墨西哥湾扇贝和扇贝“渤海红”及其杂交子代的遗传分析*
2020-09-27姚高友吴羽媛苏晓盈刘付少梅张方家熙陈楠生王春德刘志刚
姚高友 谭 杰 吴羽媛 苏晓盈 刘付少梅张 元 方家熙 陈楠生 王春德 刘志刚
墨西哥湾扇贝和扇贝“渤海红”及其杂交子代的遗传分析*
姚高友1,3谭 杰1,3吴羽媛1苏晓盈1刘付少梅2张 元1,3方家熙4陈楠生5王春德6刘志刚1,3①
(1. 广东海洋大学水产学院 湛江 524088;2. 湛江银浪海洋生物技术有限公司 湛江 524022;3. 广东省南海经济无脊椎动物健康养殖工程研究中心 湛江 524088;4. 香港理工大学应用生物及化学科技学系 香港 999077;5. 西蒙·弗雷泽大学理学院分子生物和生物化学系 加拿大不列颠哥伦比亚省本那比市 V5A 1S6;6. 青岛农业大学海洋与工程学院 青岛 266109)
利用SSR (Simple Sequence Repeats)分子标记技术,对扇贝“渤海红”、墨西哥湾扇贝()及其杂交子代3个群体共90个个体的遗传多样性进行分析。结果显示,8个SSR位点共扩增出67个等位基因,各位点等位基因数范围为4~14个,平均等位基因数为8.5个。扇贝“渤海红”有效等位基因数(N)、平均观测杂合度(H)和多态信息含量(PIC)最高,分别为2.3947、0.504和0.462;3个群体的遗传分化指数(F)、基因流(N)和固定系数(F)分别为0.1398、1.5387和0.3698。对亲代与杂交子代间的遗传分化分析表明,杂交子代与扇贝“渤海红”的遗传距离最小(0.1188),遗传相似度最大(0.888),杂交子代的遗传结构更偏向亲本扇贝“渤海红”。研究结果可为扇贝“渤海红”和墨西哥湾扇贝群体种质资源评估和杂交新品种的选育提供理论参考。
墨西哥湾扇贝;扇贝“渤海红”;杂交;遗传多样性
扇贝“渤海红”为紫扇贝()与海湾扇贝北部亚种()杂交,经 6代选育而成,于2015年通过新品种审定。与传统海湾扇贝相比,扇贝“渤海红”生长上表现出极显著的杂种优势,产生了巨大的社会及经济效益(丰玮等, 2012; 李建立等, 2012; 南乐红等, 2012; Wang, 2017),但扇贝“渤海红”高温耐受性差,96 h半致死温度为29℃ (Wang, 2017),不利于其在全国推广养殖。墨西哥湾扇贝()又称海湾扇贝南方亚种,原产于美国大西洋沿岸,其生长快、鲜肉柱率高、高温耐受性强,稚贝在32℃海水中存活率达76%(刘志刚等, 2007a),1991年引进中国后在南方北部湾成功推广,成为北部湾渔业支柱产业之一(Abbott, 1974; 刘志刚等, 2007b)。但由于最初引进群体小且经过20多年小群体繁育,墨西哥湾扇贝已出现较为严重的种质退化问题(彭张明等, 2014),影响产业健康可持续发展。
