微小RNA-558 靶向调控叉头框转录因子C1 促进卵巢癌SKOV3 细胞的增殖和侵袭力
2020-10-25李静王金铭任琛琛杨立刘泇希白杨
李静,王金铭,任琛琛,杨立,刘泇希,白杨
卵巢癌是常见的妇科恶性肿瘤,在卵巢肿瘤中它的发病率占50%~70%[1]。在早期,卵巢癌多无明显表现,具有较高的病死率[1-2]。目前研究的主要理论认为:细胞的过度增殖和侵袭力的异常参与了肿瘤的发生发展[2],微小RNA(microRNA,miRNA)和卵巢癌的发生发展存在密切关系,通过对下游靶基因的调控表达,miRNA 影响了细胞的增殖和侵袭能力,可能导致卵巢癌的发生发展[3]。其中,miR-558 是目前被发现参与肿瘤发生发展过程的一种非常重要的新的癌基因,但是miR-558在卵巢癌组织中的表达以及miR-558 在卵巢癌发生发展过程中的作用机制尚不明确,并且miR-558和叉头框转录因子C1(FOXC1)之间的调控关系尚未见报道[4]。本研究主要探讨miR-558 在卵巢癌组织中的表达情况、miR-558 与FOXC1 基因之间的调控关系和miR-558 对人卵巢癌腺癌细胞SKOV3 细胞增殖、侵袭能力的影响,在卵巢癌的发生机制及治疗思路方面提供一些新的方向。
1 资料与方法
1.1 一般资料选择2014年1月至2017年12月郑州大学第三附属医院妇产科同期收治的25 例上皮性卵巢癌、25例交界性卵巢上皮性肿瘤和25例良性卵巢上皮性肿瘤病人,经过知情同意,取其手术切除的部分卵巢组织标本;上皮性卵巢癌组年龄范围为35~76 岁,临床分期Ⅰ~Ⅱ期6 例、Ⅲ~Ⅳ期19 例,中高分化10例、低分化15例;交界性卵巢上皮性肿瘤组年龄范围为39~65岁,临床分期Ⅰ~Ⅱ期12例、Ⅲ~Ⅳ期13例,中高分化13例、低分化12例;良性卵巢上皮性肿瘤组年龄范围为31~55岁。本研究符合《世界医学协会赫尔辛基宣言》相关要求。病人对研究方案均签署了知情同意书。由第四军医大学病原微生物学教研室惠赠卵巢腺癌细胞系SKOV3细胞。
1.2 方法
1.2.1主要试剂 从广州锐博科技有限公司购买miR-558 的模拟物、抑制物及阴性对照。miR-558、小核RNA6(U6)的特异性引物、FOXC1 的引物由上海生工生物工程有限公司合成。β-肌动蛋白(β-actin)抗体、FOXC1 抗体购买于美国Abcam 生物技术有限公司。实时荧光定量PCR 试剂盒购买于北京鼎国公司产品。
1.2.2验证 miR-558对FOXC1基因的调控作用借助于权威的生物信息学靶基因预测网站(PicTar、mi-Randa 和 TargetScan),将 miR-558 的名称和物种输入,预测与miR-558相关的靶基因FOXC1;通过荧光素酶报告基因实验,验证miR-558对FOXC1基因的靶向调控作用,构建荧光素酶报告基因载体质粒[(含有FOXC1 基因3’-非翻译区(UTR)中潜在结合位点)],野生型FOXC1基因3’-UTR(野生型-FOXC1-2-3’-UTR)和突变型FOXC1 基因3’-UTR(突变型-FOXC1-3’-UTR),放入-20 ℃冰箱保存备用。把质粒和miR-558共同转染到SKOV3细胞系中,培养48 h,用荧光素酶活性检测试剂盒测定并计算海肾荧光素酶与萤火虫荧光素酶的比值,以此值表示细胞荧光素酶的水平。
