洞庭湖湿地植被长期格局变化(1987-2016年)及其对水文过程的响应*
2020-10-29万荣荣吴兴华
周 静,万荣荣,吴兴华,张 宇
(1:中国科学院南京地理与湖泊研究所,中国科学院流域地理学重点实验室,南京 210008) (2:中国自然资源经济研究院,北京 101149) (3:中国科学院大学,北京 100049) (4:中国长江三峡集团有限公司,北京 100038)
在《世界自然保护大纲》中,湿地与森林、海洋一起并称为全球三大生态系统[1]、是地球上水陆相互作用形成的特殊自然综合体、成为各种能流、物流和信息流的交换场所. 而湿地植被作为湿地生态系统三大组成要素之一[2],是主要的物质能量供应者,也是各种鱼类、鸟类和哺乳动物的觅食地. 湿地植被属性的变化意味着其他生物体生境的改变[3-4],是湿地退化和演替最直接的指示物. 而水文条件是影响湿地植物物种多样性、群落稳定性以及群落分布和演替的关键因子[5-6]. 大量研究表明,湿地植被类型与水位、淹水深度、淹没持续时间以及洪水频率等水情因子有关[7-10]. 一般而言,水位是决定植物分布的主导因素,其波动影响湿地洲滩土壤的含水量,直接导致洲滩植被面积和空间分布的改变,不同水深梯度下洲滩植物群落类型和生境有显著差异. 淹水的时空特征是湿地植被演替的重要决定因素,群落的差异分布是淹水条件差异及植被对土壤洪旱耐受力差异造成的,通常呈现明显的带状分布,是淹水持续时间及频率等水文过程共同作用下植物演替阶段的一种动态表现[11-13].
洞庭湖是我国第二大淡水湖,地处长江中游荆江南岸,是承纳湘、资、沅、澧四水、吞吐长江的兼具蓄、泄功能的洪道型湖泊[14],素有“长江之肾”的美誉. 洞庭湖受亚洲季风与江湖交互作用的复合影响,形成了较稳定的枯-涨-丰-退的季节性水位波动模式[15-16],近35年,洞庭湖年内水位变幅为6.6~15.4 m. 作为长江流域典型通江湖泊,洞庭湖水文情势复杂,其湿地植被也处于不断的演化之中[17-19]. 目前,洞庭湖湿地典型植被-水文的响应关系备受关注. 有研究显示,近二十年来,东洞庭湖湿地植被的面积在持续扩张、植被覆盖最小高程逐渐下降,并且三峡水库运行之后,这2种趋势在加快,这与洞庭湖水情条件的改变存在紧密联系[20]. Hu等[21]利用多时相遥感影像,探讨水情变化对东洞庭湖植被分布的影响,并分析三峡运行前后的差异. 唐玥等[22]通过遥感解译获得1989年以来东洞庭湖草洲出露面积的变化情况,发现低位洲滩草洲面积增加最快,而水位下降导致的洲滩出露时间变化是这一现象的主要原因. Deng等 使用多源遥感数据,分析洞庭湖植被与淹没频率的关系,得出了不同植被群落在淹没频率梯度上的分布差异. 除此之外,目前更多的研究偏向于机理方面:从形态学、解剖学、胁迫生理学等角度分析典型植物生长、分布的差异以及水情响应特征. 相关学者通过设计一系列析因实验,控制不同的水位或水深条件来观察洞庭湖湿地典型植物的相对生长率、生物累积与分配、根系特征、茎根比、茎直径、分支数以及总N/P含量等,进而探索湿地植被生长与分布对水情因子的响应[24-27]. 有学者分析洞庭湖湿地植被分布差异、格局变化的生物学机制,研究显示一方面湿地植被在不同水位下体内的可溶性糖、淀粉等多项生理指标存在差异性,这些差异可用来反映植物对洪水的忍耐力以及淹水后的恢复力,进而在一定程度上决定了湿地植被的分布[28-29]. 另一方面湿地植物还存在种内种间关系变化、繁殖对策等多种内在调控机制,如分布在低位洲滩的短尖薹草,其芽数、分株数等克隆繁殖特征会随水位降低而减少,从而影响植物生长[30-31]. 目前关于洞庭湖的研究多集中在东洞庭湖,全湖尺度上的研究相对欠缺. 此外,洞庭湖湿地植被水文过程的响应研究中,关于影响湿地植被变化的水情变量的周期性、波动性细节分析不足. 本文考虑到水文响应的滞后性,构建了枯、涨、丰、退以及水位最大值、最小值、平均值等多周期多层次的水情变量,在当前洞庭湖湿地植被水文响应研究中有十分重要的意义.
