超富集植物与重金属相互作用机制及应用研究进展
2020-11-03何玉君孙梦荷沈亚婷帅琴罗立强
何玉君, 孙梦荷, 沈亚婷, 帅琴, 罗立强,*
(1.中国地质大学(武汉)材料与化学学院, 湖北 武汉 430074;2.国家地质实验测试中心, 北京100037;3.中国地质调查局广州海洋地质调查局, 广东 广州 510760;4.北矿检测技术有限公司, 北京 102628)
在工业化进程中,大范围的矿山开采和广泛的矿物利用带来了多种生态环境问题,重金属污染的影响尤为严重。重金属将破坏植物细胞活性,引起遗传变异,影响能量代谢,降低光合作用速率,减缓植物生长。重金属也会损伤人体组织器官,危害人类健康。例如,铅会对人体神经、心血管、肝脏、视觉等多个系统造成危害[1];镉会损伤肾脏,引起骨质疏松,或导致癌症等[2]。因此,降低环境中重金属的污染程度与影响,开展无污染绿色生物修复技术研究与应用,维护生态平衡,保障人类健康,是我们当前面临的紧迫任务。
同时,随着人类社会的发展,对矿产资源的需求日益旺盛,但易勘查开采的矿产资源量已显著减少。在2003—2013年十年间,新的金矿发现量下降了45%[3]。在世界范围内,各种天然矿化土地或矿山废料中存在大量无法被工业利用的贵金属。例如,浓缩铜尾矿样品中的Au、Cu含量分别高达5.42g/t、0.408g/t[4];在我国,含金难处理资源已占黄金保有资源的15%,低品位金矿约占已查明金矿资源的1/3左右[5]。如何利用生物技术从矿化土地、低品位矿石及尾矿中开采贵金属,如何利用特异性、指示性植物寻找隐伏矿床,目前受到了国内外的广泛关注。
超富集植物能够从大范围土壤中吸收重金属并将其储存在体内,体内金属浓度比普通植物高出100倍,对重金属具有超富集能力和超耐受性[6-7]。利用超富集植物这一特性,一方面可从重金属污染土壤中去除重金属,实现植物修复[8];从金属含量低的尾矿中提取贵金属,进行植物采矿[9];另一方面,可指示矿区地球化学异常,用于植物找矿[10]。因此,对重金属与超富集植物的相互作用过程进行深入研究,探索超富集植物对重金属的超富集和超耐受机制,提高植物对重金属的富集性、选择性和指示性灵敏度,促进超富集植物的广泛应用,实现绿色矿产勘查与重金属污染修复,具有十分重要的科学意义。
本文首先概述了重金属对植物的作用,并从重金属溶解、超富集植物对重金属的吸收-转运-富集过程等方面,阐述了超富集植物对重金属的富集机理,重点评述了超富集植物对重金属的解毒、耐受机制,同时介绍了其应用前景,以期通过深入认识超富集植物体内金属稳态维持机制,探寻提高植物富集能力的有效途径。
1 营养元素和毒性元素对植物的作用
植物生长时会吸收无机营养元素,同时也会受到重金属毒性作用的影响。Zn、Cu、Mn、Ni等微量元素是生命活动中必需的营养元素,但当其过量存在时,则会对植物产生毒害作用。而As、Pb、Cd和Hg等元素,没有已知生物学功能,即使浓度很低也具有毒性,是毒性最强的重金属[11-12]。
1.1 营养元素对植物的作用
Zn、Cu、Mn、Ni、Se、Co等微量元素是生命活动中必需的营养元素,在植物生长、发育、代谢中发挥了十分重要的作用。营养元素是多种酶和蛋白质的组分,参与植物中多种化合物的形成、氧化还原反应及各类基本代谢活动(表1)。例如,Cu2+和Cu+可与组氨酸、半胱氨酸或蛋氨酸结合,参与植物体内生理活动,并可作为电子载体参与叶绿体、线粒体中的氧化还原和氧化应激反应[13]。

表1 植物中微量元素的作用与机制Table 1 Function and mechanism of micronutrients in plants
营养元素的缺乏会导致植物免疫力下降,引起多种疾病。例如,大部分植物适宜的Cu含量在5~20μg/g之间[14]。当植物中Cu浓度低于5μg/g时,将导致幼叶畸变或萎黄,叶片边缘卷曲;使木质部等组织的细胞壁生成能力减弱,木质化作用减少,引起水分输送不足;Cu缺乏还会破坏顶端分生组织,抑制胚芽的发育和种子的活力,并严重阻碍果实的形成[13]。
微量元素过量会干扰植物细胞内代谢过程,引起细胞损伤。一方面,过量Cu会与S、N和O配体过度结合,抑制酶及功能性金属蛋白活性,损害植物细胞[15]。另一方面,微量元素过量导致的氧化应激反应,是破坏植物细胞活性的主要原因。Cu作为一种氧化还原活性金属,Cu2+可被还原为Cu+(式1)[16]。Cu+参与Fenton反应,形成高毒性活性氧(reactive oxygen species,ROS),迅速引起膜脂质过氧化,导致氧化损伤(式2)[17-18]。当水稻根中Cu浓度达到38.56μg/g时,H2O2和丙二醛浓度分别上升50.11%、43.82%,水稻根生长减少17.7%[19]。