杂交育种是贝类遗传改良的重要方法,能够有效提高子代生长及抗逆性状。为解决墨西哥湾扇贝种质退化问题,国内学者进行了华贵栉孔扇贝()×墨西哥湾扇贝(潘奕达, 2016; 潘奕达等, 2017)、紫扇贝×墨西哥湾扇贝(南乐红等, 2012)种间杂交与墨西哥湾扇贝×海湾扇贝等北部亚种(Zhang,2007; Zheng, 2011)等种内杂交。其杂交子代虽然在生长上表现出一定的杂种优势,但未能完全适应我国南方海域环境,种质退化问题仍未得到有效解决。为改善墨西哥湾扇贝种质退化和扇贝“渤海红”高温敏感性问题,课题组进行了墨西哥湾扇贝与扇贝“渤海红”的杂交育种研究,并成功获得杂交子代。杂交子代在生长上表现出明显的杂种优势,并完全适应我国南方北部湾海域环境,具有良好的推广养殖前景。
种群结构的变异大小可以评判种群抵御外界环境压力及进化的潜力,即遗传多样性较高的种群对环境恶化耐受力较强,反之,则可能出现衰退(Cronquist, 1991; 钱迎倩等, 1994)。遗传多样性评估对贝类健康养殖指导及适应性进化潜力评价具有重要意义。 刘保忠(2003)和Wang等(2011)发现,我国海湾扇贝养殖群体与原引种地区海湾扇贝群体相比,等位基因数量降低了20%,遗传多样性存在瓶颈效应。谭杰等(2018)研究发现,海湾扇贝北部亚种“中科红”经过多年的定向选育,仍具有较高的遗传多样性,还具有较高的选育潜力。目前,关于扇贝“渤海红”及墨西哥湾扇贝及其杂交子代遗传多样性研究未见报道。SSR分子标记技术作为重要的DNA分子标记技术之一,在皱纹盘鲍()(聂鸿涛等, 2013)、栉孔扇贝()(张秀英等, 2012)、虾夷扇贝()(韩斐斐等, 2012; 张广明等, 2018)、栉江珧()(李东明等, 2017)和马氏珠母贝()(赵晓霞等, 2010)等多种重要海水贝类遗传多样性分析中得到广泛应用。荧光标记毛细管电泳检测技术具有自动化、精度好、高通量等优点,已在长牡蛎()(郭香等, 2018)、欧洲牡蛎()(Wang, 2010)等重要贝类品种中得到应用。本研究利用8对SSR引物,采用毛细管电泳技术,对扇贝“渤海红”、墨西哥湾扇贝及其杂交子代的遗传多样性进行分析,以期为扇贝“渤海红”和墨西哥湾扇贝的群体遗传评估和杂交新品种的选育提供一定的理论指导。
1 材料与方法
1.1 实验材料