本文基于长序列遥感影像数据、水位日观测数据以及高精度湖盆地形数据,通过提取洞庭湖1987-2016年湿地植被信息,构建表征水位波动的多周期水情变量,阐明洞庭湖湿地典型植被的空间格局及多年变化趋势,并识别影响洞庭湖湿地植被分布格局的关键水情变量并建立其与植被面积的响应关系,分析水情对洞庭湖湿地植被格局的作用机制,以期为三峡水库调度管理、洞庭湖水资源利用及生态环境保护提供科学依据.
1 数据与方法
1.1 区域概况
洞庭湖(28°30′~30°20′N,111°40′~113°10′E)位于湖南省东北部,地处长江下荆江南岸,是我国第二大淡水湖泊(图1). 本文的研究区涉及的范围为洞庭湖湿地保护区规划界限以内的区域,总面积约2587 km2. 湖区可分为东洞庭湖、南洞庭湖和西洞庭湖3个不同的湖区,其地理概况介绍见表1[32].

图1 洞庭湖地理位置及水系分布Fig.1 Geographical locations and river system distribution of Lake Dongting

表1 洞庭湖不同湖区概况介绍
洞庭湖受季节性波动的流域和长江来水双重影响,年内水位波动巨大,呈现出“涨水为湖,退水为洲”的动态景观,年内洪汛期可持续3~4个月. 4月开始涨水,6-8月为丰水期阶段,水位达到最大值,9月中旬开始退水,12月至次年3月水位维持在较低水平[16]. 多年平均水位达25 m,年内水位变幅约12.5 m. 在季节性淹水的长期作用下,洞庭湖湿地呈现出浅水、草滩、泥滩多类型复合特征,土壤水分含量存在差异,洲滩上发育了丰富的湿地植被资源,且各典型植被群落按地势高低形成不规则的条带状或环状分布,不同植物占据特定的生态位空间[33].

图2 1987-2016年洞庭湖洲滩典型植被(a:苔草;b和e:杨树;c和d:芦苇)Fig.2 Representative vegetation of the Lake Dongting during 1987-2016 (a: Carex brevicuspis; b and e: Populus euramevicana; c and d: Phragmites australis)
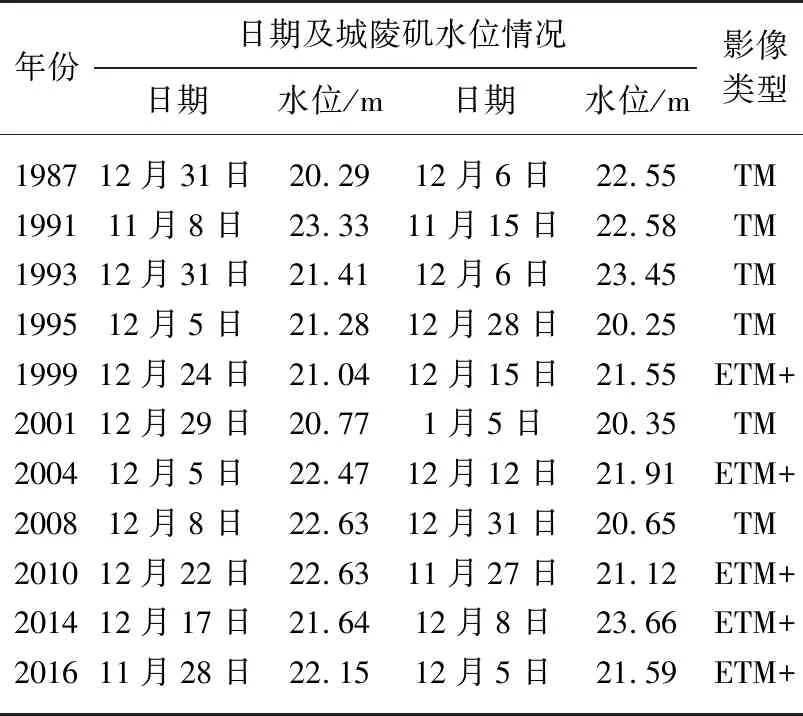
表2 影像类型及成像时间
1.2 数据来源
1)遥感数据:本文选取1987-2016年间无云条件下的Landsat TM/ETM+(空间分辨率30 m)遥感影像数据11期,共22景. 数据来源于USGS(http://earthexplorer.usgs.gov/),影像轨道编号和采集具体时间见表2. 考虑到秋冬季洞庭湖洲滩基本全部出露,且芦苇(荻)开始枯萎收割,在影像上与苔草易于区分,同时这一时期处于植被生长末期,植被生长波动较小,可保证湿地植被的年际可比性,因此影像采集时间全部在10-12月份之间.
