一株具有溶藻特征的假单胞菌分离鉴定及其溶藻动力学研究
2020-10-13王美娟吴凌云许明宸黄金杰毛林强张文艺
王美娟 吴凌云 许明宸 黄金杰 毛林强 张文艺


摘 要:从田螺内脏中分离获得一株溶藻菌株(命名为W10),利用16S rDNA序列分析及生理生化特征对该菌株进行了鉴定,探究了其对铜绿微囊藻的作用方式、菌液及其無菌上清液在不同条件下的溶藻特性,并构建了溶藻动力学模型。结果表明:菌株W10归属于假单胞菌属(Pseudomonas sp., GenBank ID为MN688696);W10溶藻式是通过分泌某些热稳定性差的溶藻物质间接作用于铜绿微囊藻,使其裂解、溶解;NA和淀粉培养基对W10菌株的培养效果无显著差异(P>0.05),二者显著优于改良基础培养基(P<0.05),但因NA培养基成分复杂,故确认淀粉培养基适宜于菌株W10培养;同一生长时期的菌液及其无菌上清液溶藻效果无显著差异(P>0.05),二者均表现为稳定期与衰亡期最高,二者无显著差异(P>0.05),然后依次是对数期和延滞期;当菌液与藻液体积比为1∶10时,W10菌液溶藻率最高为80.05%,而无菌上清液的溶藻效果与投加量呈正相关,在1∶2处理溶藻率最高为92.15%;当菌藻比1∶10以铜绿微囊藻为降解底物时,菌株W10的溶解作用遵循一级反应动力学。
关键词:溶藻细菌;铜绿微囊藻;分离鉴定;溶藻特性;溶藻动力学
中图分类号:X703 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2020.07.005
Abstract: An algicidal bacteria strain W10 was isolated from a snail's viscera and identified based on 16S rDNA gens and biochemical and morphological characteristics. The algae-lysing way of W10 and the effects of different conditions on the algae-lysing action were investigated to analyze the algicidal characteristics of strain W10 and its filtrate, and then built the algae-lysing kinetic model of W10. The results showed that strain W10 was identified as Pseudomonas sp. (Accession No.MN688696). And W10 indirectly attacked Microcystis aeruginosa by secreting algicidal substances with poor thermal stability. There was no significant difference of algicidal ratio (P>0.05) between NA and starch medium, both of which were significantly better than the modified basic medium (P<0.05). However, due to the complex composition of the NA medium, it was confirmed that the starch medium was suitable for cultivating W10. There was no significant difference of algicidal ratio (P>0.05) between W10 and its filtrate at the same growth phase. The algae lysing of W10 and its filtrate were highest in the stable growth phase and the decay growth phase. There was no significant difference of algicidal ratio(P>0.05) between the stable growth phase and the decay growth phase, followed by the logarithmic growth phase and the delay growth phase. Furthermore, when the volume ratio of W10 to Microcystis aeruginosa was 1∶10, the highest removal rate of W10 was 80.05%. But the algicidal effect of W10's filtrate was positively correlated with the dosage; the highest removal rate of W10's filtrate was 92.15% in the 1∶2 treatment. When the bacteria-algae ratio of 1∶10 and Microcystis aeruginosa was used as degradation substrates, the dissolution of Microcystis aeruginosa by W10 followed first-level kinetics.
