岩石-土壤-四川牡丹元素迁移体系中的微生物效应
2020-10-14冯玮娜彭培好谢成晟
冯玮娜,彭培好,谢成晟,罗 雯
(成都理工大学 地球科学学院,四川 成都 610059)
土壤是地球关键带的重要组成部分,生物和非生物在此进行着高频率的物质交换和能量、信息的传递[1-2]。土壤对成土母岩有一定的物质继承性,土壤中大约80%~100%的钾、钙、镁和磷来自岩石的风化输入[3]。成土母岩是植物矿质元素的初始来源,成土初期,岩体微裂隙及表面附着的微生物和低等植物在营养需求的驱动下直接与岩石作用,在促进岩石风化释放矿质元素的同时固定碳,为植物的生长储备条件;土壤中的动植物及微生物生活产生的酸性和具有生命活性的分泌物促进岩石矿物的进一步风化[2-3]。快速的风化并不等于可利用元素的快速补充,早期风化产物难以发生化学变化,而微生物快速多样的代谢过程所塑造的氧化还原微环境则为元素形态转化、次生矿物的进一步风化提供了动能[4-5]。土壤微生物的结构和功能深刻影响着全球元素(主要是碳氮)大循环和岩石—土壤—植物间的元素小循环[6-7]。然而,由于微生物种类繁多、功能多样、微生物群落间、微生物与其它生物和非生物关系复杂,其在土壤中所承担的生态功能至今未得到清晰的描述[8]。
岩石风化与元素循环直接影响土壤营养结构,从而进一步影响植物生长和生态演化[9-11]。例如,中国干旱及半干旱区,土壤元素平衡(P,S,Be,Al,Mo,Pb,Cd,Ni)对禾本科、莎草科、豆科植物的生长有重要影响。中国干旱区,磷是控制草甸草原发展的关键因素之一[12]。四川牡丹是一种自然分布带狭窄的极小种群[13],多生长于营养贫瘠的坡积物上,其独特的基质条件和营养循环特点尚未得到深入研究。本文就四川牡丹生境的岩土元素组成特点进行研究,并描述几种大量、微量营养元素的迁移富集规律,分析细菌、真菌在元素迁移吸收过程中的效应,为了解营养元素的表生地球化学行为规律、微生物在元素迁移体系中的作用提供理论依据,同时为研究四川牡丹及其它干旱河谷地带性植被的基质条件与营养循环提供参考数据。
1 材料与方法
1.1 样品采集
四川牡丹生长于岷江及大渡河上游海拔2 204~2 800 m的干旱河谷地带,谷坡多在30°~40°之间,有的甚至更高。土壤层薄、石砾多、成熟度低。根据四川牡丹野外分布调查结果,于2018和2019年8月下旬在四川牡丹的5个集中分布区(见表1),分别选择3~5个四川牡丹分布较集中的地点划分5 m×5 m的样方。每个样方内随机选择3株四川牡丹,于地茎30 cm范围内根系分布密集处采集土壤,混合为1份土壤样品。