(1)
Cu++H2O2→Cu2++OH·+OH-
(2)
微量元素过量还会导致酶活性降低,光合作用能力退化,植物组织坏死。Zn、Mn等过渡金属会与细胞中的酶和色素结合,抑制生化反应活性。过量Zn会替代光合作用中的重要二价金属离子,如叶绿素(chlorophyll,Chl)中的Mg,相比于[Mg]-Chl的三重激发态,[Zn]-Chl具有较不稳定的单重激发态。[Zn]-Chl的形成使得色素-蛋白复合物变性并引起光合系统退化,造成Zn中毒[20]。Mn作为叶绿体中磷酸化酶的重要组成成分,过量Mn会改变酶的活性,导致光合速率、CO2代谢速率和气孔导度下降,同时使蒸腾速率和植物体内CO2浓度增加,碳代谢和碳利用能力的下降,将使植物生物量降低[21]。过量Zn、Mn对叶绿体光合系统造成破坏,导致光合系统Ⅰ和Ⅱ持续吸收光能,产生的过量且未耗散的能量最终将致使活性氧增加,促进自由基链式反应,造成组织衰老和植物死亡[22]。
Se是目前广为关注的微量元素,既可是营养元素,亦可为毒性元素。微量Se有益于植物生长,可增强白菜(Brassicachinensis)中多种抗氧化酶的活性,抑制过量Cd导致的H2O2和丙二醛(malondialdehyde,MDA)积累,增强植物抗氧化防御能力[23]。然而,由于Se理化性质与S相似,过量Se可与植物体内S的活性位点竞争,形成硒代半胱氨酸,当植物中tRNAcys在转录过程中意外与硒代半胱氨酸而非半胱氨酸结合时,将会产生非特异性硒蛋白,导致中毒症状[24]。而且在过量Se作用下,细胞中维持氧化还原稳态的谷胱甘肽消耗加快,植物中活性氧积累增加,诱导产生氧化应激反应[24]。在2mg/kg硒酸盐作用下,小麦(TriticumaestivumL.)叶片中的谷胱甘肽浓度下降1.02g/kg,H2O2浓度增加1140mmol/kg;此外,Se在叶绿体中还会通过S还原途径进行代谢,使叶绿素含量和光合效率下降,从而导致小麦分蘖期叶片褪绿变白,分蘖数和穗数下降[25]。
此外,关于微量元素的毒性作用与机制目前还存在不同观点,仍需进一步深入研究。例如,对于光合植物而言,是过量Cu抑制细胞活性而产生过量ROS,还是过量Cu导致细胞产生过量ROS而抑制其活性,抑或是二者同时作用,结论尚未可知[26-27]。
1.2 毒性元素对植物的作用
重金属通常会对植物产生毒性作用。这种毒性作用主要是通过破坏植物体内酶和蛋白质结构,替代必需金属元素位点,诱导氧化应激反应等方式,干扰植物体内呼吸作用、光合作用等代谢活动,对植物造成损害。As、Pb、Cd、Hg等重金属没有任何基本代谢功能,可与其他官能团结合,抑制蛋白质功能,破坏植物体内叶绿素合成等多种生理生化过程,被认为是毒性最强的重金属[11-12,31-32]。

(3)
Fe2++H2O2→Fe3++OH·+OH-
(4)
净反应:
(5)
正常情况下,活性氧是植物细胞中叶绿体光合作用、线粒体呼吸作用等有氧活动的副产物,在细胞信号传导过程中充当第二信使,调节多种生理活动[34]。但是,过量活性氧会导致植物细胞内蛋白质、脂类、碳水化合物非特异性氧化;可攻击细胞膜脂质不饱和脂肪酸,引起细胞膜结构和功能改变,引发脂质过氧化链式反应;还可导致酶失活,造成基因毒性和DNA损伤等[35]。
砷酸盐与磷酸盐化学性质相似,可共用相同转运体进行跨膜运输进入植物根细胞[36]。As(Ⅴ)进入植物细胞后,在砷酸还原酶(arsenate reductase,ACR)ACR2作用下,将还原为As(Ⅲ)[37]。与As(Ⅴ)相比,As(Ⅲ)与植物螯合肽、谷胱甘肽中—SH基团亲和性更高。As(Ⅲ)与—SH基团结合后,将间接灭活抗氧化酶分子,导致活性氧增加[38]。还原过程之后是无机砷的甲基化,甲基化产生的单甲基胂酸、二甲基胂酸、三甲基氧化砷、砷胆碱等甲基化砷,易与O2发生反应,产生甲基砷自由基以及活性氧[39]。甲基化砷还可从线粒体铁蛋白中传递氧化还原活性铁,这些活性铁通过Haber-Weiss反应释放OH·和有害氧[33,40]。因此,我们认为砷诱导的植物氧化应激反应可能机制是由砷的还原与甲基化过程及Haber-Weiss反应组成,如图1所示。过量活性氧的产生将对植物造成不可逆破坏,在10μmol/L As(Ⅴ)的作用下,绿豆(PhaseolusaureusRoxb.)中MDA和H2O2浓度分别增加了2.4倍、2倍,导致细胞膜脂质过氧化程度增加,细胞膜受损[41]。