Key words: algicidal bacteria; Microcystis aeruginosa; isolation and identification; algicidal characteristics; algae-lysing kinetics
藻類在富营养化水体中大量繁殖,形成有害藻华(HABs)导致水质恶化、鱼类死亡,甚至会产生藻毒素对水生动物和人体造成直接危害[1],近年来,由于其严重的生态效应和负面的经济影响,引起了全世界的关注[2-3]。细菌等微生物能控制自然环境中藻类的生物量,调节藻类的动态平衡,这种藻-细菌的相互作用使得利用溶藻细菌对藻华进行生物控制成为可能[4-6]。
溶藻细菌按溶菌方式分为直接溶菌和间接溶菌[7-8],迄今为止发现的溶藻细菌约70%为间接溶藻,其余30%需要与藻细胞直接接触[9]。史荣君等[10]从赤潮爆发海域沉积物中分离出的芽孢杆菌N3,可以使锥状斯氏藻和海洋原甲藻的藻细胞变形膨胀,最终破裂死亡,属于直接溶藻。Tian等[11]从蓝藻水华爆发时期的太湖水样中筛选出对铜绿微囊藻有去除效果的细菌A27,结果表明,A27溶藻方式为间接溶藻,可分泌至少2种不同的溶藻活性物质,达到抑藻的目的。
国内外有关溶藻细菌的筛选及其溶藻特性研究报道众多,但是溶藻细菌的分离源大多为蓝藻频发的自然水体水样或沉积物[12],菌种来源较为单一。田螺在自然环境中可摄食藻类、细菌和浮游生物等,何健等[13]将田螺养殖在生活污水中,实验组栅藻的增长率比对照组低,说明田螺能去除一定数量栅藻。由此可以推断,田螺内脏中可能含有溶藻细菌。本研究以太湖流域内野生田螺作为溶藻细菌的分离源,经富集培养后分离出溶藻效果显著的菌株;以铜绿微囊藻为指示藻,研究溶藻细菌的溶藻特性,并建立其溶藻动力学模型。
1 材料和方法
1.1 材 料
1.1.1 供试藻种 试验藻种为铜绿微囊藻(Microcystis aeruginosa,FACHB-905)购自武汉水生生物所,活化后接种于BG11培养基。
1.1.2 培养基 BG11培养基[14]用于铜绿微囊藻培养;牛肉膏蛋白胨(简称NA)培养基[15]用于细菌的分离、纯化;改良基础培养基[16]、淀粉培养基[17]用于细菌的富集培养。
1.2 方 法
1.2.1 藻细胞生长曲线的测定 铜绿微囊藻置于温度28 ℃,光照强度2 000 lx,光暗周期为12 h∶12 h条件下培养。藻液中叶绿素a含量可以作为水体生物量的表征参数[18],因此采用乙醇提取分光光度计法[19],每2 d测定其叶绿素a含量,每个样品设置3组平行,绘制生长曲线。
1.2.2 溶藻细菌的分离与纯化 选择野生生长状况良好的田螺,取适量的田螺内脏剪成小份后进行研磨[20],随后放入200 mL无菌水中,于25 ℃、150 r·min-1的振荡箱中振荡培养24 h后,静置沉淀,得到粗提液。粗提液经0.8 μm滤膜过滤,将10 mL滤液加入100 mL对数生长期的铜绿微囊藻液中共同培养,以接入10 mL无菌水的铜绿微囊藻液为对照组,7 d后将黄化明显的藻液倍比稀释,在NA培养基上平板涂布、划线分离,挑取菌落形态明显不同的菌株,编号为W1~W11。将纯化后的菌株W1~W11转接至NA液体培养基中制成菌液,菌液与藻液混合培养1周,其中溶藻效果显著、生长速率快的为实验候选菌株。