四川牡丹生长的土壤层极薄,故在除去土壤表层枯落物后垂直挖取土壤截面至砾石层,不分层取样。同时使用经75%酒精消毒的铁铲、无菌手套等取样工具,取根茎抖落土壤于无菌密封袋中,并立即使用冰袋保存送回实验室。收集样方内所有植株已成熟的种子。对照区域地质图在四川牡丹垂直分布带的上、中、下部各寻找一处母岩出露处采集母岩样品。共取得35件土壤样品(包括12件根际土壤样品),17件岩石样品以及25件植物种子样品。

表1 研究区样点布置
1.2 元素含量测定
取岩石、自然风干后的土壤粉碎并过75 μm 200筛目筛,经105 ℃烘干2 h后使用硝酸、氢氟酸3∶2微波消解[14]。
植物种子先后于烘箱105 ℃烘干30 min,80 ℃烘干4 h,过60目筛,采用硝酸、双氧水8∶2湿法消解。所有消解后的样品经过滤,ICP-OES测定P,S,K,Na,Ca,Mg,Al,Fe,Mn,Cu的含量(岩石、土壤中的元素含量以氧化物计)[14]。取自然风干,过60目筛的土壤电位法[15]测定土壤pH值。
1.3 微生物基因测序
使用ZymoBIOMICSTM DNA Minipre Kit提取试剂盒提取根际土壤的细菌和真菌DNA,经PCR扩增及检测(扩增引物详见表2),产物回收后,使用TruSeq DNA PCR-Free Sample Prep Kit所建文库,于Illumina Hiseq 2500平台PE250模式测序。测序结果经过Trimmomatic2976质控及除杂,在97%的一致性水平上进行OTU聚类及分类注释[16]。

表2 土壤细菌及真菌PCR扩增引物序列[17-18]
2 数据处理
元素迁移累积系数=(X土壤/Al土壤)/(X岩石/Al岩石),表征某元素(X)相对于参比元素Al的迁移积累特征。植物吸收富集系数=(X植物/X土壤)×100%,表征元素从土壤到植物的生物吸收特征。土壤风化淋溶率以土壤、岩石的风化淋溶指数之比表示,指数包括:铝铁系数(Af)=Al2O3/Fe2O3,碱金属淋溶系数(Kn)=(Na2O+K2O)/Al2O3,碱土金属淋溶系数(Cm)=(CaO+MgO)/Al2O3和铝化率(α)=Al土壤/Al母岩[19]。微生物物种多样性(Alpha)指数包括丰富度指数ACE和Chao1,多样性指数Shannon,Simpson(此处以1-Simpson指数的值计)和InvSimpson[16,20]。
使用Canoco 5默认程序进行主成分分析和冗余分析,SPSS 25进行因子分析和Person相关性分析,R 3.6.1的corr.test函数对元素含量和微生物丰度进行Spearmen相关分析(p<0.05),并基于相关性结果使用igraph程序包构建网络图。