图1 砷诱导氧化应激机制Fig.1 Mechanism of arsenic induced oxidative stress

2 重金属超富集植物特性与富集机制
植物对重金属元素的超富集是一种复杂而罕见的自然现象,是生物适应与进化的结果。这种环境适应性进化不仅使得植物对重金属具有了选择性,还使其产生了对重金属的特异性超富集能力。超富集植物根系分泌物与微生物对重金属的溶解和地上部富集,是其实现超富集过程的关键性作用机制。
2.1 超富集植物特性与选择性进化机制
自然界富含重金属的土壤中,会生长着一些特定植物,其体内重金属浓度甚至可比普通植物高出100倍,而植物并不显现出明显中毒特征。通常此类植物被称为超富集植物[6-7]。人们也会针对元素特点,依据植物地上部的重金属浓度来定义超富集植物。例如,当植物体内Cd、As浓度>100mg/kg,Co、Cu、Ni、Pb浓度>1000mg/kg,Mn、Zn浓度>10000mg/kg,或Au浓度>1mg/kg时,则称这些植物为相应重金属的超富集植物[46-47]。
植物自身特性与来自周围超高浓度重金属环境压力,会使得超富集植物逐步适应生存环境并成为当地的优势物种。目前主要有三种观点来解释超富集植物的适应性进化过程与选择性优势:① 金属耐受/去除假说(metal tolerance/disposal hypothesis)。认为重金属首先在超富集植物地上部积累,并通过落叶去除重金属,降低植物体内重金属浓度,来实现耐受—去除—适应性生存这一进化过程[48];② 干扰假说(interference hypothesis)。认为超富集植物会通过落叶增加其周围表层土壤重金属浓度来抑制附近植物的生长,促进其有效争夺植物生长所需的空间和光照;③ 防御假说(elemental defense hypothesis)。也是目前最受支持的假说,认为植物中重金属对食草动物和病原体侵害起到了防御作用[29]。例如,摄入过量Se将导致中毒,因此,硒超富集植物沙漠王羽(Stanleyapinnata)对Se的超富集使其免受了土拨鼠等草食动物的侵害,促进了该植物Se超富集特性的进化[49]。

Mx+代表金属离子。
2.2 重金属超富集植物富集过程
重金属超富集植物的富集过程主要包含四个步骤。①土壤中重金属元素的活化、溶解;②植物根对重金属元素的吸收;③重金属元素向植物地上部分转运;④解毒、隔离。如图2所示。
2.3 重金属超富集植物富集机制
2.3.1根际土重金属活化与溶解
超富集植物根系分泌物将使根际形成酸性环境,促进土壤中的重金属溶解[50]。重金属污染条件下,超富集型东南景天(Sedumalfredii)根系可分泌更多可溶性有机质,可达38.2%,远高于非富集型(6.3%),根际有机酸浓度变化趋势与之一致,这使其根际土pH值比块土低0.27,而非富集型东南景天根际土与块土pH无显著差异[51]。酸性条件下,重金属生物有效性更高。当 pH<7.7时,游离Zn2+在溶液中占主导地位;当pH>7.7时,Zn的主要形态Zn(OH)+易于沉淀,不具生物有效性[52]。弱酸条件下,CuO纳米颗粒与弱酸性土壤中的H+相互作用,溶解为Cu2+或Cu(OH)+释放到土壤中(式6、式7),为水稻所吸收[53]。
CuO(s)+2H+(aq) ↔ Cu2+(aq)+2H2O(l)
(6)
CuO(s)+H+(aq) ↔ Cu(OH)+(aq)
(7)
超富集植物特异性根系分泌物可通过与重金属络合形成可溶性复合物,促进重金属在植物体内的富集。在As的作用下,As超富集植物蜈蚣草(Pterisvittata)、井栏边草(Pterismultifida)等蕨类植物的主要根系分泌物是植酸和草酸(>93%),而在西红柿根系分泌物未检出植酸,表明植酸是蕨类植物特异性根系分泌物[54]。As胁迫会使蕨类植物中植酸分泌量增大,蜈蚣草植酸分泌量比非富集植物剑叶凤尾蕨(Pterisensiformis)高262%,植酸含有6个磷酸根基团和12个可解离质子,与Fe有很强的配位能力,从而促进了根际FeAsO4的溶解,可有效增强蜈蚣草对Fe和As的吸收[54]。