1.2.3细胞培养及转染 在RPMI1640 培养基中(含1%青霉素及链霉素及10%胎牛血清)加入SKOV3 细胞,恒温培养箱的温度为37 ℃,含5%二氧化碳,后续实验选取对数生长期的SKOV3 细胞;选取并计数对数生长期的SKOV3 细胞,按照2.0×105个/孔接种到6 孔培养板中进行培养。转染在细胞生长态势较好的时候进行。将转染实验细胞分成三组,分别为miR-558 模拟物组、miR-558 抑制物组及阴性对照组。将miRNA 转染到SKOV3 细胞中,miRNA 由羧基荧光素(Carboxyfluorescein,FAM)标记。转染之前,充分混合miRNA 和转染试剂,在室温下放置5 min,然后放置到培养箱的细胞培养板内,开始转染反应;转染24 h 以后,在200 倍光镜及荧光显微镜下分别观察并计数,把表达荧光蛋白的细胞作为阳性细胞,计算其转染效率,转染效率为阳性细胞数与总细胞数的比值。
1.2.4实时荧光定量PCR 技术检测 卵巢组织SKOV3 细胞中 miR-558 的表达水平以及 SKOV3 细胞中FOXC1 的mRNA 的表达水平将细胞的总RNA用Trizol一步法提取,浓缩,计算总RNA的纯度和质量。采用实时荧光定量PCR技术进行试验后,行融解曲线分析,每个实验均重复3 次。对结果采用2-△△ct法分析(ct值为反应的实时荧光强度明显大于背景值时的循环数)。
1.2.5蛋白印迹法检测 SKOV3 细胞中FOXC1 蛋白的表达水平提取总蛋白后测定浓度,依标准进行凝胶电泳,封闭液进行封闭,加入兔抗人FOXC1 一抗(1∶5 000 稀释),在室温下孵育2 h,然后洗涤3次,加入二抗,室温下再孵育1 h,洗涤3次,把β-actin 作为内参,分析其灰度值,计算FOXC1 蛋白的相对表达水平(FOXC1 蛋白与β-actin 的灰度值的比值)。该实验重复进行3次。
1.2.6CCK-8 法检测 转染后SKOV3 细胞的增殖情况将对数生长期的SKOV3 细胞接种于96 孔板中,在转染后24、48、72、96 h,采用CCK-8 试剂盒来检测细胞处于450 nm的吸光度(A)值并且记录。计算细胞增殖率[(实验组A值-空白对照组A值)/(对照组组A 值-空白对照组A 值)×100%]。本实验重复进行3次。
1.2.7基质胶侵袭实验检测 转染后SKOV3 细胞的侵袭力将转染后24 h的SKOV3细胞进行收集,细胞悬液加入到穿膜小室内,培养36 h,取出小室,擦拭掉上室细胞,把那些已经侵入并且贴附于下层微孔膜的细胞用多聚甲醛固定5 min,磷酸盐缓冲液(PBS)清洗后在显微镜下观察,随机在每张微孔膜中选5个视野进行细胞计数。此实验重复进行3次。
1.3 统计学方法使用SPSS 21.0 软件包对实验数据进行统计分析。计量资料用±s 来表示,所有数据均进行正态检验并且符合正态分布,采用单因素方差分析进行多组间的比较,采用独立样本t检验对两组间进行比较。P<0.05 时,差异有统计学意义。
2 结果