3 结果与分析
3.1 元素迁移富集特征
与华东地台沉积盖层相比,四川牡丹生长区母岩的Cu,S含量丰富,Ca,Mg含量较低(表3)。K,Ca,Na的最大值与最小值均相差四倍以上,反映各样地母岩的盐基储量差异较大。四川牡丹生长土壤pH值介于7.4~8.0之间(表中未列出),呈弱碱性。与全国表层土壤相比,四川牡丹生长土壤中S,Ca,Cu含量偏高,尤其是Ca远超过全国表层土壤钙含量的95%区间。S,Cu在各样点间含量差异较大,其最大值与最小值之比分别为4.89,3.24。植物种子中Al,Fe含量差异较大,最大值与最小值之比分别为3.08,3.45。
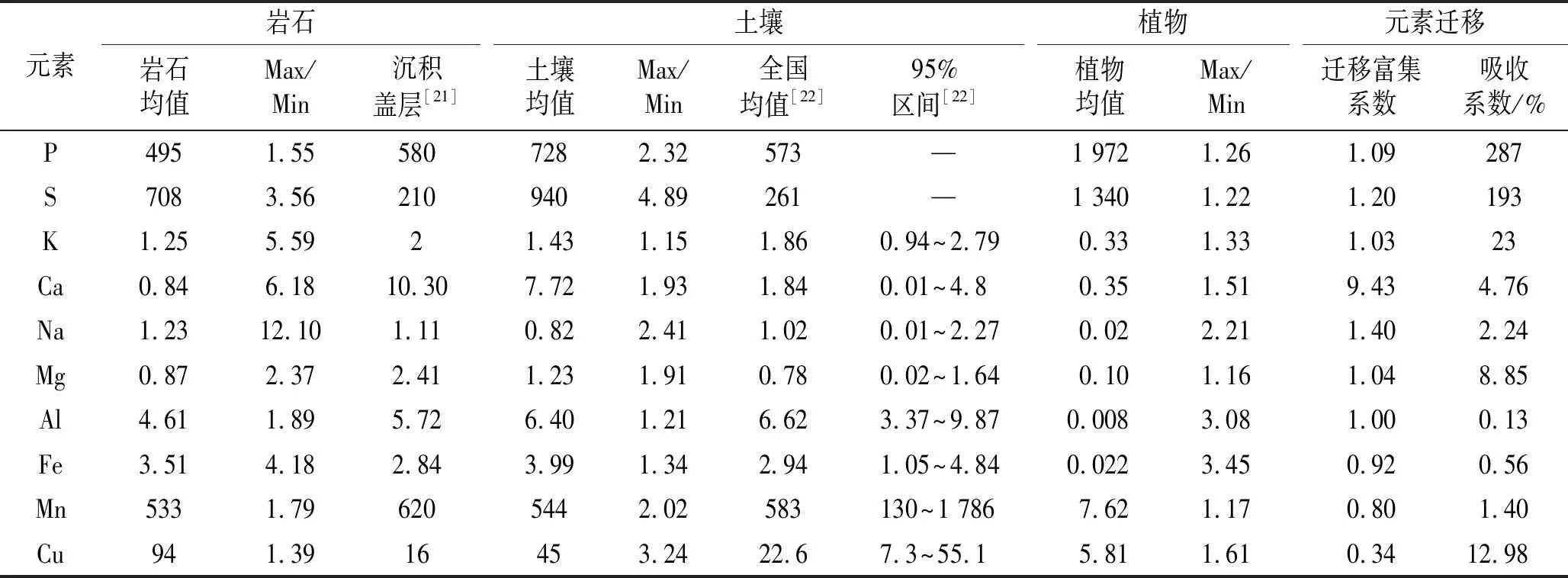
表3 研究区岩石、土壤、植物元素含量
土壤营养元素的迁移富集系数排序为:Ca>Na>S>P>Mg>K>Al>Fe>Mn>Cu。其中P,S,K,Ca,Na,Mg的迁移富集系数大于1,在土壤中相对富集,Fe,Mn,Cu的迁移富集系数小于1,表示其相对于Al发生了淋失。四川牡丹种子的元素吸收系数排序为:P>S>K>Cu>Mg>Ca>Na>Mn>Fe>Al。其中P,S的富集系数大于100%,表示其在四川牡丹种子中相对于土壤富集(见表3)。Al,Fe均是植物所需的微量元素,其吸收率均不到1%。
3.2 元素组合特征
成土过程中诸多因素的干扰使得相互影响的元素之间往往难以呈现出线性关系,通过主成分分析探讨元素间的潜在关联有助于理解成土过程中的元素行为规律[23]。母岩与土壤之间的元素组成差异由主成分1解释,各样地之间的元素组成差异由主成分2解释(见图1)。因子向量向主轴的投影为因子贡献率。相对于母岩,土壤以高Al2O3,CaO含量和低Cu,Na2O含量为最主要特征。这与元素迁移富集顺序稍有偏差,因为主成分分析更关注样点间的总体组成差异。岩石在各样点间的组成差异由主成分2解释,K2O,Na2O,Fe2O3是最主要的载荷因子,表明3者含量不同是母岩元素组成的主要差异。相较于母岩,各样点土壤的组成较为相似,这符合均一化理论,同时表明5个样点的成土过程基本相似。
因子1和因子2分别指示母岩中大量元素(氧化物)和微量元素(见图1)。主要存在两个元素组合:①因子1的主要正载荷因子为Al2O3,Fe2O3,S,K2O,P,CaO;②因子2的主要正载荷因子为Mn,Cu。