超富集植物根际微生物与根系分泌物共同作用,可增强土壤中重金属生物有效性。一方面,植物根系分泌的有机化合物、氨基酸、多糖可为微生物提供所需碳源、氮源,为微生物活动创造富营养环境,使根际微生物的活性提高10~100倍,改善重金属所导致的土壤细菌群落多样性减少现象[55-56]。另一方面,微生物代谢产生粘胶或小分子有机酸等分泌物,也可促进重金属溶解[57-58]。Mn超富集植物伏毛蓼(Polygonumpubescens)中分离出的内生菌JN6对Pb、Cd、Zn都有很强的耐受性,可使培养基pH值降低约1~2,有效溶解CdCO3、PbCO3和Zn3(PO4)2;同时,JN6分泌的吲哚乙酸、铁载体、ACC脱氨酶以及可溶性磷酸盐,还可促进植物营养物质吸收,有利于超富集植物的生长[59]。
2.3.2吸收与转运
(1)共质体与异质体吸收
超富集植物吸收重金属主要通过两个途径:共质体途径和质外体(异质体)途径。共质体途径是指利用Ca、Fe等必需营养元素离子通道、转运蛋白等方式进行跨膜运输,而质外体途径则是通过细胞壁、胞外间隙等横向传递至木质部的过程。
在共质体途径中,重金属通常借用必需营养元素的传输系统穿过根部细胞膜进入细胞。离子通道选择性较低,Cd2+、Mg2+和Zn2+等通过钙离子通道穿过苋菜(AmaranthusmangostanusL.)根细胞而被吸收[60]。此外,Zn、Fe转运蛋白(ZRT、IRT)对Cd有很强的亲和力,参与Cd的转运。当油菜(Brassicanapus)中IRT基因表达水平较高时,其根部Cd浓度更高[61]。
质外体途径无需穿过质膜,允许有毒金属元素通过。Cd2+可通过东南景天根尖和侧根间隙进入植物根部,富集型东南景天木质部中高达37%的Cd2+来自质外体途径[62]。但细胞壁间隙孔径大小具有筛分性质,使细胞壁成为植物根部排阻屏障[63]。例如,3.5nm金纳米颗粒(AuNPs)可通过根部质外体途径进入烟草(Nicotianaxanthi)维管柱及细胞质,导致叶片坏死;但18nm AuNPs受到细胞壁排阻,仅在根部外表面团聚,无法进入烟草根内[64]。
(2)木质部转运
重金属进入根部后,将通过木质部及韧皮部由根部向地上部分转运。木质部转运是超富集植物重金属转运的主要途径,小分子有机酸、蛋白酶等是其主要影响因素。例如,在超富集植物多穗稗(Echinochloapolystachya)中,Cd主要通过木质部从根部向叶片转运[65]。
木质部汁液中小分子有机酸和氨基酸有助于重金属向上转运,是重金属超富集原因之一。超富集植物龙葵(SolanumnigrumL.)根部分泌的小分子有机酸总量显著高于西红柿(SolanumlycopersicumL.),使Cd在龙葵中的转运率(1.33~1.89)高于西红柿(0.45~0.84)[66]。富集型东南景天木质部汁液中柠檬酸浓度比非富集型高6倍,Zn浓度高7~9倍;富集型东南景天木质部汁液中有机酸、柠檬酸约占36.7%~42.3%,55.9%以上的Zn以水溶态转运[67]。组氨酸会参与Zn的吸收,同时还可显著提高褐蓝菜(Noccaeacaerulescens)木质部中Zn的负载量,有助于其径向运输,是褐蓝菜中Zn超富集机制之一[68]。
重金属ATP酶(HMA)和锌铁通透酶(ZIP)等是植物体内Cd、Zn等重金属的主要转运蛋白。Cd和Zn从水稻根到地上部的转运主要依赖于重金属ATP酶OsHMA2[69];Cd胁迫下,富集型东南景天中HMA4基因表达水平比非富集型东南景天中高2.7倍[70]。此外,通过克隆技术制备特殊转运体,如烟草(Nicotianatabacum)中的NtZIP5A/B缺锌诱导转运体,可控制Zn和Cd在植物中的纵向分布以及金属转运率,实现Cd胁迫下Zn的向上转运[71]。然而,酶蛋白也会限制金属向上转运。在水稻和拟南芥中,根系表达的HMA3将重金属螯合在根部细胞中,以此限制Cd向植物地上部的转运[72-73]。不同酶蛋白作用有所不同,通过调控转运蛋白基因表达提高超富集植物重金属转运率,值得深入探索。
除木质部运输以外,韧皮部也参与植物中重金属的转运。韧皮部中重金属运输过程与营养物质转运有关,重金属既可通过韧皮部向上转运至种子、新叶中,也可向下转运到根细胞中[74]。Ni、Zn同位素示踪研究表明,褐蓝菜韧皮部汁液中的苹果酸会与Ni、Zn螯合,老叶中89%的61Ni会向上转运至幼叶中,11%则向下转运至根中[75]。超富集型东南景天老叶中68Zn再活化能力比非富集型高约7倍,并能将其优先分配到新叶韧皮部周围叶肉细胞中,以满足新叶对高浓度矿物质元素的需求[67]。鉴于超富集植物中的重金属元素主要富集在叶片中,对叶中重金属元素再活化机制进行深入研究,将有助于提高超富集植物重金属的转运率。