2)野外实测数据:于2017年12月和2018年11月对洞庭湖区的植被分布、结构等状况进行野外实地调查. 调查地点涉及东洞庭湖、西洞庭湖、南洞庭湖保护区内湿地洲滩共8处,并用手持GPS导航定位详细记录各湿地类型的地理坐标、群落名称,共获取坐标信息近130个. 未能实地上岸定位的洲滩,通过拍照并在地图上详细标记,作为遥感影像解译提取的样本点.
3)水文数据:来自洞庭湖代表水文站城陵矶、鹿角、营田、杨柳潭、南咀、小河咀1987-2016年长序列日尺度水位观测资料(水位采用吴淞高程基准),来源于《长江流域水文资料》. 其中,城陵矶、鹿角站的平均水位用于代表东洞庭湖的整体水情,营田、杨柳潭和南咀、小河咀站平均水位分别代表南洞庭湖和西洞庭湖的水文情况.
4)地形数据:1995年1∶10000洞庭湖湖盆数字高程模型.
1.3 遥感影像解译
提取湿地景观信息前需对遥感影像进行一系列处理,主要包括辐射定标和大气校正、几何精校正、光谱增强、最佳波段选择、影像镶嵌与裁剪等工作. 本文以上处理过程基于ENVI 5.1平台实现. 另外,由于Landsat-7 ETM+机载扫描行校正器故障导致2003年以后获取的影像存在条带丢失,通过ENVI的Landsat_gapfill条带修复补丁进行修复[34]. 在目视解译和实地调查数据的基础上,将洞庭湖湿地景观分为5类,包括水域、泥滩、苔草滩地、芦苇滩地(洞庭湖区芦荻混生,为方便表述,文章统一用芦苇滩地(群落)代替)和林地滩地. 本文利用CART决策树算法进行影像分类提取湿地植被信息,实际操作基于ENVI软件中的CART扩展工具.
CART算法提出到目前已有30多年历史,被公认为是一种非线性、非参数的数据挖掘与分类的预测算法,预测模型精确可靠. 其原理是通过对由输入变量和输出变量构成的训练样本数据集进行循环二分构建二叉树并展开分类. 在决策树生长时,以Gini系数的减少量为测度指标选取使Gini系数减少量最大的属性为最佳分组变量对训练样本集进行分组,建立二叉决策树,其数学定义如下:
(1)
式中,G(t)为基尼系数,t为节点,k为输出变量的类别个数,P(j|t)是节点中样本输出变量取j的归一化概率. 假设用特征属性M将样本数据集N分为两组,这时的基尼系数为:
(2)
因此,分组后的基尼系数比原来的减少量为:
(3)
式中,G(t)和N分别为分组前输出变量的Gini系数和样本量,G(t1)、N1和G(t2)、N2分别为分组后右子树的基尼系数和样本量以及左子树的基尼系数和样本量. 当使两组输出变量值的异质性总和最小,即分组后样本类别变量尽量趋于相同类别值,纯度最大时,分组变量达到最佳分割点.
