干旱区不同降雨模式对藻结皮覆被区土壤碳释放的影响
2020-10-14王黎黎曹晓明
王黎黎,曹晓明
(中国林业科学研究院 荒漠化研究所,北京 100091)
全球气候变化,尤其是其与陆地生态系统的相互作用已成为当今生态学研究的焦点问题[1]。多个气候模型预测的结果一致显示,未来气候变化背景下,全球或局部地区的降雨格局将发生变化,高纬地区和热带太平洋区域降雨将增加,许多中纬度的地区平均降水也将增加[2-3];在一些地区,降雨格局可能进一步改变,表现为强降水和干旱等极端气候事件增多。CO2作为最重要的温室气体之一,其源、汇以及排放过程受到广泛关注[4]。
生物土壤结皮(biological soil crusts,BSCs,简称生物结皮)是干旱环境下自然发育的一种微生物共生体,它是由细菌、真菌等微生物和藻类、地衣以及苔藓与表层土壤颗粒进行复合作用而形成的特殊致密层并覆盖于地表[5],是荒漠生态系统组成和地表景观的重要特征。在干旱半干旱区,生物结皮的盖度占地表面积的40%[6],在一些受干扰较少的地区,其盖度可达70%[7],具有重要的生态功能[8]。生物结皮作为具有光合固碳活性的土壤与大气系统界面层,通过光合作用固定空气中的CO2,又以呼吸作用消耗土壤碳,同时伴随着生物体的分解矿化而影响着荒漠系统的碳循环[9],成为干旱半干旱生态系统土壤CO2通量的影响因子之一。生物结皮对外界环境尤其是水分因子的变化非常敏感,降水格局的改变势必会在一定程度上影响生物结皮的碳释放过程,进而影响其覆被下的土壤呼吸过程,乃至影响整个生态系统的碳循环过程。目前,国内外关于生物结皮碳释放的研究主要集中在生物结皮的碳释放速率[10-11]、生物结皮对土壤呼吸的影响[12-14]、生物结皮碳释放与温度、水分、土壤理化性质等环境因子的关系[8-11,15-18]等方面。干旱区土壤呼吸对降雨的响应已取得部分研究进展,特别是在降雨强度如何影响碳输入和输出方面。相比较而言,较大程度上忽略了碳过程对降雨时间分布响应的相关研究。藻结皮是生物结皮发育的初级阶段,是荒漠地区的拓荒植物和先锋植物,以适应极端环境条件、忍耐干旱、适应强辐射和适应贫瘠的土壤生境为特点,广泛分布在旱区严酷环境中,是乌兰布和沙漠重要地表覆盖景观特征之一。本研究以乌兰布和沙漠藻结皮为研究对象,探究3个降雨量水平×3种降雨频率的9种降雨模式下,藻结皮覆盖土壤的碳释放过程,以期为精确估算干旱区生态系统土壤碳释放量提供科学依据。