2.3.3地上部储存与富集
对于普通植物而言,重金属主要在植物根部积累。例如,Au以纳米颗粒形式沉积在大麦、西红柿等植物根中,很少向上转运,以免对植物地上部分生理过程造成破坏[76-77]。与普通植物相比,超富集植物中的重金属木质部转运和地上部分解毒能力更强,且主要富集在地上部分。
超富集植物根部栓质形成较弱,是重金属在超富集植物地上部富集的主要原因之一。植物根部木栓质(suberin lamellae)是一种质外体屏障,可将中柱与外围细胞分开,阻碍金属离子通过质外体途径向地上部转运[62]。与非富集型东南景天相比,富集型东南景天根部栓质化基因表达下调,根尖和侧根处栓质发育不良,有利于Cd2+质外体运输;而非富集型东南景天侧根中栓质发育充分,阻碍Cd2+质外体运输[62]。在Cd胁迫下,富集型东南景天中脱落酸合成基因(SaABA2,SaNCED)不会上调,避免了脱落酸所引起的栓质形成和沉积,使Cd2+可以顺利向上转运[78]。
超富集植物根部细胞壁重金属结合能力也是影响重金属超富集的重要因素。Cd胁迫下,超富集型东南景天根部细胞壁多糖和果胶甲基酯酶浓度和活性均低于非富集型,且富集型根细胞壁中果胶甲基化程度高于非富集型,使超富集型东南景天根部细胞壁中Cd更易解吸并向上转运[79]。
超富集植物主要通过地上部叶片液泡实现重金属的富集和隔离。在非富集植物中,重金属元素主要储存在根表皮细胞、外皮和中柱[80-81]。而在超富集植物中,重金属主要储存在叶片的液泡中,并以此避免对细胞重要生理过程造成破坏[65,82]。富集型苋菜会分泌大量植物螯合肽和小分子有机酸,与Cd形成特定化合物并转运到液泡中隔离,降低Cd毒性[80]。在超富集植物九重葛(BougainvilleaspectabilisWilld.)液泡中,Cd以柠檬酸盐或Cd(H2PO4)2的形式存在,实现对Cd的隔离、富集和解毒[83]。超富集植物中重金属主要富集、隔离在叶片液泡中,对液泡中重金属的分子结构与解毒机制进行探索,对提高超富集植物重金属富集能力具有重要意义。
3 重金属超富集植物解毒与耐受机制
超富集植物之所以具有特异性富集能力,其主要原因就是它们具有独特的解毒和耐受机制。重金属既可与植物体内小分子有机酸、植物细胞壁、植物螯合肽等结合,也可通过液泡隔离的方式,降低细胞质中游离金属离子浓度来进行解毒。同时,超富集植物还可激活抗氧化酶抵抗重金属毒性。
3.1 小分子有机酸络合作用
小分子有机酸不仅可以通过络合作用调节重金属生物利用度,也可增强植物对重金属耐受能力。一方面,植物释放出的草酸和苹果酸等小分子有机酸会随Pb、Cd、Cu、Zn等重金属浓度的增加而增加,在与重金属形成稳定络合物的同时,还能改善土壤质量,使其能在高污染水平下顺利生长[84-85]。柠檬酸有很强的配位能力,与Cd2+结合后使之不带电荷,降低游离Cd2+浓度,有效减少植物吸收和由此造成的毒性伤害[83,86]。少量吸附在土壤固体表面的甘氨酸可与Pb、Cd络合,增强对重金属的吸附,从而降低其对植物的毒性[87]。另一方面,植物体内有机酸也可增强植物抗氧化能力。超富集植物秋茄树(Kandeliaobovata)会分泌酚酸,其羧基、羟基可与Cd、Zn离子结合形成络合物,酚羟基能有效清除羟基自由基,以减轻金属应力引起的氧化损伤[88]。
3.2 植物细胞壁结合
植物细胞壁可与重金属结合,减少细胞质中重金属浓度,增强植物抵御重金属毒性能力。在超富集植物伴矿景天(Sedumplumbizincicola)中,Zn主要储存在根皮层,毒性元素Cd则主要存在于质外体,尤其是在细胞壁中[82]。Pb、Cd主要以细胞壁结合态的形式储存在荻(Triarrhenasacchariflora)中,以免破坏细胞质中重要代谢活动[89]。此外,Cd胁迫下,富集型东南景天中纤维素合成酶、糖苷水解酶,以及与木质素形成相关酶的基因表达都会有所上调,使Cd主要结合在植物细胞壁中,减少游离Cd浓度[70]。