土壤中存在两个组合:①因子1的主要正载荷因子P,S,Mn,Cu;②一二象限的Fe2O3,K2O,CaO,MgO,Al2O3(见图1)。对比图1中的各元素组成特征可知,母岩与土壤的元素(氧化物)组合基本相似,反映土壤对母岩的物质继承性。不同之处在于土壤的P,S移入了Mn,Cu的组合中(其中P与Mn相关性显著,图3),表明两种元素在母岩和土壤中的主要存在形式可能不同。
此外,pH值与K2O,CaO同是因子1的主要负载荷因子,表明土壤酸碱度与土壤K,Ca的迁移积累可能存在相互影响(见图1)。因子1指示基质的营养限制,其主要正载荷因子Al,Mn,P(其中Al分别与Mn,P显著相关)在种子中的含量均受到岩石和土壤元素组成的影响;因子2的正载荷因子S,Fe,Mg,Cu均受到微生物活动的影响,其中S,Mg,Fe两两之间显著相关。Fe和Mg无论是在岩石、土壤还是植物中均在一个组合内,虽然两者没有显著的相关关系,但仍可以推测两者在迁移吸收过程中存在潜在相互影响。
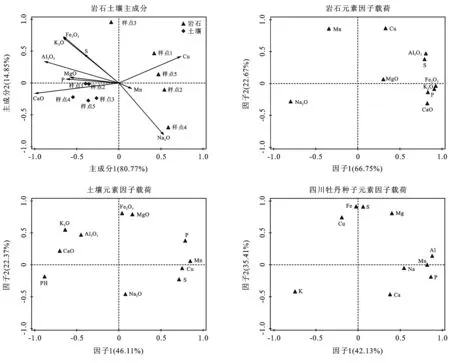
图1 岩石、土壤、植物种子的元素组成特征
3.3 土壤环境与微生物多样性
铝铁率、碱金属和碱土金属淋溶率越小,铝化系数越高表明土壤经历的风化淋溶程度越强,成熟度越高。钙的高度聚集(见表3)使得碱土金属淋溶率在此处并不适宜作为衡量土壤风化程度的指标,因此,碱土金属淋溶率仅作为衡量CaO,MgO相对淋溶或富集情况的度量。碱金属淋溶率在0.3~1之间,铝化系数和铝铁率多在0.8~2之间,总体表现为土壤淋溶程度较弱(见表4)。
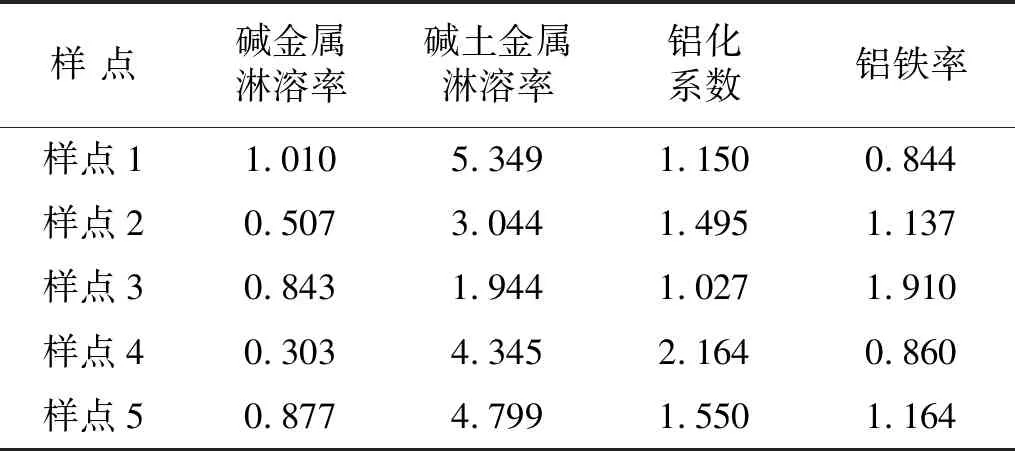
表4 研究四川牡丹生长区的土壤风化淋溶率均值
对细菌群落Alpha多样性和土壤风化淋溶率做冗余分析。前两个主轴共解释了总变量的74.68%(见图2)。4种风化率指数所指示的风化程度与微生物多样性之间的关系并不一致,表明四川牡丹生长土壤的风化进程并不是控制根际土壤微生物群落的主要因素。但是,碱土金属淋溶率(Cm∶F=4.7,p=0.036)和铝铁率(Af∶F=3.0,p=0.04)两个解释变量仍然可以很好地解释细菌的多样性。碱土金属淋溶率与细菌多样性指数(Shannon,Simpson和InvSimpson)呈正相关,与物种丰富度指数(ACE,Chao1)呈负相关,Af则正好相反。这表明随着土壤Ca,Mg,Fe相对积累程度加深,细菌多样性增加而丰富度反而有降低的趋势。所有指标均无法解释真菌的Alpha多样性,表明四川牡丹根际真菌对土壤环境变化的响应不如细菌敏感。
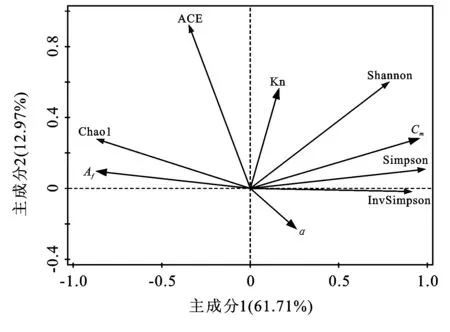
图2 土壤风化淋溶率与细菌多样性冗余分析
