miR-1182在卵巢癌组织中的表达及对肿瘤细胞增殖与转移的影响
2020-10-13李维张卫平马静
李维 张卫平 马静
天津中医药大学第一附属医院检验科300193
0 引 言
卵巢癌是常见的妇科恶性肿瘤,其早期隐匿性强,不易被发现,导致病死率高。开展卵巢癌的发生、发展及可能机制的研究,对其诊断与治疗有重要意义,已成为妇科恶性肿瘤领域的研究热点[1]。miRNAs(micro RNA)是一类约21 nt的内源性核苷酸小RNA,可作为转录后调节因子的靶向调节靶基因,对机体起调控的作用。miRNAs在肿瘤的生理和病理过程中起重要的调节作用[2]。关于miRNAs相关靶点在卵巢癌转移治疗中的应用研究是近年来的医学研究热点[3-4]。miRNAs在卵巢癌的多药耐药发生过程中,以及在克服卵巢癌化疗药物耐药性过程中起关键的调控作用[5]。miRNAs-1182是miRNAs调控网络的重要组成部分,在胃癌、肠癌、肾癌等恶性肿瘤中,及在细胞增殖、分化、转移等方面起重要的调节作用[6-8]。但是,关于miRNAs在卵巢癌中的研究并不多见。本研究中,分析miRNAs-1182在卵巢癌组织中的表达,探讨其对癌组织增殖与转移的影响,及可能的调控机制。
1 资料与方法
1.1 临床资料
选择2016~2019年在天津中医药大学第一附属医院就诊的81例卵巢癌患者,年龄范围36~67岁,年龄(51.3±9.8)岁,其中年龄<50岁患者 46例,年龄≥50岁患者35例。所有患者均经病理确诊为卵巢癌,且均为初治,既往未进行放疗及化疗。
收集患者的卵巢癌组织及对应的癌旁组织,并进行病理分析。肿瘤分期采用2013年国际妇产科联合会的分期标准。81例卵巢癌患者中:Ⅰ期10例、Ⅱ期18例,Ⅲ期32例,IV期21例;高分化11例、中分化22例、低分化48例;上皮性卵巢癌41例、性索间质卵巢癌22例、生殖细胞卵巢癌18例。
另选择同时期在天津中医药大学第一附属医院行卵巢切除术的52例非卵巢癌患者,年龄范围39~66 岁,年龄(53.3±11.4)岁。
1.2 主要材料与仪器
TrizolRNA提取试剂盒、miRNA逆转录试剂盒、实时荧光定量聚合酶链式反应(polymerase chain reaction,PCR)试剂盒 SYBR GREEN Premix Ex Taq(日本宝生物工程株式会社)、PCR扩增引物序列[生工生物工程(上海)股份有限公司],人端粒酶逆转录酶(human telomerase reverse transcriptase,hTERT)单克隆抗体(美国 ABCAM 公司),β-肌动蛋白(βactin)多克隆抗体、辣根过氧化物酶标记的山羊抗兔IgG抗体(美国Santa Cruz公司),Transwell小室(美国Corning公司)
7900HT Fast实时荧光定量PCR仪(美国ABI Life Technologies公司),Thermo MK3酶标仪、CO2细胞培养箱(美国Thermo Scientific公司),IX71倒置显微镜(日本奥林巴斯公司)。