1.2.3 溶藻细菌生理生化及分子鉴定 参照《常见细菌系统鉴定手册》[21]与《伯杰氏细菌鉴定手册》[22]对细菌W10进行生理生化鉴定。菌株W10的基因组DNA提取、PCR扩增、序列测定由生工生物工程(上海)有限公司完成,将所得序列在NCBI网站上用BLAST程序与GenBank中已有的序列进行同源性比较,利用MEGA5.0软件构建系统发育树。
1.2.4 溶藻细菌生长动力学研究 以体积比1%的接种量将W10菌液接入到100 mL淀粉液体培养基中,于25 ℃、150 r·min-1的振荡箱中振荡培养,每2 h取样,使用分光光度计测其在600 nm处的吸光度,以OD600值表征其生长状况,参考王晓旭[23]的方法建立OD600与细菌浓度的线性关系,绘制生长曲线。利用生长曲线数据拟合Logistic方程[24],建立菌株的生长动力学初级模型[25-26]。Logistic方程为:
式中,y为细菌累积生长量;t为细菌生长的时间,h;k是相对最大细菌浓度,CFU·mL-1;b为细菌生长瞬时速率,h-1;a是模型参数;e是自然对数的底数。
1.2.5 溶藻细菌溶藻方式的探究 采取以下5种处理方式判断溶藻细菌W10的溶藻方式。不做任何处理的原菌液,记为T1;菌液经8 000 r·min-1、4 ℃离心15 min,将上清液与菌体细胞分离,上清液用0.22 μm针筒式滤膜过滤2次,平板检验无菌后,记为T2(即无菌上请液);将菌液离心后的菌体用淀粉培养基重悬,记为T3; 121 ℃灭菌20 min处理离心过膜后的上清液及原菌液,分别记为T4、T5,各组均设置3组平行,7 d后测定叶绿素a含量并计算溶藻率。
式中,Ct为试验组藻液的叶绿素a含量,单位:mg·m-3;C0为空白对照组藻液的叶绿素a含量,单位:mg·m-3。
1.2.6 溶藻特性 (1)培养基对溶藻效果的影响。配制NA培养基、改良基础培养基、淀粉培养基3种培养基,分别接入溶藻菌W10,以体积比10%的接种量将菌液及其无菌上清液接入至生长良好的铜绿微囊藻藻液中,对照组分别接入等量但不接菌的3种培养基,7 d后测定叶绿素a含量并计算溶藻率。
(2)不同生长时期对溶藻效果的影响。按照测定的W10菌株的生长曲线,取不同生长时期的菌液以及不同生长时期菌株分泌的无菌上清液,以体积比10%的接种量加入藻液中,每组设置3组平行,7 d后测定叶绿素a含量并计算溶藻率。
(3)不同投加量对溶藻效果的影响。为确定菌液与无菌上清液最佳的投加量,设置体积比为1∶100、1∶20、1∶10、1∶5、1∶2的试验组,每组设置3组平行,7 d后测定叶绿素a含量并计算溶藻率。
1.2.7 溶藻动力学研究 以体积比10%的投加量将W10与铜绿微囊藻混合培养,每24 h测定培养液中叶绿素a含量,探讨W10对铜绿微囊藻的溶藻动力学。溶藻过程中不仅有溶藻细菌对藻细胞的降解,还存在着实验中藻细胞自身生长[27],以及当营养物质消耗殆尽时,藻类与细菌相互竞争等问题[28]。因此溶藻细菌对藻类的降解是十分复杂的生物反应过程,本实验忽略藻细胞自身的生长及死亡,采用一级动力学模型拟合实验数据,一级动力学方程为:
S=S0e-ωt (3)
式中,S为t时培养液中叶绿素a含量,单位:mg·m-3;S0为培养液中初始叶绿素a含量,单位:mg·m-3;t为溶藻过程进行的时间,单位:d;ω为速率常数,单位:d-1;e为自然对数的底数。
2 结果与分析