碱土金属淋溶率与细菌的拟杆菌门(Bacteroidetes)、Epsilonbacteraeota呈高度正相关,与硝化螺旋菌门(Nitrospirae)呈负相关,这三类细菌多具有与硝化相关的代谢途径(见表5)。铝铁率与真菌的座囊菌纲(Dothideomycetes)相关性显著,铝化系数与盘菌纲(Pezizomycetes)和伞菌纲(Agaricomycetes)相关性显著。上述菌类均是根际土壤中的优势菌种[16]。
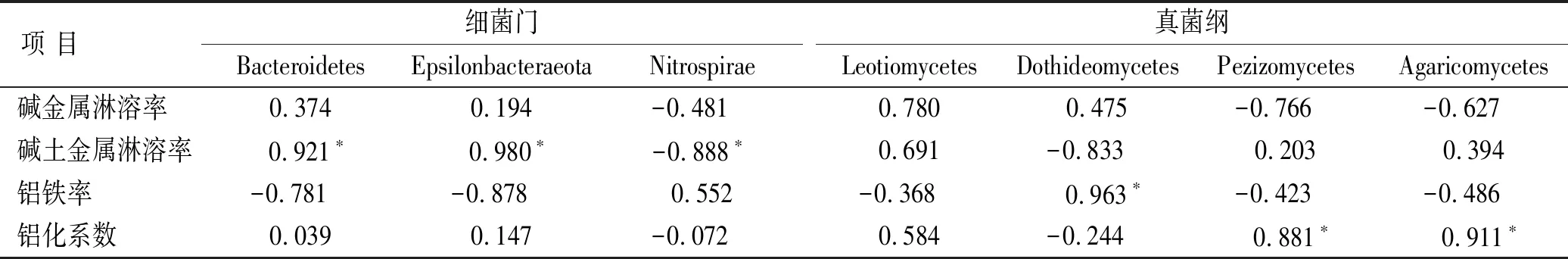
表5 土壤风化淋溶率与微生物相关性
3.4 网络分析
网络图便于反映微生物之间、元素之间、微生物与元素之间直接或间接的相关关系。模块1与微生物均无显著相关性,这部分主要包括岩石元素,以及土壤的P,Mn和植物的P,Al,Mn(图3)。说明P,Mn在岩石—土壤—四川牡丹种子的迁移体系中可能主要受控于母岩。微生物主要影响模块2,3,其中包括土壤元素:K,Ca,Mg,Al,Fe和种子元素:K,Na,Ca,Mg,Fe,S。此外,土壤S、植物Cu以及岩石Cu,Mn,Mg,P,S的含量同样与部分微生物的丰度显著相关。岩石和土壤微生物可通过土壤的桥梁作用产生相互影响,而这一过程如何发生在网络图中并未体现。
对比可知,Epsilonbacteraeota将岩石的Mn元素连接进入模块2中,细菌类的疣微菌门(Verrucomicrobia)、厚壁菌门(Firmicutes)、Latescibacteria和真菌类的粪壳菌纲(Sordariomycetes)、锤舌菌纲(Leotiomycetes)、盘菌纲(Pezizomycetes)、古菌根菌纲(Archaeorhizomycetes)作为一个整体将土壤Ca和植物Mg连接到模块2中,变形菌门(Proteobacteria)、螺旋体门(Spirochaetes)和座囊菌纲(Dothideomycetes)将模块3中的元素连接成为一个整体。上述微生物作为连接多种元素的节点,在营养反馈与调节上可能具有重要意义。微生物的加入使整个网络趋于复杂,它们不仅为元素间的反应提供了更多的途径还改变了元素之间的关系。细菌和真菌之间没有明显的分区,表明其在元素循环中的主要功能相似。
4 讨 论
