基于巢式分组设计的板栗刺苞与坚果形态多样性分析
2020-11-03
(北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083)
板栗Castanets mollisimaBlume 为壳斗科Fagaceae 栗属Castanea植物,在我国有悠久的栽培历史[1],我国驯化栽培最早的果树之一,分布范围广泛,种质资源丰富[2]。当前,我国发现并确定的板栗品种有300 多个,建立4 处栗属种质资源库,收集并保存6 个种347 份种质材料[3]。其在世界食用栗类群中占有重要地位,是世界各国进行食用栗品种改良的重要基因来源[4]。
根据产区的气候条件、土壤质地、栽培管理、人工定向选择以及品种性状特性等因素,分为6个地方品种群,即华北品种群、西北品种群、东北品种群、东南品种群、西南品种群和长江流域品种群[5]。中国栗的居群具有很高的遗传杂合度,居群的变异幅度大,居群结构复杂,遗传基础极为丰富,是世界现有的栗属植物遗传资源的宝贵基因库[6]。
生物多样性是遗传多样性、物种多样性和生态系统多样性的综合体现,遗传多样性是生物多样性的重要组成部分。遗传多样性最直接的表现是表型多样性,表现型和基因型之间存在着基因表达、个体发育、环境调控等复杂的中间环节,根据表现型差异来反映基因型差异具有重要的意义[4,7]。
目前,板栗品种划分方法主要依据其形态学特性、生物学性状和经济性状。其中,以花序、总苞和果实的形态差别为最主要的划分依据[8]。同时,果实性状的表型变异也是研究植物种群的一个重要组成部分[9]。目前,许多学者已经对不同板栗种质资源的叶片、枝条、坚果、刺苞等进行了研究,但是关于迁西地区不同产地来源的板栗刺苞及坚果的表型形态性状差异与多样性研究较少。在该研究中,以定植于河北省迁西县东寨板栗种质资源圃的39 份板栗种质资源为试验材料,对其刺苞、坚果表型性状进行多样性分析,以揭示不同板栗种质资源在迁西地区表型多样性、变异规律及变异水平,对于探讨不同板栗种质资源对迁西地区环境的适应能力、不同种质板栗坚果的综合评价及对迁西地区板栗的引种、选育新品种等工作具有重要意义。
1 材料与方法
1.1 试验材料
以源自6 个板栗主产省份(群体)的39 份板栗种质资源为试验材料,定植于河北省迁西县东寨板栗种质资源圃。该地区年平均气温10.1 ℃,多年平均降水量为804.20 mm。
在该试验地,南北方向每列种植34 株板栗,株行距2 m×3 m,南北两侧种植两行保护行,每列包含有3 个不同板栗种质资源,每个板栗种质资源定植10 株,均为2013年以接穗的形式嫁接到3年生实生母树砧木上。试验地气候、土壤等条件、水肥用量以及其它管理措施相一致。种质编号、名称及来源如表1所示。
1.2 试验方法
于2018年9—10月(果实成熟期)采集果实。在每个种质的10 棵树中选择长势一致并能代表该种质的5 株试验树,挂牌标记并记录,每株试验树选择树冠中部外围东、南、西、北四个方向的结果枝,随机采取2 个刺苞,每株试验树采集8个刺苞,每种质共计采集40 个刺苞,以供刺苞与坚果表型性状调查。
1.2.1 刺苞表型性状调查
刺苞表型性状调查参考平文倩[10]的板栗新品种燕秋植物学特性的调查方法、陈佳佳等[11]对板栗果实特性的调查以及刘庆忠等[12]编著的《板栗种质资源描述规范和数据标准》。
刺苞横、纵、高三径:在测量时,因刺束的关系往往不易测量。该试验在测量时,选取生长正常、苞内含有3 粒坚果、成熟接近开裂且暂未开裂的刺苞,用游标卡尺在刺苞沟谷处测得。每个板栗种质选取样本10 个求其平均值,数据单位为cm,精确到0.01 cm。
刺苞肉厚度:在坚果成熟时期,每个板栗种质从成龄树上随机选择30 个生长正常的开裂刺苞,用游标卡尺测量刺苞中部的苞肉厚度,并求其平均值,单位为mm,精确到0.01 mm。
刺束粗细:在坚果成熟期,从试验板栗树上随机选择生长正常的初裂刺苞10 个,用游标卡尺测量刺束直径,单位为mm,精确到0.01 mm。
刺束长度:在坚果成熟期,从每个板栗种质随机选取生长正常的开裂刺苞10 个,用游标卡尺测量20 个刺束长度(刺束长度测量时包括刺及刺座),并求其平均值。单位为cm,精确到0.01 cm。
1.2.2 坚果表型性状调查
边果性状调查:取栗苞中板栗边果20 粒,用游标卡尺测量其长度、宽度,单位为mm,精确到0.01 mm。果形指数计算公式为[13]:

表1 板栗采种群体与种质来源Table 1 Origin sites and seed groups of Castanea mollissima
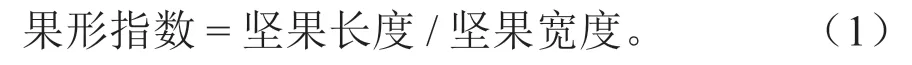
坚果种皮厚度:取板栗坚果20 粒,剥掉外层种皮,用游标卡尺测量其厚度,取均值,单位为mm,精确到0.01mm。
1.3 数据分析
利用Excel 2010 进行数据记录与制作表格,用SPSS 20.0 软件进行差异性分析,用R 语言软件对数据进行双因素巢式分组设计方差分析和聚类分析。
采用双因素巢式设计方差分析,线性模型为:Yijk=μ+Si+T(i)j+ε(ij)k。式中:Yijk为第i 个群体第j 个家系第k 个观测值,μ 为总均值,Si为群体效应(固定),T(i)j为群体内家系效应(随机),ε(ij)k为试验误差[14]。

式中:VST为表型分化系数,表示群体间变异占遗传总变异的百分比[15-16],式中:δ2t/s为群体间方差分量,δ2s为群体内方差分量[14-15]。
性状变异系数(CV)表示性状离散程度[14,16],式中:为性状平均值,s 为标准差。

2 结果与分析
2.1 板栗刺苞、坚果表型性状群体间多样性分析
通过对不同来源的39 份种质板栗刺苞、坚果表型性状进行一般性分析,由表2~5 可发现,板栗刺苞、坚果相关的10 个表型性状在种质间具有极显著差异;说明不同来源的板栗刺苞及坚果的表型形态在种质间存在广泛的变异,丰富度大。通过对6 个群体板栗的刺苞、坚果表型性状进行分析得出,北京、江苏、湖北板栗群体较河北、河南、山东板栗群体种皮厚;湖北、山东板栗群体刺苞横、纵、高三径较大,苞肉较薄,其坚果形态较大;河南板栗群体与北京、河北板栗群体相比较,其刺苞三径值无显著差异,苞肉厚度存在显著差异,在坚果表型形态上表现出坚果较小。
采用双因素巢式分组试验设计方差分析,探究不同来源板栗的刺苞、坚果性状在群体间和群体内两个层次上的差异显著性,见表6。结果表明:刺束长度这一指标在群体内存在显著性差异(F=7.72**),而在群体间差异性不显著(F=3.17),而其余9 个性状指标在群体内和群体间差异均达到了显著水平,说明板栗坚果这9 个农艺性状在群体内和群体间多样性丰富,存在着广泛变异,具有较强的选择性。