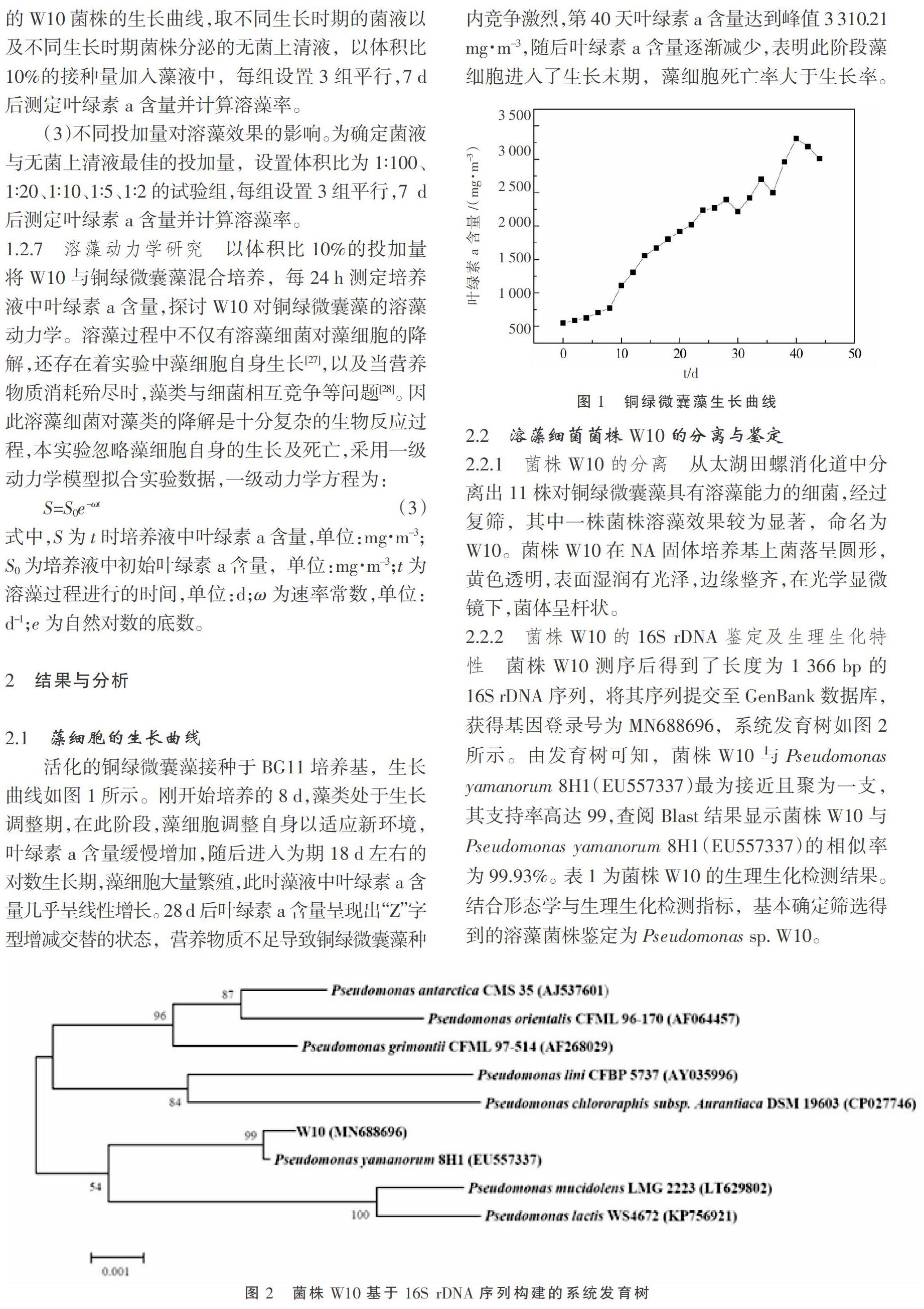
2.1 藻细胞的生长曲线
活化的铜绿微囊藻接种于BG11培养基,生长曲线如图1所示。刚开始培养的8 d,藻类处于生长调整期,在此阶段,藻细胞调整自身以适应新环境,叶绿素a含量缓慢增加,随后进入为期18 d左右的对数生长期,藻细胞大量繁殖,此时藻液中叶绿素a含量几乎呈线性增长。28 d后叶绿素a含量呈现出“Z”字型增减交替的状态,营养物质不足导致铜绿微囊藻种内竞争激烈,第40天叶绿素a含量达到峰值3 310.21 mg·m-3,随后叶绿素a含量逐渐减少,表明此阶段藻细胞进入了生长末期,藻细胞死亡率大于生长率。2.2 溶藻细菌菌株W10的分离与鉴定
2.2.1 菌株W10的分离 从太湖田螺消化道中分离出11株对铜绿微囊藻具有溶藻能力的细菌,经过复筛,其中一株菌株溶藻效果较为显著,命名为W10。菌株W10在NA固体培养基上菌落呈圆形,黄色透明,表面湿润有光泽,边缘整齐,在光学显微镜下,菌体呈杆状。
2.2.2 菌株W10的16S rDNA鉴定及生理生化特性 菌株W10测序后得到了长度为1 366 bp的16S rDNA序列,将其序列提交至GenBank数據库,获得基因登录号为MN688696,系统发育树如图2所示。由发育树可知,菌株W10与Pseudomonas yamanorum 8H1(EU557337)最为接近且聚为一支,其支持率高达99,查阅Blast结果显示菌株W10与Pseudomonas yamanorum 8H1(EU557337)的相似率为99.93%。表1为菌株W10的生理生化检测结果。结合形态学与生理生化检测指标,基本确定筛选得到的溶藻菌株鉴定为Pseudomonas sp. W10。
2.3 溶藻细菌的生长动力学分析
利用Origin9.0软件,对试验数据和Logistic方程进行非线性拟合,得到溶藻细菌生长动力学初级方程:
溶藻细菌的生长情况如图3所示。由图3可知,溶藻菌株W10在6 h内处于生长延滞期,6 h后进入对数生长期,稳定期为34~56 h,56 h后菌株W10浓度开始逐渐减少,进入衰亡期。菌株W10的生长曲线接近“S”型,Logistic模型曲线与菌株实际生长曲线拟合度较好,相关系数R2=0.990 8(P<0.001),说明Logistic方程能很好的描述在实验室环境中菌株生长过程中浓度的变化。菌株的瞬时生长速率为0.126 h-1,代时约为5.5 h,与郭惠娟等[29]研究的溶藻菌Microbacterium oleivorans最大生长速率0.05 h-1相比,菌株W10生长速度较快,代时较短,能够在短时间内达到溶藻的效果。
2.4 溶藻细菌W10溶藻方式的探究
初始铜绿微囊藻液的叶绿素a含量为2 543.99 mg·m-3,将铜绿微囊藻与5种不同处理的溶藻菌混合培养,7 d后测各组叶绿素a的含量,试验结果如图4。在实验室环境中,空白组藻液的叶绿素a含量增加了174.52 mg·m-3,证明实验过程中铜绿微囊藻自然衰亡率低于增长率。T1、T2组都表现出较好的溶藻活性,溶藻率分别为78.70%,76.81%;T3组几乎没有溶藻能力,溶藻率仅4.37%;而经过高温高压处理后的菌液、无菌上清液对铜绿微囊藻的去除率低于T1、T2组,表明菌液及无菌上清液中具有溶藻活性的物质热稳定性较低。