1 材料与方法
1.1 研究区自然概况
研究区位于内蒙古西部河套平原源头,乌兰布和沙漠东部边缘,隶属于巴彦淖尔市磴口县,地处东经106°9′—107°10′,北纬40°9′—40°57′之间。该县东西长约92 km,南北宽约65 km,总面积4 167 km2。地势东南高、西北低,海拔最高2 046 m,最低1 030 m。气候属温带大陆性季风气候,年平均气温为7.6 ℃,年平均降雨量144.5 mm,年均蒸发量2 397.6 mm。土壤类型以灰漠土、棕钙土和风沙土为主,整体表现出退化趋势。该地区为荒漠与干草原的过渡地带,植被类型为荒漠区沙生植被,主要物种为梭梭(Haloxylonammodendron)、白刺(Nitrariatangutorum)、油蒿(Artemisiaordosica)、柽柳(Tamarixchinensis)、杨柴(Hedysarummongolicum)、花棒(Hedysarumscoparium)、盐爪爪(Kalidiumfoliatum)、旋覆花(Inulabritanica)等。研究区内生物结皮广泛分布,主要包括藻类结皮、地衣结皮和苔藓结皮,其优势种分别为具鞘微鞘藻(Microcoleusvaginatus)、胶衣(Collematenax)和拟双色真藓(Byumargenteum),其盖度一般都在30%以上。其中地衣结皮和苔藓结皮大部分分布于植物覆被下,而在植物覆被外的绝大部分沙地上,藻结皮的分布占比最大[19]。
1.2 研究方法
1.2.1 试验设计 根据气候评估报告[2],预计到21世纪末,中国西北地区年降雨量将增加10%~60%,许多地方的降雨强度和降雨时间分布变化将进一步增加。基于此,本研究设计3种降雨量水平,以1986—2015年30 a间8月份的平均降雨量(5.26 mm)为基准(对照),设增雨30%(6.84 mm)和增雨60%(8.12 mm)两个增雨处理(见表1)。在以上3个降雨量水平下,各设置3种降雨频率,分别是基于1986—2015年30 a间8月份的平均降雨频率(6次,该地区8月平均降雨次数为5次,由于本试验从7月末开始,故增加1次)为基准(对照),设增加降雨频率50%(12次)和减少降雨频率50%(3次)两个处理。研究共9种降雨模式(3个降雨量水平×3种降雨频率),每种处理3个重复。增加降雨频率50%:将单次降雨平均分为两次降雨,减少降雨频率50%:将相邻两次的降雨叠加。保证同一降雨量水平下不同降雨频率处理下的总降雨量不变。为避免自然降雨对试验的干扰,用透明塑料布在样品上方搭建遮雨棚。