表2 39 份板栗种质刺苞表型性状数据一般性分析†Table 2 General analysis of phenotypic traits data of 39 chestnut thorns bract
2.2 板栗刺苞、坚果表型性状群体内多样性分析
由表7可知,板栗刺苞、坚果10 个表型形态性状平均变异系数在5.37%~21.02%之间,平均变异系数差异较大,其中,种皮厚度、刺束长度与粗度、苞肉厚度的平均变异系数较大,均大于10.82%,表明其性状遗传稳定性较低,具有较大的遗传潜力,果形指数平均变异系数最小(5.37%),表明其具有较高的遗传稳定性;板栗不同部位其平均变异系数也存在较大差异,坚果表型性状平均变异系数为9.27%,较板栗刺苞表型性状平均变异系数(11.85%)小,表明坚果表型性状具有较高的遗传稳定性;不同群体板栗刺苞坚果表型形态性状平均变异系数也存在较大差异,其中北京、河北、湖北、山东板栗群体平均变异系数较大,相较于其他2 个板栗群体,其板栗表型形态性状多态性较为丰富,性状遗传稳定性较低,在很大的程度上能够选育出新种质,具有较大的遗传潜力。
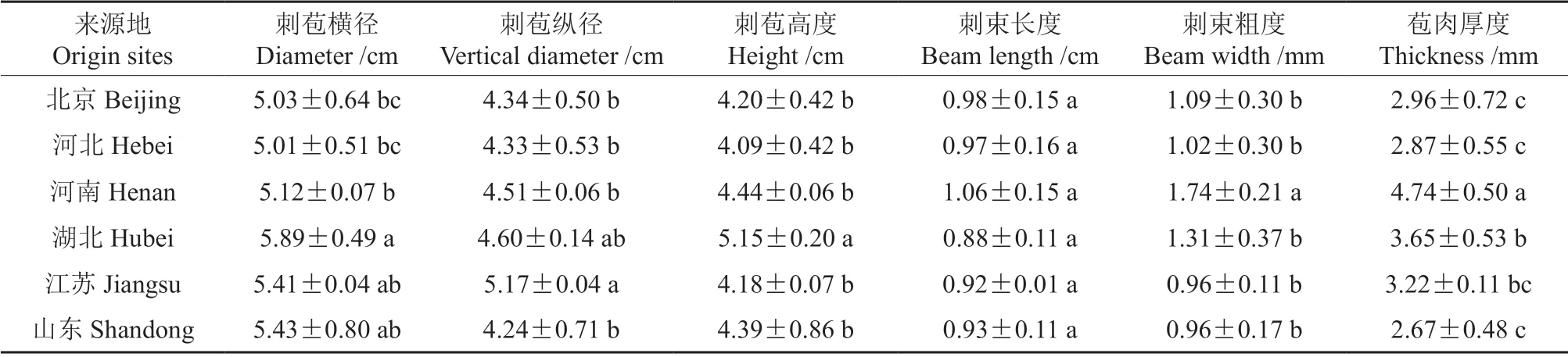
表4 不同板栗群体刺苞表型性状方差分析†Table 4 Variance analysis of phenotypic traits of thorns bract in different chestnut populations

表5 不同板栗群体坚果表型性状方差分析†Table 5 Variance analysis of phenotypic traits in different chestnut populations

表6 板栗刺苞、坚果表型性状群体间和群体内的方差分析Table 6 Analysis of variance between agronomic traits and population within chestnut thorns bract and nuts

表7 6 个板栗群体表型性状平均变异系数Table 7 Average coefficient of variation of phenotypic traits in six chestnut populations
2.3 板栗刺苞、坚果表型性状多样性来源
按双因素巢式分组设计方差分量比组成,进一步分析出各方差分量占总变异的比例,得到表8。结果表明:板栗表型形态性状的群体间方差分量分百分比在2.72%~27.60%,群体内方差分量百分比在44.72%~90.91%,板栗刺苞及坚果的表型形态性状在群体遗传组成上存在较大差异;各性状表型分化系数(VST)存在较大差异,表型分化系数(VST)值变化范围为5.21%~35.35%,平均值为18.13%,即群体间的平均变异与群体内的平均变异比约为2∶11;针对板栗总苞不同部位来说,板栗坚果表型分化系数(VST)均值为14.16%,板栗刺苞表型分化系数(VST)均值为18.31%。表明各性状群体内的变异远高于群体间的变异,群体内变异是板栗刺苞、坚果10 个表型形态性状呈多样性的主要来源。