墨西哥湾扇贝群体为北部湾普通养殖群体,由湛江银浪生物技术有限公司提供。扇贝“渤海红”群体由青岛农业大学王春德教授提供。杂交贝为扇贝“渤海红”×墨西哥湾扇贝正反交子代(各1/2)中的橙色壳色群体。每个群体随机选取30个个体,取闭壳肌于无水乙醇中,放入–20℃保存备用。
1.2 基因组DNA提取与检测
采用全式金公司海洋生物基因组DNA提取试剂盒(Marine Animal Genomic DNA Kit)提取各群体基因组DNA。以1%琼脂糖凝胶电泳检测DNA质量。
1.3 微卫星引物
从本课题组开发的墨西哥湾扇贝SSR引物中,挑选多态性好、等位基因数较多的8对引物用于遗传分析,引物信息见表1,荧光SSR引物由天一辉远公司合成。
1.4 PCR扩增程序及反应
PCR反应采用全式金公司Super mix试剂盒,PCR反应体系为10 μl,其中,模板0.5 μl,引物对各0.5 μl,2×PCR Master Mix 5 μl,无菌水3 μl。PCR扩增反应程序:95℃预变性5 min,10个循环,每个循环降低1℃(95℃变性30 s,62℃~52℃退火30 s,72℃延伸30 s),25个循环(95℃变性、52℃退火、72℃延伸),72℃延伸10 min,4℃,保存。扩增产物采用ABI3730XL测序仪进行片段长度分析。
1.5 数据分析
利用POPGENE 1.32软件计算群体的等位基因数(N)、有效等位基因数(N)、期望杂合度(H)、观测杂合度(H)、遗传相似系数及遗传距离等。使用PIC-CALC软件计算多态信息含量。
2 结果与分析
2.1 微卫星位点的多态性
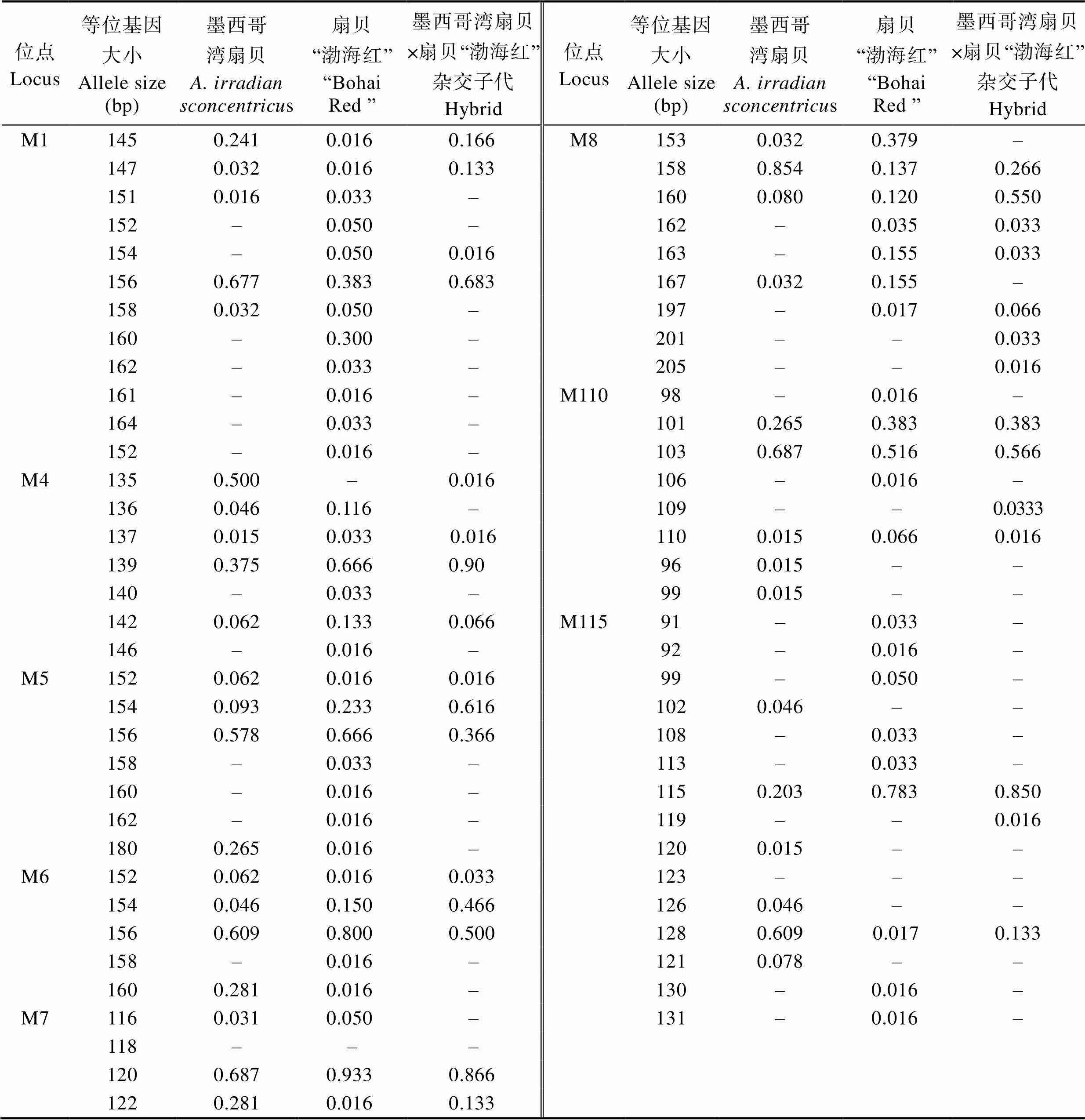
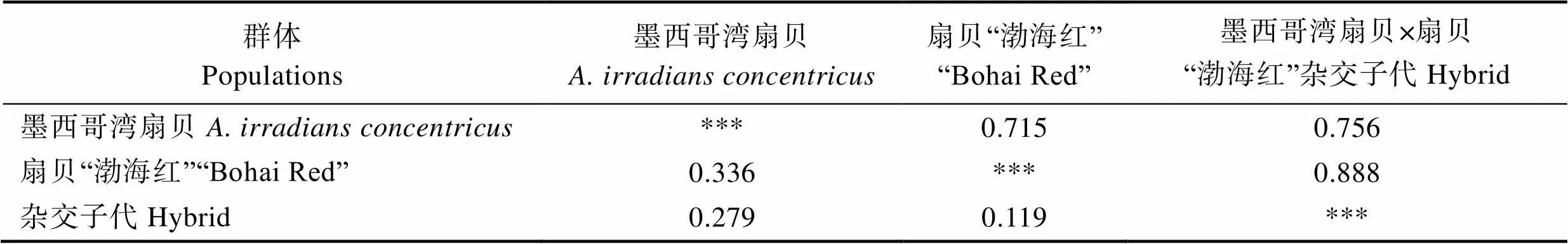
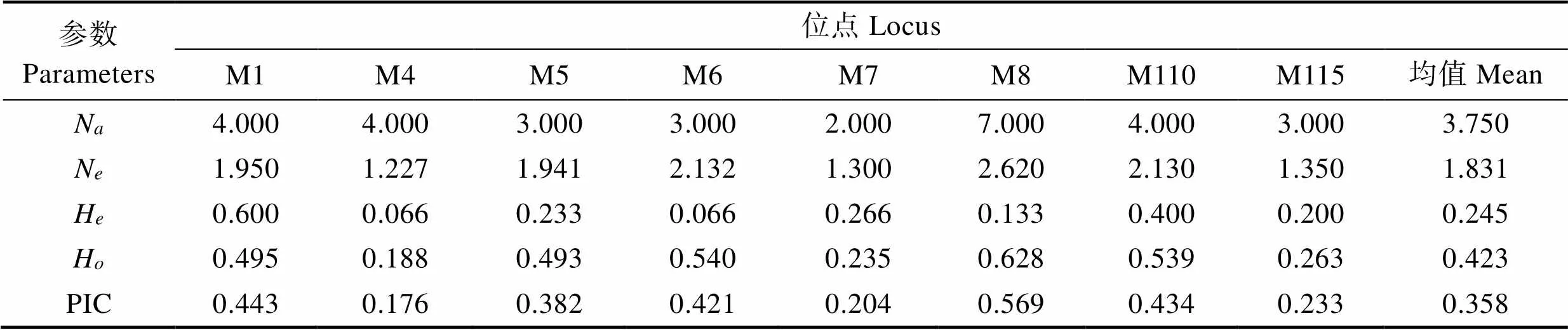
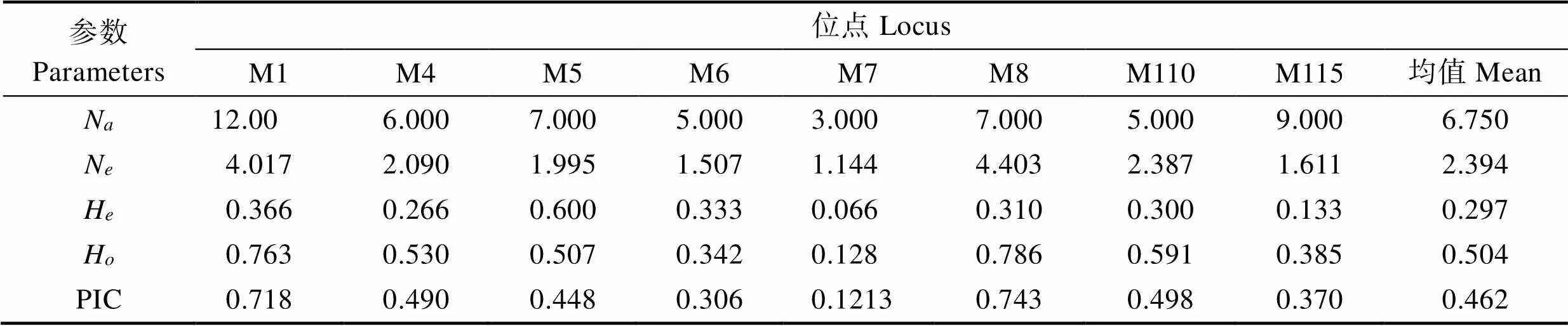
8个简单重复序列标记(Simple Sequence Repeats, SSR)位点均能在扇贝“渤海红”、墨西哥湾扇贝及其杂交子代中有效扩增。表2为8个位点的遗传多样性信息。8个微卫星位点共得到68个等位基因,其中,位点M1的等位基因最多(12个),位点M7等位基因最少(4个),各微卫星位点的期望杂合度(H)范围为0.2973~0.6265,观测杂合度(H)范围为0.1556~ 0.5385,平均期望杂合度为0.5491,平均观测杂合度为0.2960,多态信息含量(PIC)介于0.2657~0.6255之间,平均多态信息含量为0.49836,说明位点具有较高的遗传多样性,其中,高度多态位点5个(PIC>0.5),中度多态位点3个(0.25 