表1 试验设计
1.2.2 采集样品 2017年7月,选取地势平坦、远离植物、藻结皮发育良好的区域为采样区域。利用直径为20.3 cm(面积0.032 m2),高为20 cm的PVC自制采样器采集藻结皮覆盖原状土27个。样品采集前,用蒸馏水湿润结皮表面,以保证采集样品的完整性。
1.2.3 藻结皮覆被区土壤呼吸监测 为确保样品稳定性,采集后的样品按照算好的降雨量开始模拟降雨,但不测量土壤呼吸速率,待降雨一轮后,于2017年7月末至8月末,测定土壤呼吸速率。模式1,模式2,模式3共进行6次降雨试验,每次测定土壤呼吸速率。模式4,模式5,模式6共进行3次降雨试验,每次测定土壤呼吸速率。模式7,模式8,模式9共进行12次降雨试验,共测定9次土壤呼吸速率(另外3次只降雨,未测量)。降雨日上午6:00,利用LI-8150(LI-COR)土壤呼吸测量系统连接不透明气室测定降雨前的土壤呼吸速率。7:00开始,将计算好的水分用喷壶缓慢、均匀地喷洒在藻结皮土柱内,尽量避免产生径流,加水后立即开始测量,第0,1,2,4,8,12,16 h各测一次。
1.2.4 数据处理 采用时间加权法,计算不同降雨模式下藻结皮覆被区16 h土壤累计碳释放量和总累积碳释放量。采用单因素方差分析(one-way ANOVA)进行土壤呼吸月平均峰值、16 h土壤累计碳释放量差异性分析。采用双因素方差分析(two-way ANOVA)进行总累积碳释放量差异性分析。采用SPSS 20.0,Excel 2010软件进行数据的统计分析、制图。
2 结果与分析
2.1 不同降雨模式对藻结皮覆被区土壤呼吸速率的影响
降雨能够显著地激发土壤呼吸,各降雨模式下,土壤呼吸速率均在降雨后大幅度提升,并在1 h内达到峰值,之后逐步下降,到降雨后12 h左右基本恢复到雨前水平。其中,部分土壤呼吸速率曲线呈双峰或多峰波动,在降雨后4~8 h左右形成另一个小高峰,之后再逐渐下降至雨前水平(见图1)。随着降雨试验次数的不断增加,即随着干湿交替次数的不断增大,9种模式下藻结皮覆被区土壤呼吸速率的峰值逐渐降低,土壤呼吸速率曲线逐渐趋于平缓。第一次降雨试验,9种降雨模式下藻结皮覆被区土壤呼吸速率的峰值分别为3.82,3.98,4.38,4.29,6.52,7.18,2.51,3.42,4.20 μmol/(m2·s),最后一次降雨试验,9种降雨模式下藻结皮覆被区土壤呼吸速率的峰值分别为2.32,1.89,2.08,2.17,2.66,5.66,1.40,1.27,2.23 μmol/(m2·s),与第一次相比分别降低了40%~60%。