表8 板栗刺苞、坚果表型性状方差分量和群体间农艺性状性状分化系数Table 8 Variance factors of phenotypic traits of chestnut thorns bract and nut traits and agronomic trait traits among populations
2.4 板栗刺苞、坚果表型性状聚类分析

图1 板栗群体树状聚类图(组间联接)Fig.1 Cluster tree of chestnut populations (Average linkage among groups)
采用系统聚类中的ward 聚类法,对6 个板栗群体39 份板栗种质资源进行聚类分析,如图1所示。在欧氏距离为5时,6个板栗群体被分为3大类,河北、山东、北京板栗群体聚为一类,河南、江苏板栗群体聚为一类,湖北板栗群体单独聚为一类。根据地理分布及我国板栗群体的划分:其中北京、河北、山东相互毗邻,属于地理上相近的群体,且北京、河北、山东板栗群体属于华北板栗品种群;河南、江苏大致处与同一纬度上,地理位置上相临近,同时江苏、河南板栗群体同属于长江流域板栗品种群,因此其表型相近,遗传上聚在一起,这与其地理分布格局大致吻合;湖北、河南、江苏同属与长江流域板栗品种群,但湖北地理位置靠南,与河南、江苏存在着纬度的差异,导致其气候条件出现差异,受到较大的环境压力影响,因此其单独聚为一类。
3 结论与讨论
3.1 结 论
板栗刺苞、坚果表型形态性状在各种质间存在显著差异,除刺束长度这一性状指标外,其余性状指标在群体间均存在显著差异。
板栗表型形态性状多样性在群体内和群体间存在丰富的变异。与刺苞表型形态性状相比,坚果表型形态性状的遗传稳定性高;各群体间,北京、河北、湖北、山东板栗群体遗传稳定性较低;各表型形态性状间,种皮厚度、刺束长度与粗度、苞肉厚度的平均变异系数较大,离散程度高。
根据双因素巢式分组设计方差分量比组成,表明群体内的变异远高于群体间的变异,群体内变异是板栗刺苞、坚果10 个表型形态性状呈多样性的主要来源。
在聚类分析中,将6个省份(群体)聚为3大类,河北、山东、北京板栗群体聚为一类,河南、江苏板栗群体聚为一类,湖北板栗群体单独聚为一类。其聚类结果与地理分布格局大致吻合。
3.2 讨 论
了解种质资源的遗传多样性是育种的工作前提与基础,对挖掘有益种质、引种栽培、发现新种质等多方面具有重要意义[17]。板栗坚果大小是反映板栗品种及其栽培价值的关键性状[18],该研究结果表明:板栗刺苞、坚果10 个表型形态性状在各种质间存在显著差异,除刺束长度这一性状指标外,其余9 个性状指标在群体间均存在显著差异。李颖[14]等人对不同省份板栗的结果母枝相关表型性状进行多样性分析,得到来源地区不同的板栗种质资源,其结果母枝的相关表型性状在群体间和群体内均存在显著性差异;周连第[19]等人通过对板栗自然分布区8 个产地的板栗叶片采集、测定并分析,其结果表明叶片各指标在同一群体不同品种间存在显著性差异,叶片形态在群体内和群体间差异显著;江锡兵[4]等人以10 个省份90 个板栗地方品种为研究对象,通过对其叶片表型、坚果表型、坚果品质的12 个重要农艺性状进行多样性分析,研究结果表明不同来源板栗的12 个板栗重要农艺性状在群体间和群体内均存在广泛变异,差异均达到显著水平,前人在板栗表型形态性状指标多样性分析中的出的结论与该研究结果相似。表明板栗坚果10 个表型性状多样性水平很高,育种选择潜力巨大,通过对现有板栗种质资源选择来改良刺苞、坚果表型是有可能够取得成功的。