分类完成后利用ENVI中的混淆矩阵对影像进行精度评价. 后期处理基于ArcGIS 10.2平台进行计算,最终得到湿地植被面积及分布信息(图3).
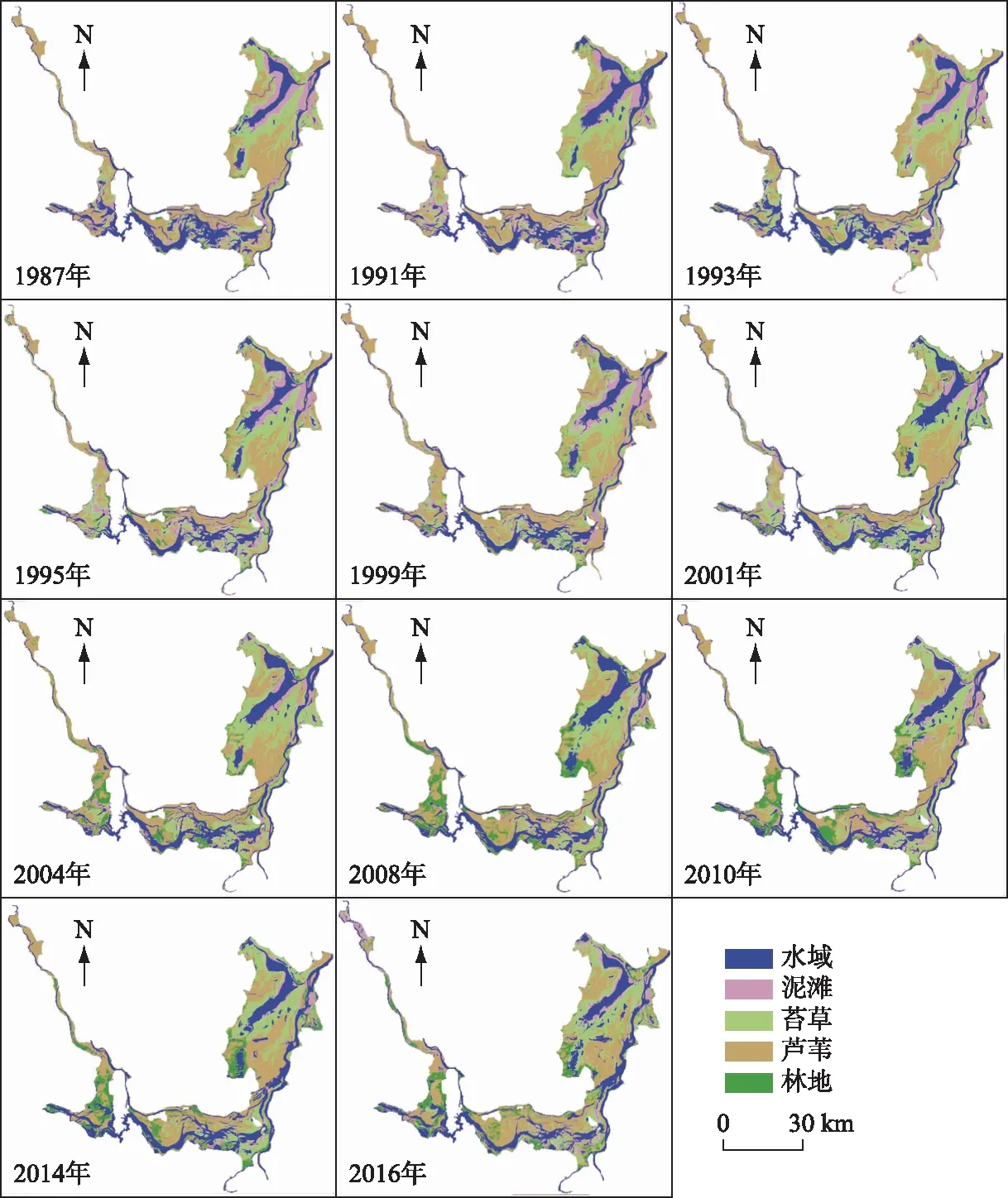
图3 1987-2016年洞庭湖湿地分类结果Fig.3 Classification of the Lake Dongting wetland during 1987-2016
1.4 洞庭湖湿地植被分布高程统计
在湿地环境中,淹水深度、淹水历时、淹水频率及土壤理化性质等的差异,会对湿地植物的生长繁殖、群落分布产生影响,因此湿地植被沿高程梯度占据特定的生态位空间. 高斯假说(Gause’s Hypothesis)认为,在生态学研究中,不同物种由于存在竞争其生长、繁殖最适环境范围总是相互分离[35]. 在最适范围以下或以上,物种所表现出的生理生态特征指标都会随之下降. 本文基于高斯假说理念,分析苔草和芦苇2个湿地典型植被的高程分布特征,由于林地群落主要是人为栽种,故不进行高斯拟合. 本文首先对洞庭湖地形数据以0.5 m为步长设置高程梯度,从19~33 m共28个梯度;然后,基于得到的植被类型数据,分湖区分别统计每个高程区间2种湿地典型植被群落的分布面积. 最后,对统计数据进行正态性检验,若满足条件,则得到洞庭湖湿地植被群落面积沿高程梯度的分布特征.
1.5 洞庭湖湿地水文过程的表征
洞庭湖水位呈现涨、丰、退、枯的周期性季节波动,水文情势的转变直接影响洞庭湖湿地植被的生物量、健康状况及空间分布等. 观测日期之前的枯水期、涨水期、丰水期以及退水期水位序列作为对湿地植被生态过程与景观格局特征有潜在生态意义的水位波动周期. 为了更全面量化水位波动对植被的影响细节,因此采用波动周期内的水位平均值、最高值、最低值以及水位变幅4个指标来刻画水位过程. 另外,统计大于不同水位梯度的淹水持续时间,来反映植被的淹水条件,不同湖区由于地势差异,故分湖区考虑其水位梯度,东洞庭湖统计水位大于22、24、26、28 m 4个梯度的持续时间,南洞庭湖统计大于25、27、29、31 m的水位持续时间,西洞庭湖统计水位大于28、29、31、33 m的持续时间,各湖区分别定义了20个水情因子,如表3. 以上水情指标基于城陵矶、鹿角、杨柳潭、营田、南咀、小河咀水文站的逐日径流数据来进行计算.