1.3 方法
1.3.1 RT-PCR检测卵巢组织中miR-1182的表达水平
采用Trizol法,按照RNA提取试剂盒说明书提取组织的总RNA,采用miRNAs逆转录试剂盒,以总RNA为模板进行逆转录反应成cDNA。U6引物:5′-GACACGCA-AATTCGTG-3′,5′-GTGCAGGGTCCG-AGGT-3′。miR-1182 引物:5′-ACCTTCCTCAGGACCCTGGTCCGAGGT-3′,5′-CAACATCTACAAGATCCTCCTGCTGCAGG-3′。PCR 反应体系为:cDNA 1.5 μl,miR-1182 或 U6 引 物 1 μl,SYBR GREEN premix Ex taq 10 μl,H2O 7.5 μl。反应条件为:预变性95℃、30 s,变性 95℃、5 s,退火延伸 60℃、30 s,共40个循环;72℃延伸10 min。使用7900HT Fast型实时荧光定量PCR仪,根据2-ΔCt法计算miR-1182的表达量。所有实验均重复3次,取3次结果的均值±标准差为实验结果。
1.3.2 miR-1182病毒转染人卵巢癌细胞株SKOV-3
人卵巢癌细胞株SKOV-3用DMEM培养基(10%胎牛血清)培养,以5×105密度接种于6孔板。正常对照组按照常规培养,病毒转染组采用miR-1182 mimics(1.0 μg/ml)转染细胞,另设立空白病毒组,6 h后换常规培养基。转染48 h后收集各组细胞,按照RNA提取试剂盒说明书提取细胞总RNA,RT-PCR方法见1.3.1节。
1.3.3 Western Blot检测SKOV-3细胞hTERT蛋白的表达情况
转染48 h后收集各组SKOV-3细胞,SDS裂解液提取细胞总蛋白,蛋白定量后每孔蛋白上样量为30 μg,SDS-聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,PAGE)后湿转法转膜,5%脱脂奶粉封闭1 h,1∶500 hTERT单克隆抗体4℃下孵育过夜,1∶1000 二抗室温孵育 1 h,以 1∶1000 的 β-actin作为内参,进行化学发光法显影。
1.3.4 噻唑蓝法检测SKOV-3细胞增殖情况
将SKOV-3细胞以1×104密度接种于96板,按照1.3.2中的分组与病毒转染方法,继续培养24、48、72h后,以每孔加入5mg/ml的噻唑蓝溶液20μl,继续培养4h后弃上清液,每孔加二甲基亚砜150μl,振荡溶解15 min,使用酶标仪检测各孔于450 nm处的吸光度(A450)值。
1.3.5 细胞侵袭实验检测SKOV-3细胞穿膜情况
转染48 h后收集细胞,以1×105密度接种于Transwell小室内,将小室至于含有常规培养基的48孔板内,培养24 h后取出小室,刮去室内细胞,用4%多聚甲醛固定,0.1%结晶紫染色,双蒸水清洗,显微镜下选取6个视野进行细胞计数,算平均值。每组设3个复孔。
1.4 统计学方法