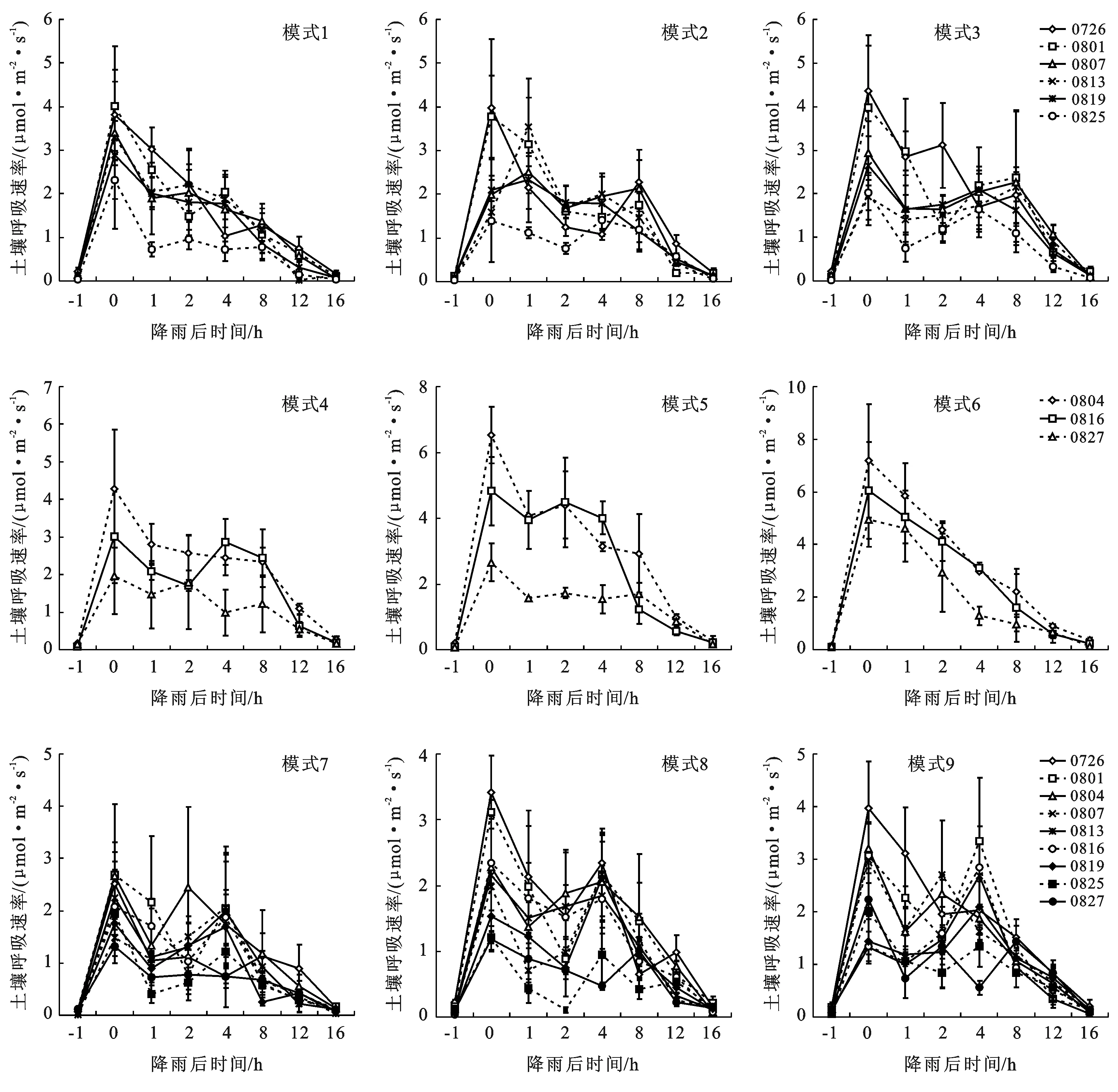
图1 藻结皮覆被区土壤呼吸速率
对比9种降雨模式月平均峰值大小依次为:模式6>模式5>模式1>模式4>模式3≥模式2>模式9>模式8>模式7(见图2),可以看出,降雨频率越低且单次降雨量越大,其土壤呼吸速率的峰值越高。
2.2 不同降雨模式对藻结皮覆被区降雨后16 h累积碳释放量的影响
9种降雨模式下,藻结皮覆被区降雨后16 h累积碳释放量总体上是随着单次降雨量的增大而增加。总体上来看,降雨频率减50%的3种处理>正常降雨频率的3种处理>降雨频率加50%的3种处理(见图3)。在正常降雨频率下,增雨30%(模式2)和增雨60%(模式3)显著大于正常降雨量(模式1)(p<0.05),但模式2和模式3之间差异不显著(p>0.05);在降雨频率减50%的情况下,增雨30%(模式5)和增雨60%(模式6)显著大于正常降雨量(模式4)(p<0.05),但模式6和模式5之间差异不显著(p>0.05);在降雨频率加50%的情况下,增雨60%(模式9)显著大正常降雨量(模式(7)(p<0.05),但模式7和模式8之间,模式8和模式9之间差异不显著(p>0.05)。
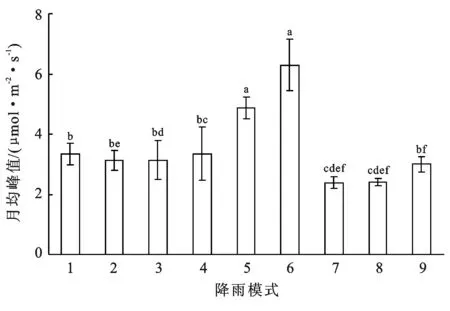
注:不同字母表示不同降雨模式之间差异显著(p<0.05)。下同。
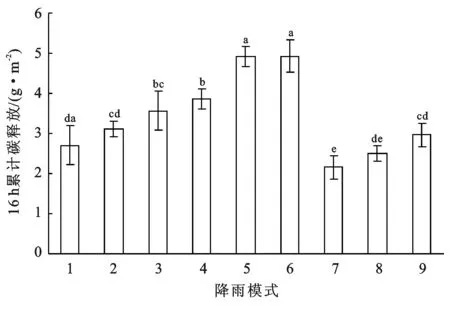
图3 藻结皮覆被区降雨后16 h累积碳释放量
2.3 不同降雨模式对藻结皮覆被区测定期总累积碳释放量的影响
降雨量和降雨频率都显著影响着藻结皮覆被区总累积碳释放量(p<0.05),但降雨量和降雨频率对藻结皮覆被区总累积碳释放量不存在交互影响(p>0.05)(见表2)。从整体上看,3种降雨频率下土壤总累积碳释放量表现为:降雨频率加50%>降雨频率减50%>降雨频率正常,且3种频率间差异显著(p<0.05)。总累积碳释放量随着降雨量水平的增大而增加,其中增雨30%和增雨60%情况下的总累积碳释放量显著大于正常降雨量水平(p<0.05),但增雨30%和增雨60%间的差异不显著(p>0.05)。