表3 描述水位过程的水情因子

图4 1987-2016年洞庭湖 湿地植被总面积变化趋势Fig.4 The total area of vegetation of the Lake Dongting during 1987-2016
2 结果与讨论
2.1 典型湿地植被面积变化趋势
近30年洞庭湖湿地植被面积的多年平均值为1669 km2,湿地植被分布总体上呈现增加趋势(R2=0.55),但2004年开始湿地典型植被总面积有所下降(图4),这一现象可能与当年秋季湖泊水位较高有关.
1987-2016年间在南洞庭湖和西洞庭湖,苔草群落面积呈现先增加后减少并在2004年以后趋于稳定,西洞庭湖其面积下降趋势更为明显,苔草群落面积基本维持在20 km2以内(图5a). 图5b为洞庭湖各湖区芦苇群落分布面积统计:1987-2016年间,东洞庭湖芦苇群落分布面积较稳定,多年平均值为434.27 km2;西洞庭湖芦苇群落分布面积有弱减少趋势,2000年前后,其芦苇群落面积分别为216.30、184.97 km2;而南洞庭湖其面积呈现微弱的上升,1987-1999、2001-2016年面积分别为304.86和315.67 km2. 总体上各湖区芦苇景观面积在进入2000年后趋于稳定. 近30年,洞庭湖各湖区林地面积快速增加,尤其是2001年以后,在2008-2016年间,林地扩张趋于稳定. 整个湖区,林地面积由1987年的45 km2上升到2016年313 km2,面积比例由1.77%上升为7.24%,林地面积增加主要发生在地势较高的湖岸大堤附近,包括东洞庭湖的漉湖附近,南洞庭湖湘江、资水入湖三角洲、万子湖北胜洲,西洞庭湖王塔洲、牛屎洲、大连湖和目平湖洲滩. 林地面积增加最快的区域为西洞庭湖,2016年该湖区林地面积占比达13.73%(图5c).