采用SPSS22.0软件进行统计学分析,计量资料以均值±标准差(Mean±SD)表示,两组间数据比较采用t检验。以P<0.05为差异具有统计学意义。
2 结 果
2.1 miR-1182的表达情况
miR-1182在正常卵巢组织、卵巢癌组织和癌旁组织中的表达水平如图1所示。结果显示,miR-1182在正常卵巢组织(3.06±0.29)与癌旁组织(2.92±0.34)中的表达水平的差异无统计学意义(P>0.05);miR-1182在卵巢癌组织中的表达水平(0.66±0.19)明显低于正常卵巢组织(3.06±0.29)与癌旁组织(2.92±0.34)(均 P<0.05)。
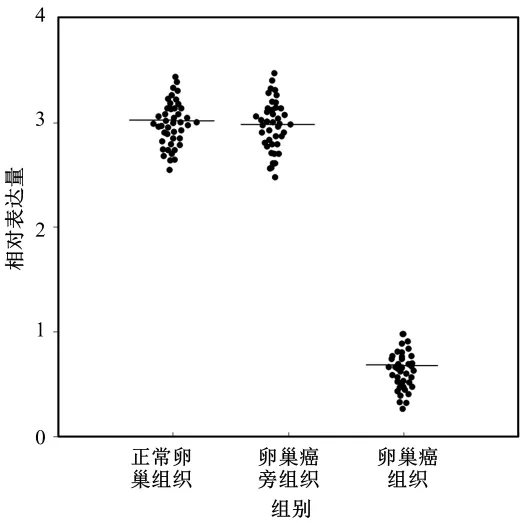
图1 miR-1182在组织中的表达水平
2.2 卵巢癌组织中的miR-1182的表达水平与临床病理特征的关系
结果显示,对于卵巢癌组织中miR-1182的表达水平,不同肿瘤分期、组织分化程度患者间的差异有统计学意义(均P<0.05),而患病年龄与肿瘤类型的差异无统计学意义(均P>0.05)。(表1)
2.3 卵巢癌细胞miR-1182及hTERT蛋白的表达情况
在SKOV-3卵巢癌细胞转染miR-1182 mimics病毒后,转染组的miR-1182表达水平较空白病毒组和对照组明显升高(均P<0.05),而对照组和空白病毒组的miR-1182表达水平差异无统计学意义(P>0.05)。(图 2)
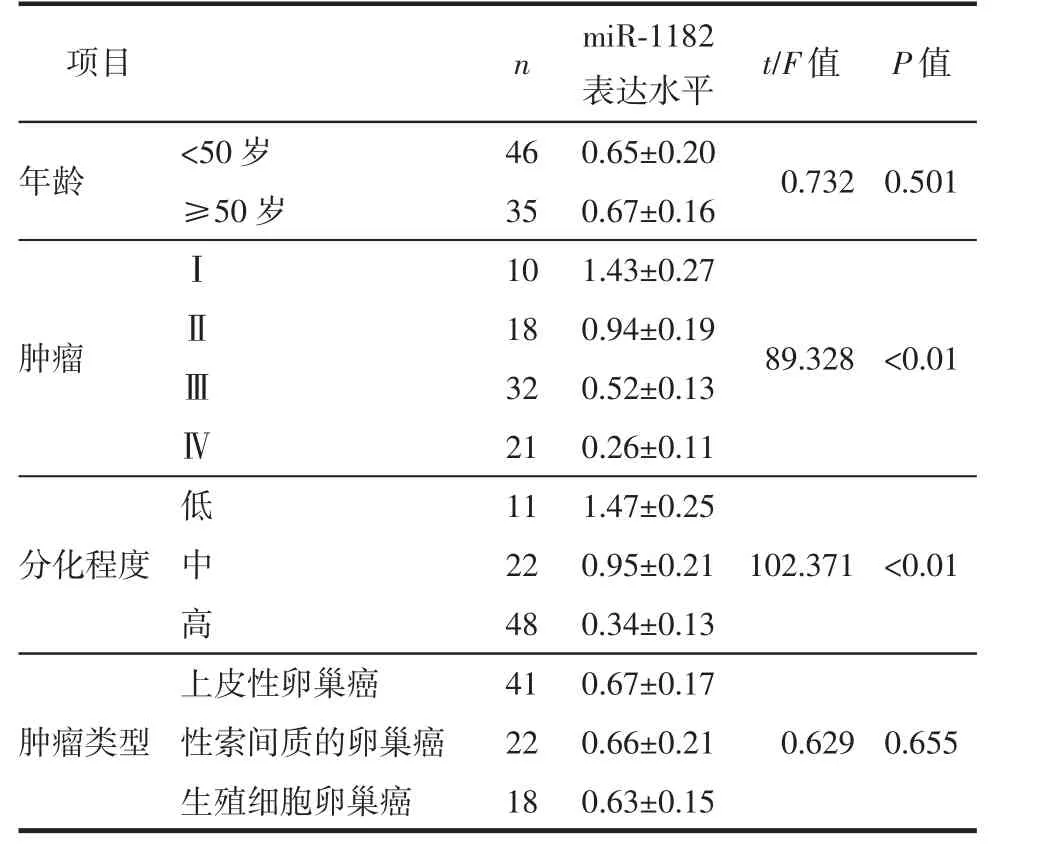
表1 卵巢癌组织中的miR-1182的表达水平与临床病理特征间的关系
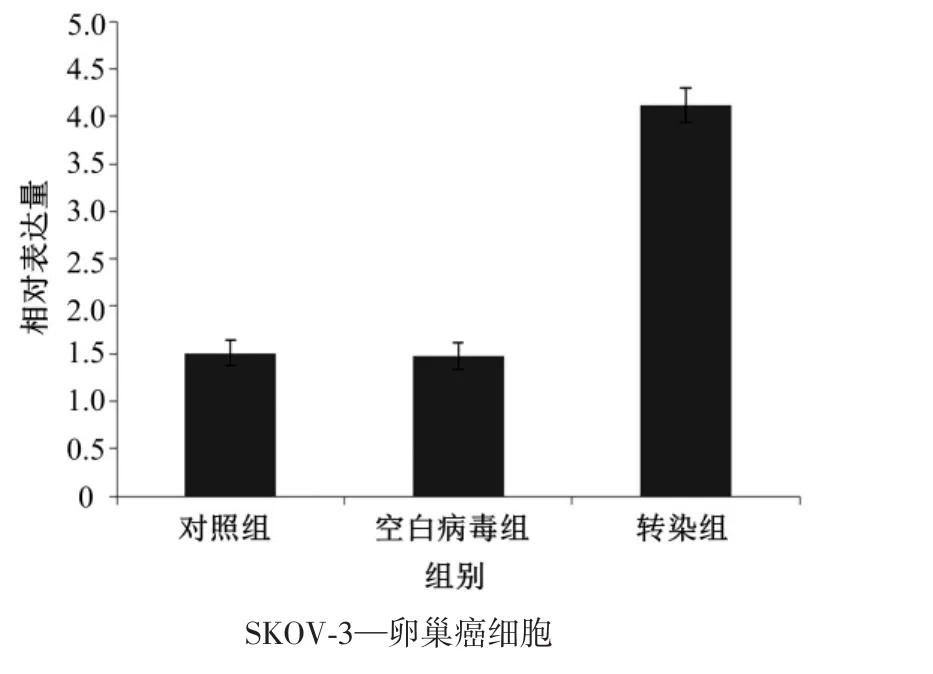
图2 各组SKOV-3卵巢癌细胞转染miR-1182 mimics病毒后的miR-1182表达水平
Western Blot结果显示,在SKOV-3卵巢癌细胞转染miR-1182 mimics病毒后,转染组hTERT蛋白表达水平较空白病毒组和对照组明显降低(均P<0.05),而对照组和空白病毒组的hTERT蛋白表达水平差异无统计学意义(P>0.05)。(图3)

图3 各组SKOV-3卵巢癌细胞转染miR-1182 mimics病毒后的hTERT蛋白表达
2.4 miR-1182对卵巢癌细胞增殖与侵袭的影响
细胞增殖实验结果显示,对照组和空白病毒组在各时间点A450值的差异无统计学意义(均P>0.05),转染组在培养24、48、72 h后,A450值明显低于对照组和空白病毒组(均P<0.05)。
细胞培养48 h后,细胞穿膜数量结果显示,对照组和空白病毒组穿膜细胞数的差异无统计学意义(P>0.05),而转染组穿膜细胞数明显低于对照组和空白病毒组(均 P<0.05)。(表 2)。

表2 各组SKOV-3卵巢癌细胞转染miR-1182 mimics后细胞的增殖与侵袭情况
3 讨论与结论