表1 8个多态SSRs引物信息 Tab.1 Primer information for eight pairs of polymorphic SSRs 表2 3个群体在8个微卫星位点上的遗传多样性信息 Tab.2 The polymorphism information of eight microsatellite loci in three populations 8个位点在3个群体中扩增的等位基因片段大小范围为91~205 bp(表3),3个群体均存在稀有等位基因(0<基因频率<0.050),墨西哥湾扇贝、扇贝“渤海红”及杂交子代分别存在15、29和13个稀有等位基因。各个微卫星位点在3个群体的等位基因频率分布不均,群体间存在一定的遗传差异。 墨西哥湾扇贝、扇贝“渤海红”及杂交子代的遗传距离及遗传相似度见表4。结果显示,3个群体间均具有较高的遗传相似度,其中,扇贝“渤海红”与杂交子代的遗传相似度(0.888)高于墨西哥湾扇贝与杂交子代(0.736),墨西哥湾扇贝与扇贝“渤海红”遗传相似率最小(0.714)。杂交子代与墨西哥湾扇贝的遗传距离(0.279)高于扇贝“渤海红”与杂交子代的遗传距离(0.118)。杂交子代的遗传结构更倾向亲本扇贝“渤海红”。 杂交子代、墨西哥湾扇贝、扇贝“渤海红”的平均多态信息含量分别0.358、0.4376、0.462(表5~表7),说明3个群体都具有中等遗传多样性(0.25<0.5)。其中,扇贝“渤海红”平均期望杂合度、平均观测杂合度和平均有效等位基因最高,分别为0.2971、0.5045和2.394。杂交子代平均期望杂合度、平均观测杂合度、平均有效等位基因最低,分别为0.2458、0.4230和1.8316。 群体间的基因流(N)和遗传分化指数(F)是反应群体遗传分化的重要指标(表8)。当0<F<0.05时,说明群体的遗传分化程度很弱;当0.05<F<0.15时,群体处于中等程度的遗传分化;当0.15<F<0.25时,群体有较大的遗传分化;当F>0.25时,群体的遗传分化极大(Wright, 1951)。在本研究中,3个群体8个微卫星位点F值介于0.0825~0.2755,平均值为0.1398,表明13.98%的遗传变异由群体分化而导致,86.02%的遗传变异来源于群体内部,说明群体中存在较低水平的随机交配,3个群体平均基因流(N)为1.5387,平均固定系数(F)为0.3698。3个群体间出现中等程度的遗传分化。 