2.1 验证miR-558对FOXC1基因的靶向调节作用上述3个权威的生物信息学靶基因预测数据库预测得出miR-558的靶基因是FOXC1,荧光素酶报告基因实验显示,含有miR-558的质粒以及含有FOXC1基因的重组质粒共同转染SKOV3细胞后,模拟物组荧光素酶相对活性为(0.56±0.01),阴性对照组荧光素酶相对活性为(1.12±0.02),与阴性对照组相比较,模拟物组的荧光素酶活性下降了49.50%(P<0.05)。
2.2 卵巢组织中的miR-558 的相对表达水平比较实时荧光定量-PCR技术结果显示,上皮性卵巢癌、交界性卵巢上皮性肿瘤和良性卵巢上皮性肿瘤病人卵巢组织中miR-558 的相对表达水平分别为(3.43±0.42)、(2.47±0.35)、(1.37±0.31),各组之间两两比较,均差异有统计学意义(均P<0.05)。
实时荧光定量-PCR技术的结果显示,在miR-558模拟物组、抑制物组和阴性对照组中,SKOV3 细胞miR-558 的表达水平分别为(2.37±0.17)、(0.64±0.17)、(1.14±0.11),miR-558各组间两两比较,差异有统计学意义(均P<0.05);在miR-558模拟物组、抑制物组和阴性对照组中,SKOV3细胞的FOXC1基因转录出的mRNA的表达水平分别为(0.51±0.10)、(2.27±0.12)、(0.99±0.11),FOXC1的mRNA表达水平各组间两两比较,差异有统计学意义(均P<0.05)。
蛋白印迹法结果显示,在miR-558模拟物组、抑制物组和阴性对照组SKOV3细胞中,FOXC1蛋白的表达水平分别为(0.83±0.07)、(2.17±0.15)、(1.47±0.21),FOXC1 蛋白的表达水平各组间两两比较,差异有统计学意义(均P<0.05)。
CCK-8 结果显示,miR-558 模拟物组的细胞增殖率显著高于阴性对照组,抑制物组细胞的细胞增殖率明显低于阴性对照组,模拟物组中增殖率水平高于抑制物组,各组间两两比较,差异有统计学意义(均P<0.05),见图1。
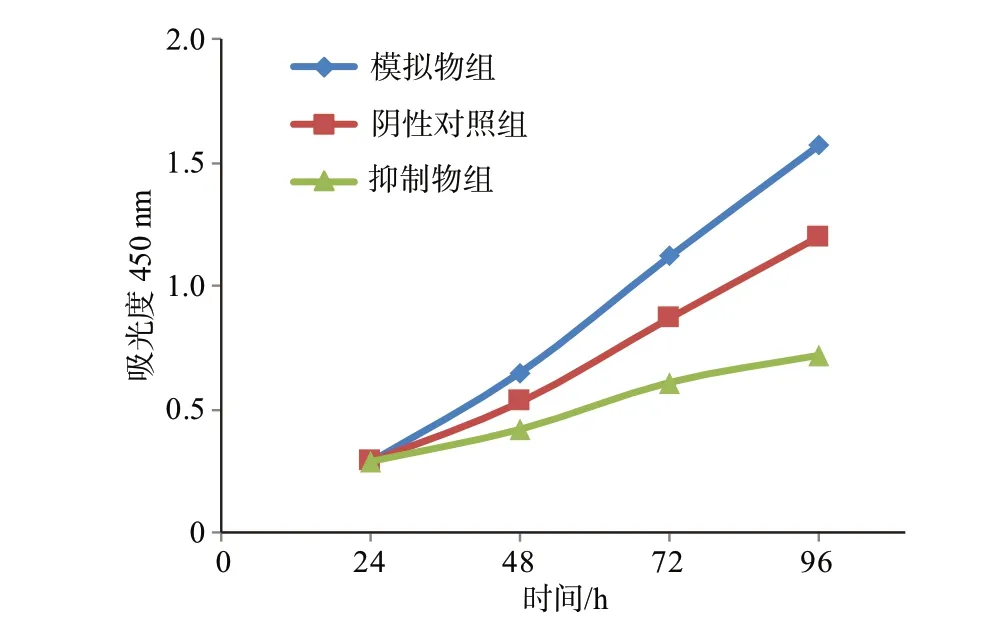
图1 卵巢癌组织CCK-8实验结果
基质胶侵袭实验结果显示,在miR-558 模拟物组、抑制物组和阴性对照组SKOV3 细胞中,SKOV3细胞穿透基底膜的细胞数分别为(158.33±9.45)、(67.01±10.58)、(117.67±16.86)(P<0.05)。miR-558细胞侵袭力水平各组间两两比较,差异有统计学意义(均P<0.05)。
3 讨论
卵巢癌的发病率位居女性生殖系统恶性肿瘤的前3 位,而它的死亡率却位居妇科恶性肿瘤之首[1]。据不完全统计,2015年,我国大约有5万多名女性被确诊为卵巢癌,约3 万例病人死于卵巢癌[1-2]。肿瘤细胞异常的增殖和侵袭能力是恶性肿瘤的主要生物学特征,细胞的增殖和侵袭能力是影响癌症治疗和预后的重要因素。肿瘤细胞异常的增殖力与侵袭力相互作用,共同促进癌细胞的生长,最终导致癌症的发展转移和化疗耐药[1,5-7]。因此,明确卵巢癌细胞增殖和侵袭能力过度的具体原因对控制卵巢癌的发生发展具有重大意义。腺癌是卵巢上皮性癌中的主要病理类型,本研究所选用的SKOV3 细胞系是卵巢腺癌细胞系能够更为真实地反映卵巢癌的生物学特性。