(1) 四川牡丹的基质条件。四川牡丹生长土壤以弱碱积钙为主要特征。母岩钙含量低,不足以形成高钙土壤。其分布区少雨干燥的气候特点(年干燥度约1.6~2.5)或为土壤钙化提供条件[13]。除Ca在土壤中发生大量积聚外,K,Na,Mg相对于Al均发生轻微的富集。一方面,四川牡丹生长区受焚风效应影响,在高水分蒸发状态下,盐分可随着土壤毛细管道上移并在表土层积聚,这一现象易发生在干旱、半干旱及半湿润区[24]。另一方面,土壤中的盐基组分可能存在外源输入。四川牡丹生长区属于青藏高原东南缘,构造抬升导致山体破碎,河谷谷坡陡峭,水土流失严重,初期风化产物在重力及流水作用下堆积于谷坡凹陷或坡度减缓处发育成壤是植被发展的基础[25],谷坡上部土体及岩体经风化淋失的元素同样易在此汇集沉淀[26],最终可在植被、气候等作用下形成盐基组分的富集。相比于一般的植物吸收顺序,Cu较之Ca,Mg,Na位置较靠前,但4者含量均在正常范围内。
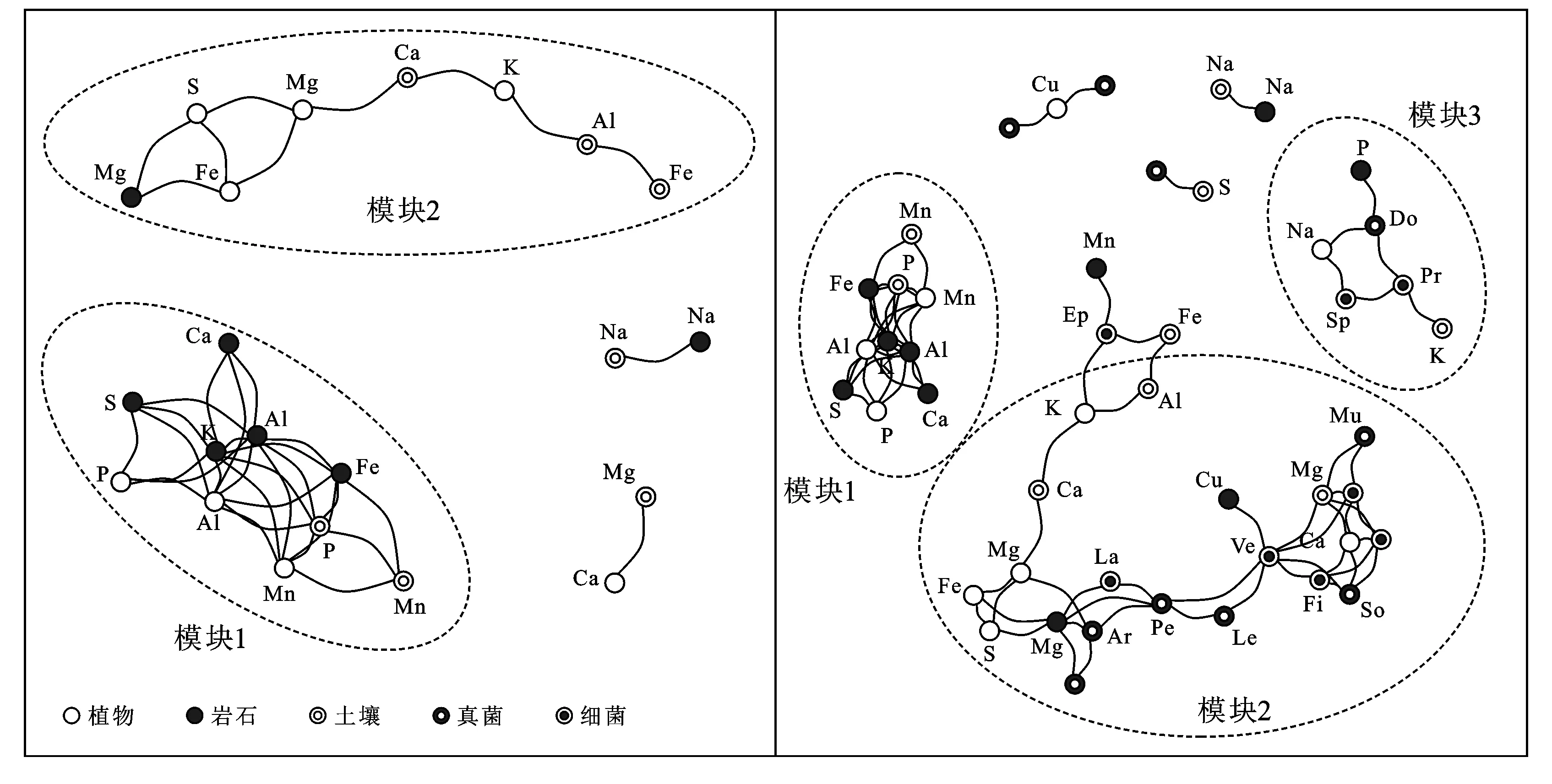
a 岩石—土壤—四川牡丹种子元素网络图 b 微生物与元素网络图
(2) 元素组合特点。P和S是表生系统中较为活跃的元素,两者组合关系的变化可能与其赋存形式、元素迁移动力等因素有关。岩石中的S与Ca,K,Al,Fe在同一组合内且相关性显著(图3),表明S可能主要以硫酸盐、硫铁化合物等形式存在于岩石中,而土壤中的S可能主要以Cu或Mn的硫化物形式存在(见图1)。碱性土壤中的P多以Ca-P的形式存在[25],但P与Mn的相关性说明土壤中可能有相当一部分P以Mn-P形式存在[27]。四川牡丹生长土壤常以坡积物为成土母质,土壤成熟度低,因此土壤元素组成得以保留其母岩的主要特征,其主要造岩元素组合关系未发生变化,这一特征在干旱、半干旱区等风化程度弱的区域较为常见[28]。
(3) 土壤环境与微生物多样性。一般而言,土壤微生物的多样性和丰富度随着土壤风化程度的加深而趋于增加[29]。这一点并未在四川牡丹的根际土壤微生物中体现,其原因可能在于微生物群落中普遍存在的结构冗余,即多个群落单元共同行使同一项功能的现象,使得微生物对环境变化有一定的承载能力,微小的环境变化与群落结构间往往没有确定关系[30]。但是,Ca,Mg,Fe的相对积累程度仍然是控制细菌多样性的主要因素之一。钙镁可通过改变微生物代谢活性及途径影响微生物群落结构。Rosenberg和Balland[31-32]通过控制试验证明钙镁可直接影响土壤微生物的生物量、活性,以及改变微生物的矿物风化能力。铁对微生物来说是理想的电子传递载体,微生物与Fe的反应控制着有机物矿化、反硝化等关键性土壤元素循环过程[33]。元素迁移对微生物的影响并非单向作用,如土壤中钙镁含量的变化可影响细菌从云母中提取铁的能力[32],进而推动多种元素的迁移。