重金属在细胞壁中以稳定性强、不溶性高的复合物形式存在,降低其毒性,是超富集植物实现重金属耐受与解毒的分子机制。印度芥菜根细胞壁中,Zn主要与羧基结合,以Zn-聚半乳糖醛酸(C12H14O12Zn·4H2O)形式与细胞壁结合;形成难溶Zn-植酸、Zn-碳酸也是印度芥菜调节细胞毒性的主要形式[90]。在荻根中,Pb主要以不溶性磷酸结合态、草酸结合态存在;在玉米根中,Pb以难溶磷氯酸铅、碳酸铅的形式存在,降低了游离态金属离子浓度,增强了毒性耐受能力[89,91]。
3.3 植物螯合肽螯合作用
天然配体植物螯合肽(Phytochelatins,PCs)、金属硫蛋白(Metallothioneins,MTs)、谷胱甘肽(Glutathione,GSH)等可与重金属形成硫醇键,在植物对重金属的解毒、隔离中发挥了重要作用[92]。
在植物螯合肽合成酶的作用下,由GSH可产生PCs。与游离金属离子相比,PCs-金属复合物稳定性高、毒性小;同时,在液泡膜转运蛋白作用下,PCs-金属复合物将转移至液泡中,两者共同作用降低了细胞质中游离重金属离子的浓度[93]。随Ni浓度升高,紫花苜蓿(Medicagosativa)中参与PCs合成的相关基因的表达将逐渐上调,随后PCs与Ni结合形成PCs-Ni复合物,实现对重金属的解毒[94]。
MTs是低分子量蛋白质且富含半胱氨酸,其中的硫分子对金属离子具有很强的亲和力,可通过硫分子或半胱氨酸中的硫醇基团与不同的金属离子结合实现解毒[95]。GSH与MTs结构相似,在绿藻(Ulvacompressa)中,两者以协调、互补的方式参与绿藻中铜积累和解毒[96]。此外,施加硫肥可显著提高水稻根中GSH和PCs含量,以及植物螯合肽合成酶和液泡膜重金属ATP酶的基因表达水平,使根部螯合与液泡隔离的Cd量增加,减少水稻籽粒中Cd的吸收和转运[97]。如何通过基因调控,进一步提高植物对Cd等毒性元素的隔离能力,是下一步的发展方向。
3.4 植物液泡隔离
重金属隔离在植物液泡中是植物维持体内重金属稳态、减少其毒性作用的重要方式。液泡中的含硫肽和有机酸,可与重金属离子螯合,并将其限制在有限位点,从而降低游离金属离子的活性,避免其对代谢活跃细胞器造成毒害[98]。在超富集植物九重葛、多穗稗、伴矿景天、苋菜中,Cd主要储存在叶片的液泡中,并以柠檬酸盐的形式存在,减少Cd对植物的毒性作用,也防止其对光合作用产生不利的影响[65,80,82-83]。

PCs—植物螯合肽:MTs—金属硫蛋白;Acids—小分子有机酸,Mx+—金属离子。
在液泡膜HMA、ABC等多种重金属转运蛋白作用下,与植物螯合肽结合的重金属会被泵入液泡中,实现对毒性元素的隔离,如图3所示。转运蛋白HMA3位于液泡膜,介导重金属离子的液泡隔离[99]。SpHMA3是伴矿景天中Cd超耐受相关基因。Cd胁迫下,野生型伴矿景天并无明显毒性症状,但经RNA干扰后,会使SpHMA3基因表达减少约85%,从而导致SpHMA3-RNAi型伴矿景天幼茎生长迟缓,幼叶萎黄[100]。C型ABC转运蛋白AtABCC1和AtABCC2是介导拟南芥As耐受性的主要植物螯合肽转运蛋白[101]。在野生型拟南芥中,Cd主要储存于液泡中,但在去除AtABCC1和AtABCC2两个转运蛋白的突变体拟南芥中,Cd则主要储存于细胞质中,导致耐受性降低;而 AtABCC1的过表达,则可增强拟南芥对Cd(Ⅱ)耐受性和积累[102]。
金属转运蛋白具有物种特异性和器官特异性,对各液泡转运蛋白分子结构和功能特性进行研究,利用转基因技术调控转运蛋白表达,设计出适合植物修复的特异性植物,对进一步提升植物重金属超耐受性、超富集性具有重要意义。
3.5 抗氧化酶抗氧化作用
重金属胁迫下,植物会激活多种抗氧化酶,提高植物体内抗氧化酶水平,减轻活性氧过量所引起的细胞损坏。这些酶包括过氧化氢酶(catalase,CAT)、过氧化物酶、超氧化物歧化酶和抗坏血酸过氧化物酶,谷胱甘肽还原酶(glutathione reductase,GR)等[12]。不同重金属胁迫下,超富集植物抗氧化酶表达和活性差异很大,如表2所示。

表2 在不同培养条件下生长的不同植物中,重金属激活抗氧化酶差异Table 2 Heavy metal-induced activation of antioxidant enzymes in different plant species grown in different condition