微生物的溶藻方式分为直接溶藻与间接溶藻,W10的无菌上清液对铜绿微囊藻的抑制作用较强,且溶藻效果没有因为将菌体去除而减弱,溶藻率与不做任何处理的T1组接近,因此可以判断对铜绿微囊藻有溶解作用的物质存在于W10的无菌上清液中,菌株W10是通过分泌某些胞外活性物质溶解藻类而非与藻类竞争营养物质,胞外活性物质热稳定性较差,可能为蛋白质类物质,属于间接溶藻。
2.5 溶藻细菌W10溶藻特性
2.5.1 培养基对溶藻效果的影响 由图5可知,在叶绿素a含量为1 024.63 mg·m-3的初始条件下,W10在NA培养基中培养,溶藻率表现为菌液(80.46%)>无菌上清液(73.66%)>对照(24.88%),三者之间差异均达显著水平;W10在淀粉培养基和改良基础培养基中培养,其菌液及无菌上清液的溶藻率差异不显著,二者均显著高于对照组(P<0.05)。3种培养基中,W10菌液及无菌上清液在NA和淀粉培养基中的溶藻效果差异不显著(P>0.05),二者均显著优于改良基础培养基(P<0.05),但由于NA培养基为天然培养基[30],其成分复杂,会对铜绿微囊藻的生长产生消极影响,故其对照组的溶藻率显著高于淀粉和改良基础培养基对照组(P<0.05),因此后续使用成分简单明了的淀粉培养基,有利于排除培养基对试验结果的干扰。
2.5.2 不同生长时期对溶藻效果的影响 试验初始叶绿素a含量为1 109.58 mg·m-3,经过7 d混合培养后,各试验组的叶绿素a含量均低于初始值。由图6可知,4个生长时期的菌株W10及其无菌上清液对铜绿微囊藻的生长均有抑制作用,溶藻率表现为除稳定期与衰亡期差异不显著(P>0.05)外,其他各时期之间差异均达显著水平(P<0.05),且同一生长时期W10菌液与其无菌上清液之间的溶藻率差异均不显著(P>0.05)。延滞期的菌液溶藻效果最低,原因可能是细菌在延滞期还未完全适应新环境,生长较为缓慢,并且分泌的胞外溶藻活性物质少。随着培养时间的延长,菌株W10产生的胞外溶藻活性物质逐渐积累,进入对数生长期和稳定期后,W10活性较高,溶菌效果均显著提升。而衰亡期的菌液中含有大量死亡的菌体,添加等体积的菌液与铜绿微囊藻共同培养,菌液中的胞外溶藻活性物质相对稳定期时减少,导致溶藻率降低,这也从侧面证明了菌株W10的作用方式为间接溶藻。司晓光等[31]也证明溶藻效果与菌株分泌的溶藻物质数量有关,溶藻物质越多,则溶藻效果越好。衰亡期W10及其无菌上清液溶藻活性与稳定期相比不再增加,说明稳定期的无菌上清液的溶藻效果几乎达到极限,细菌进入衰亡期后,几乎不再继续分泌胞外溶藻活性物质。
2.5.3 不同投加量对溶藻效果的影响 如图7所示,初始铜绿微囊藻叶绿素a含量为1 124.07 mg·m-3,随着投加量的增加,W10菌液溶藻率呈先升高后降低的趋势,当菌液与藻液的体积比为1∶10时达到最大值,且各投加量处理间差异均达显著水平(P<0.05);无菌上清液溶藻率呈上升趋势,至体积比1∶2时达到最大值,各投加量处理之间除1∶10与1∶5差异不显著(P>0.05)外其他处理间差异均达显著水平(P<0.05);在菌藻比1∶100、1∶20和1∶10处理,同一投加量处理W10菌液与其无菌上清液溶藻率差异不显著,而在菌藻比1∶5和1∶2处理,W10菌液溶藻率显著低于无菌上清液。无论是W10菌液还是无菌上清液,均在体积比1∶100处理溶藻率最低,几乎没有溶藻现象,此时细菌或无菌上清液添加量少,藻细胞占主导地位,藻细胞的胞外分泌物可保护藻细胞不被溶解。溶藻过程进行7 d后,当菌藻比超过1∶10时继续增加菌液的投加量,细菌对铜绿微囊藻的叶绿素a的去除效果不增反而减少,这可能是因为随着时间的延长,锥形瓶环境中营养物质消耗殆尽,大量细菌因缺乏营养物质衰亡,同时细菌种內竞争增大,最终导致溶藻率下降。