在正常降雨频率下,正常降雨量(模式1)、增雨30%(模式2)、增雨60%(模式3)总累积碳释放量分别为26.97,30.78,32.39 g/m2,增雨30%(模式2)和增雨60%(模式3)总累积碳释放量较正常降雨量(模式1)分别增加了14%和20%;在减少50%降雨频率下,增雨30%(模式5)和增雨60%(模式6)总累积碳释放量较正常降雨量(模式4)分别增加了12%和15%;增加50%降雨频率下,增雨30%(模式5)和增雨60%(模式6)总累积碳释放量较正常降雨量(模式4)分别增加了18%和37%。在正常降雨量水平下,降雨频率减50%(模式4)和降雨频率加50%(模式7)比正常降雨频率(模式1)总累积碳释放量分别大了25%和30%;在增雨30%水平下,正常降雨频率(模式2)、降雨频率减50%(模式5)、降雨频率加50%(模式8)总累积碳释放量分别为30.78,37.65,41.20 g/m2;在增雨60%水平下,正常降雨频率减50%(模式6)、降雨频率加50%(模式9)总累积碳释放量分别38.76和47.99 g/m2,是正常降雨频率(模式3)的1.2和1.5倍(见图4)。
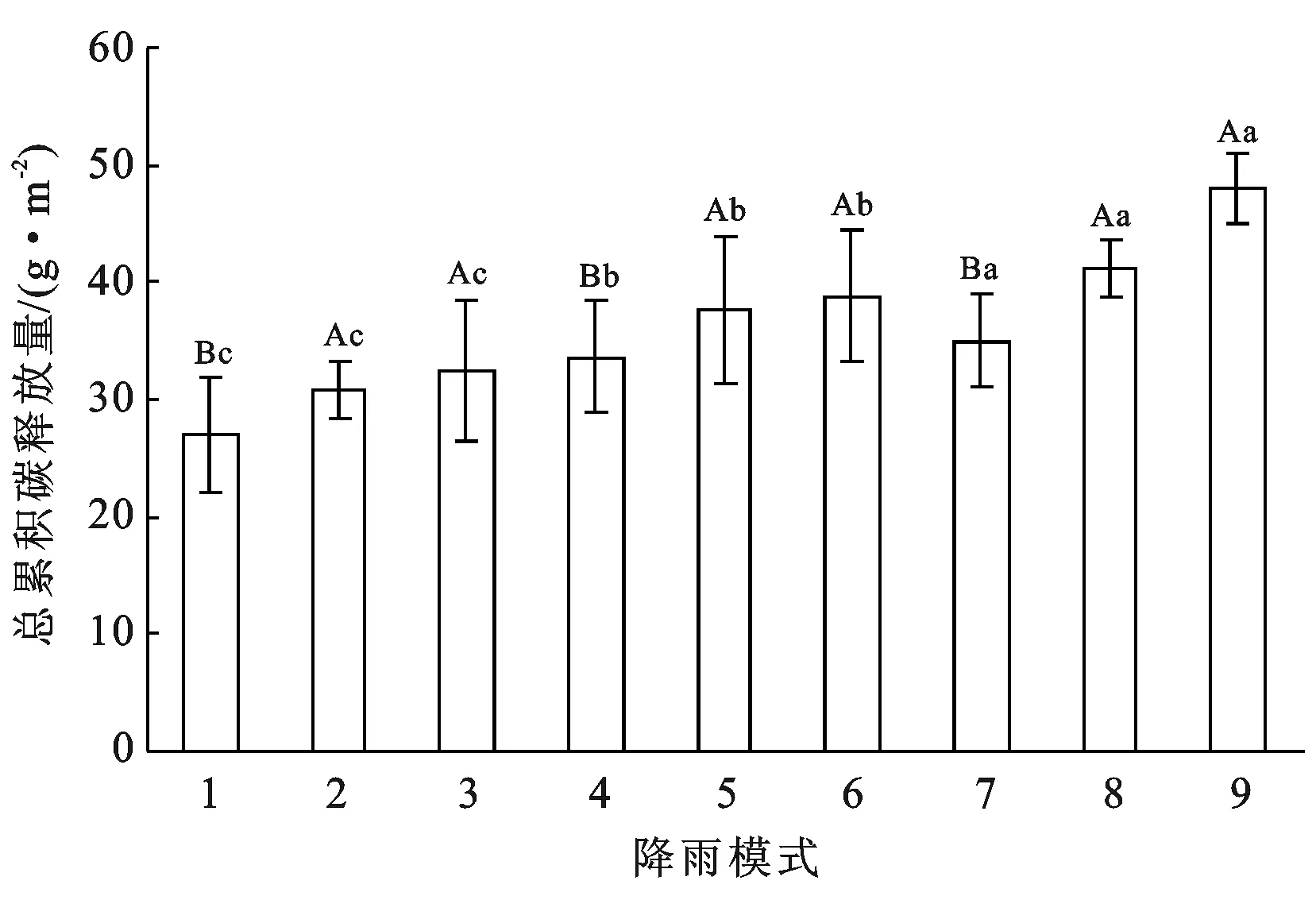
注:不同大写字母表示不同降雨量之间的差异显著(p<0.05),不同小写字母代表不同降雨频率之间的差异显著(p<0.05)。

表2 降雨量和降雨频率对总累积碳释放量影响方差分析
3 讨 论
在干旱区,水分是生态系统生物活动的主要限制因子,降雨可通过改变土壤的干湿状况直接影响土壤的生态过程,继而引起土壤碳库的变化。生物结皮中几乎所有的生物都是变水性[20],对水极度敏感。本研究表明,降雨能够刺激藻结皮覆被区土壤呼吸速率迅速大幅度提升,之后随着时间的推移逐渐降低至较低水平。这与前人的研究结果一致[21-22]。这可能由以下几个原因所导致:首先,在干旱环境中,生物结皮一般呈干燥休眠状态,降雨后,结皮能够在短时间能迅速恢复生理活性,进行呼吸作用。其次,干旱区土质松散,干旱少雨,土壤空隙中可能积累了一定量的CO2,雨水的渗入将这部分CO2排出[23]。此外,还与土壤呼吸底物数量、微生物的活性状态等密切相关。但随着干湿交替次数的不断增大,藻结皮覆被区土壤呼吸速率的峰值逐渐变小,即土壤再湿润后所产生的呼吸脉冲逐渐减弱[24]。