3.5圈;肛前附器很明显,但数量较少(通常5~9个);引带有中间板。目前pulchra类萨巴线虫共有9种。
(4)celtica组:化感器通常为2.5~3.0圈;虫体相对肥大;引带突呈弯曲型;肛前附器明显,数量为9~27个不等。目前celtica类萨巴线虫共有5种。
(5)ornata组:化感器通常为2.5~3.0圈。与celtica组类似,但该类群的肛前附器在后部更密集。目前ornata类萨巴线虫共有5种。
除了上述5组外,另有2种萨巴线虫(S.dispunctata和S.megadena)无法归入上述 5个类群中。它们区别于其他5组种类的主要特征在于体表无斑点和口腔非常小。萨巴线虫属中有3个种类(S.dodecaspapillata,S.longisetosa和S.mortenseni)体表也没有斑点。S.dispunctata与这些种类的区别在于头刚毛短(0.6~0.7倍相应体宽),化感器螺旋为 4.5圈,无成对排列的颈刚毛,无肛前附器等特征[55]。S.megadena与这些种类的区别在于化感器螺旋数仅有2圈;肛前附器较少,仅有6个[64]。
上述73个萨巴线虫种类的形态特征测量参数如附表1所示。
3.3.6 弧头短尾线虫属(ScholpaniellaSergeeva, 1972)
弧头短尾线虫属由SERGEEVA[60]在1972年建立,以ScholpaniallaponticaSergeeva, 1972为模式种,目前仅纪录1种。该属主要特征为头呈弧形;尾短,呈圆锥形;表皮弱环纹,具斑点,未侧分化;头刚毛排列成 1圈;体刚毛遍布全身;口腔呈杯形,食道末端弱膨大;交合刺短,呈弧形,有一软腭;引带为瘦弱型,无凸起。
3.3.7 毛萨巴线虫属(SetosabatieriaPlatt, 1985)
毛萨巴线虫属由 PLATT[25]在1985年建立,以 DE MAN[65]在1922年发现的Sabatieriahilarula为模式种。毛萨巴线虫属与阿卡塔线虫属形态最相似,区别在于前者种类表皮光滑无斑点,在侧面有几排长的颈部刚毛。WORMS目前收录有11个种类,还未收录CHEN et al[66]在2014年报道的新加坡毛萨巴线虫(S.singaporensisChen & Shabdin, 2014)以及HUANG et al[42]2019年在南海新发现的微小毛萨巴线虫(SetosabatieriaminorHuang, Xu & Huang,2019)。但是WORMS收录的Setosabatieriachitwoodi,原种名为SabatieriachitwoodiWieser, 1954,PLATT[25]在1985年认定该种与Setosabatieriahilarula(de Man, 1922)为同种异名,在WORMS收录的萨巴线虫属名录中也指出这一点,因此目前该属种类数应该为 12 个。本属种类检索表见表14。

表14 毛萨巴线虫属种类检索表Tab.14 Key to the species of Setosabatieria
3.4 其他疑似属
3.4.1 长体线虫属(DolichosomatumAllgén, 1951)
ALLGÉN[67]1951年基于2个雌性标本和1个幼体标本建立该属,此后一直未见雄性样本描述。本属主要特征如下:虫体修长,表皮薄且光滑;头部呈圆形;颈部中央刚毛显著,长度约为头长一半;口腔简单,很小,有1颗腹齿;化感器位于口腔后面,为典型的螺旋结构,螺旋数略多于2个,化感器直径7.14 μm,约为头直径的2/5;食道很短,末端中等程度膨大。本属仅包含1个种:DolichosomatumklattiAllgén, 1951,其头部和尾部结构特征见图4a。
3.4.2GrahamiusGerlach & Riemann, 1973
原属名为Grahamia,由ALLGÉN[68]在1959年基于一个雌性标本建立,属名在1973年由GERLACH和RIEMANN[22]变更为Grahamius。主要特征包括:个体小,虫体瘦长;表皮光滑;头呈弧形,外唇刚毛短;口腔呈细管状;化感器远离前端,圆形,在后部有断开;尾直,圆锥形。目前报道仅1个种:GrahamiusodhneriAllgÉn, 1959,其头部和尾部结构特征见图4b。
3.4.3NotosabatieriaAllgén, 1959
Notosabatieria属由ALLGÉN[68]在1959年基于1个雌性标本建立,原始记录描述简单。该属与萨巴线虫属相似,区别在于化感器位置相对更靠后,化感器还有后分支;表皮薄且光滑;2圈头部感觉毛;口腔小,呈漏斗状;食道短,后部有膨大的食道球;尾细长。目前本属仅报道1个种:NotosabatierialeptosomaAllgén, 1959,其头部和尾部结构特征见图4c。
3.4.4UngulilaimusAllgén, 1958
ALLGÉN[69]在1958年基于1个雌性标本和1个幼体形态建立该属,原始描述较简单。主要特征包括:表皮有斑点;头部呈弧形;化感器短;口腔直管状,有几丁质内壁,口腔无背齿,有2个腹齿。该属仅报道1个种:Ungulilaimusfilicaudatus,其头部和尾部结构特征见图4d。

图4 联体线虫科疑似种类头部和尾部示意Fig.4 Anterior end and tail structure of dubiousspecies in the family Comesomatidae(a—Dolichosomatum klatti[66]; b—Grahamius odhneri[67];c—Notosabatieria leptosoma[67]; d—Ungulilaimus filicaudatus[68].)
4 展望
如前所述,近20 a来分子生物学研究方法被引入海洋线虫的分类中,给海洋线虫分类系统带来了很多重要改变,但是当前海洋线虫的分类鉴定仍主要基于形态学特征,对于联体线虫科亦是如此。虽然近些年来许多学者也陆续开展了联体线虫科线虫的测序分析工作[32,70-71],但是相比已有超过230个种类的联体线虫科记录,GenBank数据库中有关联体线虫科的SSU rDNA和LSU rDNA序列各只有20余条,仅涉及10余个种类,大多数联体线虫科仍缺乏分子信息。为此相关研究者呼吁共同利用DNA条形码技术来鉴定线虫物种及物种间亲缘关系,修订已有的分类学体系。

附表1 萨巴线虫属种类形态特征测定Attached tab.1 Morphometric of Sabatieria species