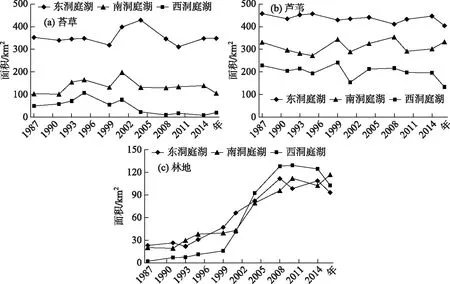
图5 1987-2016年洞庭湖各典型植被面积统计Fig.5 Area variation of typical vegetation types in Lake Dongting from 1987-2016
2.2 典型湿地植被分布高程变化
基于各湖区苔草和芦苇2种群落在特定高程梯度上的面积统计结果,分别计算1987-2016年间2种典型植被群落在最适高程范围内各高程梯度上的年际变化. 由图6可以明显看出,东洞庭湖苔草群落最适分布范围有下移的趋势. 苔草在高程24.66 m左右达到最大分布面积,根据近30年22.5、23、24、25、25.5 m高程上其面积的变化,发现22.5、24和23 m高程上,群落相对丰度均显著增加,而25和25.5 m高程上苔草群落面积有所下降,表明该群落最适生长范围有向湖心扩张的趋势. 在南洞庭湖,植被分布下限迁移这一现象并不明显,主要原因是该湖区地势较高,且河道曲折、洲滩遍布,苔草通常仅在湖岸有少量狭长分布,因此其高程变化趋势不明显. 西洞庭湖,各高程梯度上苔草群落面积均显著下降,无法说明该湖区苔草群落最适高程分布范围发生变化,除湖区地势因素外,另一原因是西洞庭湖近30年来苔草群落分布面积显著下降.
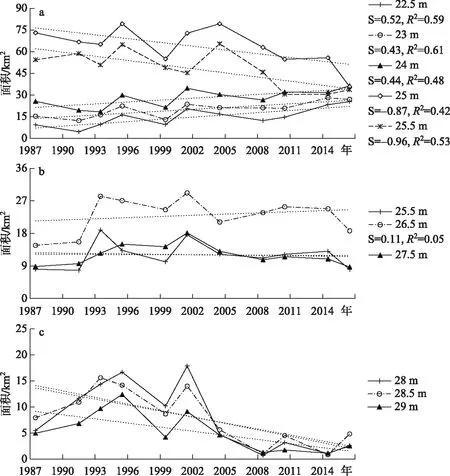
图6 1987-2016年洞庭湖不同湖区苔草各高程的面积变化(a:东洞庭湖;b:南洞庭湖;c:西洞庭湖)Fig.6 Area variation of Carex brevicuspis in different height during 1987-2016 (a: east Lake Dongting; b: south Lake Dongting; c: west Lake Dongting)
图7为近30年芦苇群落在最适高程范围内各高程梯度上面积的年际变化,东洞庭湖芦苇群落分布的多年最适高程范围在24.26~27.91 m之间,芦苇群落在26 m左右达到最大分布面积. 在东洞庭湖芦苇总体面积微弱减少的条件下,23、24、25 m高程上其分布相对丰度仍有所增加,表明东洞庭湖芦苇景观带分布最适高程范围呈下降趋势(图7a). 南洞庭湖芦苇群落分布的多年最适高程范围为25.22~28.57 m. 南洞庭湖芦苇群落在26、27、28 m高程上其面积分布均有所增加,表明南洞庭湖芦苇群落最适高程分布范围无显著迁移趋势(图7b). 同样,西洞庭湖芦苇群落最适分布范围变化不明显(图7c).