miRNAs表达异常发生在很多常见的人类肿瘤组织中。由于miRNAs具有靶向基因的调控作用,对细胞生长、分化和凋亡的调控起重要作用,其与肿瘤的发生、发展密不可分[9-10]。因此,部分miRNAs在肿瘤的诊断与治疗中,存在潜在的应用价值[11-12]。
miRNAs-1182是miRNAs调控网络的重要组成部分。多项研究结果证实,miRNAs-1182对许多肿瘤发挥重要的调节作用[13]。circABCC4通过miR-1182促进FOXP4的表达并促进前列腺癌的恶性发展,circABCC4/miR-1182/FOXP4调节环可能是前列腺癌介入治疗的一个潜在的理想靶点[14]。LINC00339与miR-1182相互作用促进SKA1的表达,表明LINC00339/miR-1182/SKA1在肝癌进展中起重要作用[15]。卵巢癌中高表达circWHSC1,促进了肿瘤的发生,而通过调节miR-145和miR-1182其外显子,作用于腹膜间皮,可诱导肿瘤转移[16]。本研究中,检测了正常卵巢组织、卵巢癌组织和癌旁组织中的miR-1182表达水平,发现miR-1182在卵巢癌组织中表达明显降低,这与Hou等[17]的研究结果一致。此外发现:对于卵巢癌组织中的miR-1182表达水平,不同肿瘤分期、分化程度的患者间有明显差异;随临床分期的进展及组织分化程度增加,miR-1182的表达水平逐渐降低。提示miR-1182可能在卵巢癌的发生发展中起重要作用。
本研究中,通过体外实验进一步研究miR-1182在卵巢癌中的作用机制。在用miR-1182 mimics病毒转染SKOV-3卵巢癌细胞后,检测细胞中miR-1182和hTERT的表达情况。结果显示,转染miR-1182病毒后,miR-1182的表达明显升高,hTERT的表达明显降低。提示在卵巢癌细胞中,miR-1182对hTERT蛋白有负调节作用。有研究结果表明,miR-1182通过与hTERT的开放读码框(open reading frame,ORF)结合,在转录后抑制 hTERT 的表达[4]。有研究者通过荧光素酶检测证实,hTERT是miR-1182的直接靶点,miR-1182在卵巢癌中的抑制作用是通过miR-1182/hTERT轴实现的[17]。
本研究中进一步发现,高表达miR-1182的卵巢癌细胞在培养24、48和72 h后,细胞增殖受到明显的抑制,且高表达miR-1182的卵巢癌细胞的穿膜数量明显减少。该结果提示,miR-1182的表达可明显抑制卵巢癌细胞的侵袭与浸润能力。有研究结果表明,miR-1182的过度表达能显著抑制膀胱癌细胞增殖、集落形成和侵袭,miR-1182通过结合其3'UTR直接靶向hTERT[18]。hTERT是端粒酶催化亚基,具有调控端粒酶活性和促进端粒延长的作用。端粒酶是细胞癌变过程中的重要调控因子,其无直接致癌作用,但是它具有促进肿瘤细胞周期,延长肿瘤细胞生存时间,进而促进肿瘤形成的作用。
综上,miR-1182在卵巢癌组织中低表达,而过表达miR-1182可抑制卵巢癌的增殖与侵袭,其通过靶向调节hTERT表达实现调控作用。
利益冲突 所有作者均声明不存在利益冲突