2.6 W10对铜绿微囊藻的降解动力学分析
菌株W10降解铜绿微囊藻的一级动力学方程为:
S=1 079.003 9e-0.235 7t(5)
溶藻细菌W10溶解铜绿微囊藻过程中叶绿素a含量变化与动力学模型拟合的结果如图8所示。溶藻菌株与藻液混合培养的第1天,培养液中藻细胞浓度较高,藻类占主导地位,溶藻反应还未完全开始,此时叶绿素a含量下降缓慢。随着溶藻时间的增加,W10的胞外溶藻活性物质逐渐累积,同时利用已裂解的藻细胞来满足自身的生长繁殖,培养液的叶绿素a含量不断下降。表2为菌株W10实际溶藻过程中的叶绿素a含量与一级动力学模型预测的叶绿素a含量之间的比较,实际结果与预测结果的相对误差范围为0.568 3%~6.825 0%。此外,一级动力学模型曲线与菌株W10实际溶藻过程叶绿素a含量变化拟合度较好,相关系数R2=0.988 2(P<0.001)。由此我们可以推断,W10菌株在溶藻过程中,对铜绿微囊藻的溶解遵循一级反应动力学。
3 结 论
(1)从太湖流域内野生田螺内脏中分离出溶藻菌株W10,经鉴定W10与假单胞菌属的16S rDNA序列同源性达99%,归属于假单胞菌属(Pseudomonas sp.)。溶藻菌株W10对铜绿微囊藻的溶解方式为间接溶藻,且胞外溶藻活性物质热稳定性较差,可能为蛋白质类物质。以铜绿微囊藻为底物时,溶藻菌株W10的溶解作用遵循一级反应动力学,其溶藻动力学方程为S=1 079.003 9e-0.235 7t。
(2)溶藻菌株W10及其无菌上清液在不同培养基、生长时期及投加量等条件下均具有一定溶藻效果。其中,NA和淀粉培养基显著优于改良基础培养基;同一生长时期的菌液与其无菌上清液溶藻效果无显著差异(P>0.05),二者均表现为:稳定期与衰亡期最高,二者差异不显著(P>0.05),然后依次是对数期和延滞期;当菌液与藻液体积比为1∶10时,W10菌液溶藻率最高为80.05%,而无菌上清液的溶藻效果则与投加量呈正相关,以100 mL铜绿微囊藻加入50 mL无菌上清液处理溶藻率最高,可达92.15%。
参考文献 :
[1]SU J F, MA M, WEI L, et al. Algicidal and denitrification characterization of Acinetobacter sp. J25 against Microcystis aeruginosa and microbial community in eutrophic landscape water[J]. Marine pollution bulletin, 2016,107(1):233-239.
[2]MIZUTA D D, WIKFORS G H. Can offshore HABs hinder the development of offshore mussel aquaculture in the northeast United States[J]. Ocean and coastal management, 2020, 183.