图7 1987-2016年洞庭湖不同湖区芦苇各高程的面积变化(a: 东洞庭湖;b: 南洞庭湖;c: 西洞庭湖)Fig.7 Area variation of Phragmites australis in different height during 1987-2016 (a: east Lake Dongting; b: south Lake Dongting; c: west Lake Dongting)
综合近30年洞庭湖湿地植被总面积及苔草群落、芦苇群落、林地群落的分布面积变化趋势可以发现,20世纪末,由于各典型植被分布面积变化并不显著,而2000年以后,林地人为大面积扩张,即使挤占了草洲的分布空间,湖区植被的总面积仍呈现增加趋势. 洞庭湖典型植被高程分布范围迁移在不同湖区存在较大差异,东洞庭湖趋势最明显,可能与湖区地形差异及近年气候变化、水利工程等导致湖区水情变化的空间差异有关[36]. 此外,洞庭湖水情偏枯[16],洲滩提前出露,淹没历时减少,进而导致植被带向下推移,而南洞庭湖与西洞庭湖地势本身较高,加上受人为干扰较大,因此湖区典型植被分布高程范围无明显变化.
2.3 洞庭湖湿地植被空间格局与水情的响应关系
通过Spearman相关系数法,计算洞庭湖各湖区湿地植被面积与相关水情因子间的相关性,结果显示东洞庭湖苔草群落和芦苇群落分布面积与水位波动变量间的相关性较大,而南洞庭湖与西洞庭湖湿地植被面积与水位波动因子间的相关性不显著, 原因可能是人类活动的干扰超出了湖泊水位波动对湿地植被分布的影响. 为进一步揭示东洞庭湖湿地植被对水情变化的响应关系,增加了6期影像数据(1989、1996、2003、2005、2006、2013年),进行湿地植被解译.
东洞庭湖各年份植被面积提取结果可以看出,1987-2016年间东洞庭湖湿地典型植被总面积有增加,其中苔草群落面积呈减少趋势,而芦苇群落处于扩张趋势,这一结论与Hu等[21]关于洞庭湖湿地植被变化趋势的研究结果一致(表4). 东洞庭湖苔草群落分布缩小的情况下,植被总面积在增加,是芦苇群落不断扩张并挤占苔草生长空间导致的. 李旭等的研究表明,土壤含水量随高程增加而减少,而洞庭湖荻-苔草群落的丰富度、多样性与土壤含水量有较好的相关性,苔草生物量表现出随高程的增加先增加后减少的趋势,荻生物量则呈增加趋势[37]. 因此结合三峡运行以来,洞庭湖水情偏枯的现象,可进一步分析是洲滩出露时间延长,滩地旱化导致了湿地植被由湿生植被向芦苇类旱地景观植被演替. 另外,可以看出植被面积与当年水位情况密切相关. 例如,2008年汛期较长,退水期平均水位仍有26 m,植物淹水时间过长影响其长势,植被总面积仅约757 km2. 而2006年、2013年则是典型枯水年,洲滩出露时间延长,植被面积总体较高,且由于土壤湿度降低,湿生植被生长会受到一定影响,面积下降,2006年苔草面积仅为299.31 km2.

表4 1987-2016年东洞庭湖植被面积统计(km2)
采用逐步回归分析法构建水情因子与东洞庭湖湿地植被面积之间的定量关系,揭示湿地植被分布随水位波动的变化规律. 依据逐步回归分析法,分别构建湿地典型植被苔草和芦苇群落面积与多周期水情因子的回归模型,见表5. 结果显示,CA2与水情因子有更好的拟合效果(R2=0.70),东洞庭湖芦苇面积受水位波动的影响更显著. 通过逐步回归识别影响湿地芦苇群落分布面积的关键水情因子有Fmax、Tmean、Rmean,回归模型均显示芦苇群落面积与丰水期最大水位、退水期平均水位及涨水期平均水位均为负相关关系. 可见丰水季节与涨水季节偏枯的水情对芦苇群落的发育生长有促进作用. 芦苇生长对水分的需求较小,只要土壤保持湿润即可,但汛期水位过高会不利于其生长,因此适度的土壤湿润有利于芦苇生长,而长期淹水则会起到反向作用. 影响苔草群落面积分布的关键水情因子为大于26 m水位持续时间以及丰水期的平均水位,但拟合效果稍差. Dur_26对苔草生长为促进作用,而丰水期平均水位则对其分布起到抑制作用. 前面研究结果显示该湖区苔草分布的最适高程范围在23~26 m之间,水位大于26 m的持续时间对湿生植被的生长发育有促进作用,但东洞庭湖丰水期平均水位达29.17 m,因此当汛期水位过高超过某一阈值时,将不利于湿生植被的二次生长.