变异系数(CV),又称“离散系数”,是概率分布离散程度的一个归一化量度,反映数据离散程度的绝对值,变异系数越大,则性状值离散程度越大[20],可用它来比较、衡量不同性状之间的变异程度。同时变异系数的大小也可也作为性状选择改良潜力的一个重要指标,决定着种质资源选择余地的大小[21]。本研究中,同一群体不同性状间及同一性状不同群体间的变异程度存在差异(表7),表型形态性状的差异是基因与生态环境共同作用的结果,说明群体间板栗的基因型和环境异质性导致群体表型变异的差异,也体现了群体间板栗种质资源在育种上的不等值性。刺苞表型形态性状的平均变异系数(11.24)>坚果表型形态性状的平均变异系数(9.27);种皮厚度、刺束长度与粗度的平均变异系数均>10%,其余表型形态性状的平均变异系数在5.37%~8.01%间,说明板栗群体内表型形态性状离散程度高,且坚果表型形态性状的遗传稳定性较大。北京、河北、湖北、山东板栗群体平均变异系数较大,其板栗表型形态性状多态性较为丰富,性状遗传稳定性较低,在很大的程度上能够选育出新种质,具有较大的遗传潜力。在育种工作中,考虑不同板栗群体平均变异系数大小,同时还要关注群体内的极性个体。例如,湖北群体在育种上具有选择出坚果形态较大种质的潜力,但在本研究中,单一性状较为突出的个体出现在北京和山东群体中。
生物群体间的变异在某种程度上反映了该生物对不同生存环境适应的广泛程度,生态位宽度的大小,值越大适应的生态环境越广[22]。该研究中,6 个板栗群体39 份板栗种质资源,群体间10 个性状指标的表型分化系数(VST)值介于5.21%~35.35%,平均值为18.13%,即群体间的平均变异与群体内的平均变异比约为2∶11,表明各性状群体内的变异远高于群体间的变异,群体内变异是板栗坚果10 个农艺性状呈多样性的主要来源。此研究结果与江锡兵[4]基于板栗叶片、果实表型性状群体间、群体内的平均表型分化系数研究所提出的群体内变异远高于群体间变异的研究结果相似。
在该研究中采用系统聚类中的Ward 聚类法,结果表明:河北、山东、北京板栗群体聚为一类,这3 个省份在地理位置上相互毗邻,又同属于华北板栗品种群;河南、江苏板栗群体聚为一类,江苏、河南板栗群体同属于长江流域板栗品种群,大致处于同一纬度上。聚类结果与地理分布格局基本相吻合,板栗群体基本按地理距离聚类,揭示了群体间表型性状变异的连续性,这与江锡兵[4]和周连第[21]等人研究结果相似。在该研究中,湖北板栗群体单独聚为一类,探究其原因可能是湖北与其他5 个省份均存在着纬度的差异,因此导致其气候条件出现差异,受到较大的环境压影响,因此其单独聚为一类。
生物表型形态变异是由其遗传组成和生态环境的共同作用下产生的,是群体生物遗传信息的重要表征,也是长期适应与进化的产物。环境与栽培管理措施对植物表型性状的影响较大,在相同的生长环境、栽培管理等条件下探究植物不同性状的多样性能够更好的研究其遗传本质[21-23]。在该研究的不足之处在于,所涉及到的试验材料来源于6 个板栗主产省份,试验材料定植于河北省迁西县东寨板栗种质资源圃,虽然保证了其生长环境、栽培管理等条件的一致性,但未能探究板栗表型形态多样性与来源地生态因子间的相关性。同时,栗属植物为雌雄同株,其雌雄花序较为明显,为异交植物,同时其分布范围有较大的重叠,存在自然杂交的可能[6]。因此在育种工作中应运用人工授粉、亲本回交等方法,在分子水平上利用不同类型的优良基因资源进行基因重组等,才能更加深入的揭示板栗的交配机制,这些问题均有待于进一步的探讨与研究。