[3]ZHENG N N, DING N, GAO P K, et al. Diverse algicidal bacteria associated with harmful bloom-forming Karenia mikimotoi in estuarine soil and seawater[J]. The science of the total environment, 2018, 631-632.
[4]LI Y, ZHU H, ZHANG H, et al. Toxicity of algicidal extracts from Mangrovimonas yunxiaonensis strain LY01 on a HAB causing Alexandrium tamarense[J]. Journal of hazardous materials, 2014, 278:372-381.
[5]SU J F, SHAO S C, MA F, et al. Bacteriological control by Raoultella sp. R11 on growth and toxins production of Microcystis aeruginosa[J]. Chemical engineering journal, 2016, 293:139-150.
[6]YANG Y F, HU X J, ZHANG J, et al. Community level physiological study of algicidal bacteria in the phycospheres of Skeletonema costatum and Scrippsiella trochoidea[J]. Harmful algae, 2013, 28:88-96.
[7]SU J, YU Z, TIAN Y, et al. Biological activity of a red-tide alga—A. tamarense under co-cultured condition with bacteria[J]. Journal of environmental sciences, 2005, 17(6): 1047.
[8]WANG X, LI Z J, SU J Q, et al. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere[J]. Biological control, 2009, 52(2):123-130.
[9]ROTH P B, TWINER M J, MIKULSKI C M, et al. Comparative analysis of two algicidal bacteria active against the red tide dinoflagellate Karenia brevis[J]. Harmful algae, 2008, 7(5):682-691.
[10]史荣君, 黄洪辉, 齐占会, 等. 海洋细菌N3对几种赤潮藻的溶藻效应[J]. 环境科学, 2013, 34(5): 1922-1929.