Pb胁迫下,金丝草(Pogonatherumcrinitum)叶中SOD、POD酶活性提高,可降低重金属所引起的细胞膜损坏[85]。Ni作用下,紫花苜蓿中Prx1C基因表达上调,POD和谷胱甘肽硫转移酶(glutathione-S-transferase,GST)活性大幅提升,减轻了Ni对植物细胞膜造成的破坏[94]。此外,低浓度Pb还会刺激长根菇(Oudemansiellaradicata)产生硫醇,参与非酶类抗氧化剂(非蛋白巯基、谷胱甘肽)的合成,并在2d/4d后激活抗氧化酶[103]。
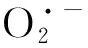
细胞内H2O2含量主要受POD和CAT调节。CAT通常存在于线粒体和过氧化物酶体中,通过CAT,H2O2将分解为H2O和O2;而POD则通过酚类化合物/抗氧化剂的氧化作用降解H2O2[105]。然而,过量重金属诱导的氧化应激产生的H2O2可能抑制CAT、POD活性,此时,则需结合APX等其他抗氧化酶共同作用清除SOD产生的H2O2[106]。APX对H2O2亲和力强于其他酶,可将H2O2分解为H2O和O2,在清除活性氧中起到了十分重要的作用[107]。
除抗氧化酶抗氧化作用外,某些重金属对植物的氧化胁迫会存在协同拮抗作用。亚硒酸盐会增强白菜中APX、GR、SOD、CAT、POD的活性,抑制H2O2和MDA的积累,增强其抗氧化防御能力,减少白菜中Cd富集以及Cd引起的氧化应激水平[23]。Cd胁迫下,低浓度Zn有助于恢复并增强西红柿中抗氧化酶的活性,显著降低植物氧化应激水平;而高浓度Zn、Cd共同作用时,氧化应激水平要高于过量Cd或Zn单独作用时的水平[108]。
在重金属胁迫下,超富集植物会激活多种防御策略抵抗重金属元素生物毒性。但有些观察还存在相悖的实验报道和理论解释,例如,小分子有机酸与重金属络合作用一方面可增强重金属溶解性[54],另一方面可提高重金属稳定性,降低其生物毒性[87]。而且,超富集植物超富集和超耐受性与植物螯合肽、抗氧化酶等多种蛋白质相关,如何准确理解超富集植物的隔离与耐受分子机制,是未来需要深入探索的研究方向。
4 超富集植物的作用与价值
由于对重金属具有超富集能力及特定的选择性和指示性,超富集植物已在植物修复、植物采矿和植物找矿中得到广泛应用。截至2017年7月已有721种(Ni:523种,Cu:53种,Co:42种,Mn:42种,Zn:20种等)植物被判定为超富集植物[115]。表3列出了部分超富集植物及其富集能力。

表3 部分重金属超富集植物及其富集能力和应用Table 3 Accumulation ability of several hyperaccumulators and their application
利用超富集植物对重金属的超富集能力,不仅可以去除土壤中重金属,实现土壤重金属污染绿色可持续修复;还可通过植物提取,回收利用土壤中贵金属,获取经济效益;此外,利用某些植物对特定金属元素的指示作用,还可应用于生物地球化学找矿[3]。
4.1 植物修复
植物修复是指利用超富集植物从土壤中提取重金属并将其转移到植物组织中,通过收获和处理植物,去除土壤中重金属污染的过程。农田中超富集植物藿香蓟(AgeratumconyzoidesL.)Cd富集系数可达6.93,种植三茬藿香蓟Cd去除率约为13.2%~15.6%[121]。菊芋(Helianthustuberosus)地上部分中重金属浓度分别为:Mn(1.14%)、Zn(6.1g/kg)、Ni(2.6g/kg)、Cd(0.56g/kg),且Ni、Zn、Mn从根到地上部分的转运率在11~15之间,可实现多种重金属的超富集以及土壤修复[122]。金合欢树(Acacia)能有效吸收尾矿土壤中的多种重金属,并在尾矿上快速建立植被,避免污染物向周围环境扩散,是修复多元素污染的重要候选植物[123]。
在植物修复中,植物从土壤中去除污染物的量与其组织浓度及生物量大小成正比[124]。通过传统育种、基因工程等途径,可有效克服自然界超富集植物生长缓慢、产量低的缺点,提高和改善超富集植物修复土壤重金属污染能力[98]。传统育种主要是利用物种内部现有的遗传多样性来改进所需性状。对13种藜属植物(Chenopodiumspp.)进行3次连续随机插条实验后,株高、叶片大小、蛋白质和叶片产量都会增加,可有效提高植物生物量[125]。此外,利用基因工程可为植物修复创建理想植物,有效控制植物耐受、积累污染物的能力。利用汞离子还原酶基因merA或merB,通过核或叶绿体基因组修饰植物,在细胞质、内质网中表达有机汞裂解酶或汞离子还原酶,所获得的转基因植物在被有机(约400μmol/L PMA)或无机(约500μmol/L氯化汞溶液)汞污染的土壤中生长极佳,其根中的Hg累积可达约2000μg/g[126]。