对比9种降雨模式月平均峰值大小,减少降雨频率50%并增雨60%(模式6)的峰值最大,其次为减少降雨频率50%并增雨30%(模式5),说明,在一定的时间范围和降雨量水平下,降雨间隔时间越长,单次降雨量越大,所产生的呼吸脉冲越强。这可能是由于经过不断地干湿交替过程,土壤中可利用的有机质、土壤中微生物生物量、微生物群落结构的变化所导致的。呼吸脉冲的大小直接影响着降雨有效期内的累积碳释放量。本研究中,9种降雨模式下藻结皮覆被区降雨后16 h累积碳释放量即随着单次降雨量的增加而增大。低频率、大雨量的降雨事件所引起的碳释放量明显高于高频率、小雨量的降雨事件。主要是由于干旱区蒸发量大,小雨量的降雨事件仅能湿润土壤表面,短时间内就会被蒸发掉。但将时间尺度延长到月,在相同的总降雨量下,高频率的小降雨事件所产生的总碳量最高,其次为低频率的大降雨事件,正常降雨频率下最小。在一定时间内,低频率的大降雨事件所产生的总碳量大于正常降雨频率,这与赵荣[25]等的研究结果一致。但高频率的小降雨事件所产生的总碳量最高,说明降雨频率对总碳量的影响是呈“V”字形的。同样在月的时间尺度上,相同的降雨频率下,生物结皮覆被区的碳释放量随着降雨量的增大而逐渐增加[26-29],但当降雨量增大到一定程度后,其对土壤呼吸的促进作用不再明显。本研究中,相同降雨频率下,增雨30%和增雨60%情况下的总累积碳释放量显著大于正常降雨量水平,但增雨30%和增雨60%间的差异不显著。有研究表明,土壤水分过多会抑制土壤呼吸[30-31],因此,降雨量过大可能会对土壤呼吸起到抑制作用,关于这部分问题,有待将来进一步研究。
4 结 论
(1) 降雨对藻结皮覆被区土壤呼吸速率具有显著的激发作用,土壤呼吸速率在降雨后立即大幅度增加,之后逐渐降低至较低水平。但随着干湿交替次数的增加,土壤呼吸速率的峰值逐渐变小,土壤呼吸速率曲线逐渐趋于平缓。
(2) 藻结皮覆被区降雨后16 h累积碳释放量总体上是随着单次降雨量的增大而增加,但当降雨量增大到一定程度后,其对土壤碳释放量的促进作用不再明显。
(3) 降雨量和降雨频率都显著影响着藻结皮覆被区总累积碳释放量,但降雨量和降雨频率对藻结皮覆被区总累积碳释放量不存在交互影响。在相同降雨频率下,总碳释放量随着降雨量的增加而增大。在相同的总降雨量下,高频率的小降雨事件所产生的总碳量最高,其次为低频率的大降雨事件,正常降雨频率下最小。因此,由气候变化所引起的降雨量增加和降雨频率的变化将会增加藻结皮覆被区的碳排放量,在预测碳收支时,也应将生物结皮的碳排放量变化作为考虑因素之一。