[11]CHUAN T, XIANGLONG L, JING T, et al. Isolation, identification and characterization of an algicidal bacterium from Lake Taihu and preliminary studies on its algicidal compounds[J]. Journal of environmental sciences (China), 2012, 24(10):1823-1831.
[12]张文艺, 李仁霞, 陈雪珍, 等, 从泥鳅体内筛选的溶藻菌及利用其去除铜绿微囊藻的方法[P]. CN103484403A, 2014-01-01.
[13]何健, 谷孝鸿, 黄诚. 短钝溞(Daphnia obtusa Kurz)在“藻-螺-溞”系统中的反馈养殖与污水净化[J]. 湖泊科学, 1999(4): 328-332.
[14]朱晓漫, 罗玉双, 李娜, 等. 一株溶藻细菌的分离、筛选与分子鉴定[J]. 湖南文理学院学报(自然科学版), 2017, 29(1): 28-34.
[15]崔亚青, 雍晓雨, 张风革, 等. 一株溶藻细菌的分离鉴定及溶藻效果[J]. 应用与环境生物学报, 2012, 18(5): 752-760.
[16]许珂, 吴刚, 余文平, 等. 溶藻细菌W5培养条件优化及发酵培养[J]. 环境科学与技术, 2009, 32(2): 12-15.
[17]林敏, 潘伟斌, 张太平, 等. 三株溶藻细菌溶藻活性代谢产物的初步研究[J]. 生态环境, 2007(2): 358-362.
[18]刘波, 崔莉凤, 刘载文. 北京市城区地表水体叶绿素a与藻密度相关性研究[J]. 环境科学与技术, 2008(8): 29-33.
[19]陈建勋, 王晓峰. 植物生理学实验指导(第二版)[M]. 广州:华南理工大学出版社, 2006.
[20]董小娜, 陈泽慧, 毛林强, 等. 太湖土著激浪鱼内脏中溶藻菌R1的筛选及其特性研究[J]. 工业安全与环保, 2018, 44(8): 69-72.
[21]东秀珠,蔡妙英. 常见细菌鉴定手册[M].北京:科学出版社, 2001.
[22]布坎南RE,吉本斯NE. 伯杰氏细菌鉴定手册(第8版)[M].北京:科学出版社. 1984.
[23]王晓旭, 徐峰, 沈丽萍, 等. 分光光度法测定猪链球菌菌悬液浓度[J].上海畜牧兽医通讯, 2017(5): 40-41.
[24]VOGELS M, ZOECKLER R, STASIW D M, et al. P. F. Verhulst's “notice sur la loi que la populations suit dans son accroissement” from correspondence mathematique et physique. Ghent, vol. X, 1838[J]. Journal of biological physics, 1975, 3(4): 183-192.
[25]孫鹏, 孙源. 3种方法对中肋骨条藻Logistic生长模型拟合的比较研究[J]. 江苏农业科学, 2019, 47(2): 229-232.
[26]王福林, 王吉权. 生长曲线参数估计的一种新方法—优化回归组合法[J]. 生物数学学报, 2007(3): 533-538.
[27]CHOI H J, KIM B H, KIM J D, et al. Streptomyces neyagawaensis as a control for the hazardous biomass of Microcystis aeruginosa (Cyanobacteria) in eutrophic freshwaters[J]. Biological control, 2005, 33(3):335-343.
[28]李小彩. 一株红球菌的溶藻特性及应用研究[D].济南:山东大学,2007.
[29]郭惠娟, 张伟, 张小梅, 等. 溶藻细菌Microbacterium oleivoran的溶藻进程与叶绿素降解动力学[J]. 环境化学, 2019, 38(6): 1274-1281.
[30]吕伟英, 赵以军, 周瑞, 等. 一种快速检测分离溶藻细菌方法的初探[J]. 微生物学通报, 2007(1): 119-122.
[31]司晓光, 张晓青, 郝建安, 等. 芽孢杆菌dhs-330-021对链状亚历山大藻的溶藻机理研究[J]. 生物技术通讯, 2017, 28(4): 485-489.