(4) 微生物与元素迁移。本文以网络模块为单元分析微生物在元素迁移体系中的作用,模块2是整个网络的核心,几乎所有元素都与某类微生物或微生物群存在直接或间接的相关关系(见图3)。其中Ca,Mg,Fe,S是微生物链接进入该网络的主要节点,亦是微生物作用的主要对象。上述元素的相互作用间接影响着包括Mn,Al,K,Cu在内的诸多元素的活动。所有关键节点的真菌均属于子囊菌门,其中包括多种菌根和地衣,被认为是陆地植被进化的关键[34]。细菌中的厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)和变形菌门(Proteobacteria)广泛寄生于岩石中,对岩石的风化和土壤的初始形成有着重要作用[35]。其中以典型的铁氧化细菌——变形菌门(Proteobacteria)的希瓦氏菌(Shewanella)的研究最为广泛[2]。由于缺少诸如植物、流水、气候、地形等关键性因子,微生物与元素是如何发生相互作用的我们并未得知。
5 结 论
(1) 四川牡丹生长土壤以弱碱富钙为主要特征。元素迁移富集系数排序为:Ca>Na>S>P>Mg>K>Al>Fe>Mn>Cu;吸收系数排序为:P>S>K>Cu>Mg>Ca>Na>Mn>Fe>Al。
(2) 母岩元素组成特征在土壤中基本得以保留,P和S在成土过程中较为活跃。
(3) Ca,Mg,Fe的相对淋溶或相对积累程度是四川牡丹根际土壤细菌多样性的主要控制因素之一。
(4) P和Mn在岩石—土壤—四川牡丹种子中的迁移主要受岩石组分控制。细菌和真菌在元素迁移体系中的作用相似,主要参与Fe,S,Mg,K,Ca的相关反应过程。参与其中的主要细菌菌门包括Epsilonbacteraeota,Verrucomicrobia,Firmicutes,Latescibacteria,Spirochaetes,Proteobacteria。主要真菌菌纲包括Sordariomycetes,Leotiomycetes,Pezizomycetes,Archaeorhizomycetes和Dothideomycetes。
(5) 在干旱、碱性条件下,土壤化学反应相对较弱,多种矿物(元素)难以发生溶解,植物根系—微生物耦合风化作用常是有效的应对措施。这一过程十分复杂,过去常以模拟试验的方式进行研究,但这与自然状态下的过程相却甚远。本文直接测定四川牡丹生长土壤营养元素含量以及微生物组成,但这种方式难以揭示特定物种的作用。因此在门、纲水平进行研究,并不强调微生物与元素、矿物间的对应关系,而是试图寻找参与元素循环过程的主要微生物类群以及微生物与之作用的主要元素,以期为使用矿物、矿物—微生物等手段改善四川牡丹及干旱河谷地带性植被的基质营养条件提供指导或参考。