表5 东洞庭湖植被面积与关键水情因子回归模型汇总

图8 苔草群落面积与关键水情因子的关系Fig.8 Relationship between Carex brevicuspis community area and key WLF variables
为进一步分析影响洞庭湖湿地植被分布的水位阈值,分别对CA1、CA2与通过逐步回归识别的关键水情因子进行多项式拟合,结果如图8、图9. 可以看出,苔草群落面积分布与Dur_26为线性关系,随着水位大于26 m淹水历时的增加,苔草面积呈增加趋势(P<0.05);而苔草面积与丰水期平均水位的多项式拟合结果显示,当Fmean在29 m左右,其分布面积维持在较高水平,当丰水期平均水位大于30 m,东洞庭湖湿地苔草群落面积将出现下降. 图9为芦苇群落与关键水情变量的多项式拟合结果,可知芦苇分布面积与丰水期最大水位以及退水期平均水位呈线性关系,且Tmean影响较弱,剔除左上角的2个点,可以看出随着退水期平均水位的变动,芦苇群落的面积基本处于小幅度波动,维持在350 km2左右. 而涨水期平均水位对东洞庭湖湖芦苇群落的面积有一个较明显的阈值,即当Rmean达到27 m以后,其分布面积将呈减少趋势. 反之,芦苇群落的生长将不会太受到影响,且涨水期水位的适度偏高,对芦苇群落的分布扩张有一定的促进作用.
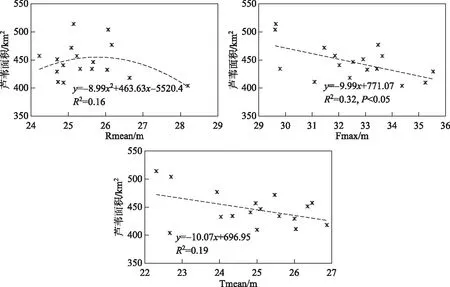
图9 芦苇群落面积与关键水情因子的关系Fig.9 Relationship between Phragmites australis community area and key WLF variables
3 结论
1)近30年,洞庭湖湿地典型洲滩植被面积在全湖尺度上呈稳定增加趋势,其中最为显著的是林地分布的快速增加. 湿地格局演变上,东洞庭湖呈现芦苇群落挤占苔草群落空间,并推动湿地植被整体向湖心扩张的趋势;而南洞庭湖与西洞庭湖其最适高程分布范围与高程下限均无显著变化,可能与该湖区地形特点关系较大.
2)丰水期、涨水期和退水期水情是影响东洞庭湖湿地典型植被带面积的关键水情变量,南洞庭湖、西洞庭湖湿地分布格局的水文响应关系不明显.
3)东洞庭湖湿地格局的水文响应关系表现为:①影响东洞庭湖苔草和芦苇分布最关键的水情变量是丰水期水位. 苔草对丰水期水情存在非线性阈值响应,丰水期平均水位维持在29 m左右,最适宜苔草生长;对于芦苇,丰水期偏枯的水情条件对其生长发育起到促进作用. 2种典型植被对丰水期水情因子的响应方式存在差异,苔草对丰水期平均水位敏感,芦苇对丰水期最大水位更为敏感. ②涨水期和退水期水文过程是影响东洞庭湖湿地植被分布的次为重要的水情因子. 涨水期、退水期水位偏低的水情条件对芦苇分布面积的扩张起促进作用.