miRNA 是小分子单链 RNA,由 21 至 23 个核苷酸组成,位于内源非编码区,通过与靶mRNA 特异性的碱基配对,将靶mRNA 降解,或者抑制它的翻译,继而实现对目的基因的调控[3]。miR-558 是miRNA 中的一种,位于人类2 号染色体p22.3,能够调节环氧合酶2和白细胞介素-1β的表达,诱导人关节软骨细胞分解代谢[4]。有研究表明,miR-558可通过调控细胞的凋亡、增殖以及细胞的迁移、侵袭等生物学行为从而参与肿瘤的进展,如胃癌、成神经细胞瘤[8,10]。Li Y等[9]通过研究发现,在膀胱癌病人体 内 ,环 状 RNAHIPK3(circHIPK3)是 低 表 达的,circHIPK3 能够抑制miR-558 的表达水平,降低乙酰肝素酶的表达,从而调控细胞的迁移和侵袭及膀胱癌的血管生成,由此提示我们,miR-558能够促进膀胱癌的肿瘤发生。华中科技大学同济医学院的 Qu H 教授团队[10]通过研究发现 miR-558 能够与缺氧诱导因子2的5’-非编码区结合,促进缺氧诱导因子2 的表达从而促进成神经细胞瘤细胞的生长、迁移、侵袭和血管生成,从而促进肿瘤的发生,研究表明,高表达miR-558 的病人生存预后较差。同时Qu H教授团队[11]也在成神经细胞瘤的细胞中,通过敲除内源性miR-558 的表达,证实其能够通过活化乙酰肝素酶促进成神经细胞瘤的肿瘤生成和侵袭。华中科技大学同济医学院的Zheng L 教授团队[8]发现在胃癌的体内实验和体外实验中,miR-558 能够识别乙酰肝素酶启动子中的互补位点,以AGO蛋白家族成员1(Argonaute 1)依赖的方式结合SMAD 家族成员4(Smad4)降低Smad4的表达,从而促进胃癌的进展。这些研究与我们的结果相一致,从而更加明确了miR-558的致癌作用。
FOXC1 基因长约1787kb,位于人类6 号染色体p25.3,经转录翻译形成FOXC1 蛋白。最新的研究表明,FOXC1通过激活趋化因子受体4(CXCR4),能够参与癌症的发生发展,并且能够与活化T-细胞核因子1(NFATC1)构成基因遗传轴参与疾病的发展[12]。本课题组的前期研究已经通过QRT-PCR 技术和免疫组化法分析上皮性卵巢癌、交界性卵巢上皮性肿瘤和良性卵巢上皮性肿瘤病人的卵巢组织,明确在上皮性卵巢癌中FOXC1 的表达水平是降低的,与本次研究中的miR-558 的表达水平呈负向相关性[13-14]。同时本研究我们通过分析miRNA 靶基因预测(PicTar、miRanda 和TargetScan)数据库和荧光素酶报告实验明确了miR-558 对FOXC1 的靶向调控作用。本课题组通过前期研究发现在重组FOXC1慢病毒表达稳定的SKOV3的细胞中,细胞的增殖和侵袭能力受到明显抑制,在本次研究中我们进一步通过细胞实验明确了FOXC1 对卵巢癌细胞的增殖和侵袭能力的抑制作用是由miR-558 对FOXC1的靶向调控实现的[15]。
综上所述,miR-558 通过靶向调控FOXC1 的表达,能够促进卵巢癌细胞的增殖和侵袭能力,这为卵巢癌的发病机制提供了重要的理论依据。但是miR-558在卵巢癌病人血清中的具体表达情况以及在卵巢癌裸鼠移植瘤动物模型中的具体作用尚不明确,且与卵巢癌病人生存预后之间的关系尚不清楚,这仍然是本课题组下一步要深入研究的重点。