植物修复无需添加可引起二次污染的化学试剂,具备绿色可持续的特点,在农业土壤和尾矿污染修复中具有良好应用前景,是当前和今后一段时期污染修复的发展方向。
4.2 植物采矿
植物采矿与传统采矿相比,对环境影响较小,还可改善土壤质量,为从金属含量低尾矿和矿化土地中开采贵金属提供了一种新途径。目前对于植物采Ni的研究较多。例如,在富Ni蛇纹石土壤中种植Odontarrhenachalcidica、小野芥菜,Ni产量分别可达55kg/公顷、36kg/公顷[116]。
与植物采Ni不同,由于已知Au、Ag等惰性贵金属超富集植物较少,目前Au、Ag植物采矿通常在化学诱导剂辅助下进行。硫氰酸铵或硫代硫酸铵化学诱导剂可显著提高欧洲油菜的根、茎、叶中Au、Ag、Cu浓度,使植物器官中Au、Ag、Cu浓度高于超富集标准,有效提高欧洲油菜植物采矿能力[127]。
植物采矿成本较低,可获取一定经济效益,可行性高。利用超富集植物Berkheyacoddii从富Ni土壤中获取Ni,预计可获利约11500AU$公顷;印度芥菜植物采Au,预计可获利约26000AU$/公顷[128]。
4.3 植物找矿
植物找矿是寻找隐伏矿床的潜在有效手段。环境中的地球化学异常会使植物生理和生态特征发生变化,逐渐成为当地优势物种。例如,海州香薷(ElscholtziahaichowensisSun)是铜矿区优势植物,对Cu具有较强富集和耐受能力,可作为铜矿指示植物[129]。Cu耐受型海州香薷由于长期受Cu胁迫,根中酸性转化酶基因启动子上的甲基化状态可能已经发生变异并可稳定遗传给后代。因此,在Cu胁迫下耐受型海州香薷根中酸性转化酶活性及其基因转录水平明显高于非耐受型,以抵抗Cu生物毒性[130]。
除指示作用外,通过植物中金属含量,可获得基质中金属元素浓度的宝贵信息,以此识别矿物勘探潜在区域。Masjed-Daghi金-铜矿区的Stachysinflata对Cu、Hg生物吸收系数>1,达到超富集植物的标准;Au、Ag生物吸收系数分别为0.33、0.71,与土壤中金属含量有一定相关性,可作为指示植物用于植物找矿[131]。
桉树(Eucalyptus)是一种受到关注的特殊树种。桉树根系长达40m,可穿过风化层抵达富金区域,主动吸收土壤中的金,且金在桉树叶中以粒子形式存在;Freddo矿区上方大型桉树(高度>10m)叶、枝等部位Au浓度异常高,该结果为植物找金展示了良好前景[3]。此外,在澳大利亚Boddington金矿区,除桉树外,山龙眼(Banksia)、大泽米(Macrozamia)等其他植物,也可指示Au元素地球化学异常,有助于发现深埋地层矿藏[10]。
目前,利用特异性植物寻找专属隐伏矿床的研究,仍是国际上正待发展和探索的前沿领域。生物找矿的指示性、特异性和指示灵敏度,是植物找矿研究中面临的主要挑战。
5 结论与展望
超富集植物对重金属具有超富集、超耐受能力,是降低环境重金属污染、保障人类健康,实现绿色矿产勘查、利用的有效途径,在植物修复、植物采矿和植物找矿中已获得了广泛应用。国内外研究表明,土壤重金属在超富集植物根系分泌物与根际微生物作用下溶解,经共质体和质外体途径吸收并由木质部转运至地上部后,会被植物体内小分子有机酸、细胞壁和植物螯合肽结合,或隔离在液泡中,以此降低细胞质中游离金属离子浓度,增强植物耐受性。受重金属胁迫的超富集植物,还会激活多种特异性抗氧化酶抵御氧化应激反应,从而实现对重金属的解毒、耐受与富集。此外,本文分析认为,砷诱导的植物氧化应激反应机制可能由砷的还原与甲基化过程及Haber-Weiss反应三部分构成。
从当前的研究进展来看,超富集植物目前所具有的选择性、指示性和耐受性还不理想,富集能力不足;其超富集、超耐受所涉及的小分子有机酸、蛋白质、酶等的结构与作用过程复杂,作用过程与反应机制还需深入探索。深刻认识有机酸改变重金属吸收、转运能力的作用过程与机理;从基因组学和代谢组学的角度,探索植物富集及耐受相关基因,揭示影响超富集植物特性的关键因素和作用规律,利用基因编辑等技术设计并开发选择性和指示性高、富集能力强的理想超富集植物,将可推动超富集植物在生物修复和资源勘察利用中的应用与发展,为保护人类健康、保障资源供给提供有力支持。