表3 3个群体8个微卫星位点上的等位基因频率 Tab.3 Allele frequency of three populations in eight microsatellite loci 表4 墨西哥湾扇贝、扇贝“渤海红”及其杂交子代的遗传距离(对角线下方)和遗传相似系数(对角线上方) Tab.4 Genetic similarity coefficient (above the diagonal) and distance (below the diagonal) of three populations 表5 杂交子代在8个微卫星位点的遗传多样性参数 Tab.5 Genetic diversity index of hybrid (A. irradians concentricusדBohai Red”) in eight microsatellite loci 表6 墨西哥湾扇贝在8个微卫星位点的遗传多样性参数 Tab.6 Genetic diversity index of A. irradians concentricus in eight microsatellite loci 表7 扇贝“渤海红”在8个微卫星位点的遗传多样性参数 Tab.7 Genetic diversity index of “Bohai Red” in eight microsatellite loci 表8 3个群体8个微卫星位点统计量及基因流值 遗传多样性可用于评估一个种群抵御外界环境压力的能力及进化的潜力。在水产动物育种中,遗传多样性指数常作为一个养殖品种是否需要进行遗传改良的基础指标之一。本研究中,墨西哥湾扇贝遗传多样性参数(PIC0.358)明显低于扇贝“渤海红”(PIC= 0.462)。研究表明,亲本数量的减少会引起一些低频率位点的丢失(万俊芬等, 2004a)。在贝类人工养殖过程中的近亲繁殖、遗传漂变及瓶颈效应使得养殖群体基因库中某些等位基因丢失,造成养殖群体遗传多样性和遗传变异度降低(吕林兰等, 2008)。墨西哥湾扇贝养殖群体最初引种基础群体小(222个),且经过20多代的累代养殖(张福绥等, 1994),遗传多样性出现明显下降。作为新兴的养殖品种扇贝“渤海红”群体,其遗传多样性参数接近高度遗传多样性(PIC>0.5),说明扇贝“渤海红”仍具有较大的选育潜力。 杂种优势是指杂交子代在生长、存活等方面均优于双亲的现象(张国范等, 2004)。一般认为,杂种优势的强弱与亲本自身的遗传差异有关,即在一定的范围内,亲本间的遗传差异越大,其杂交子代往往具有更高的杂种优势(魏继海等, 2016; 谢晓东等, 2003)。而亲本间的遗传差异大小可以用遗传距离来衡量(张佳佳等, 2018)。本研究中,墨西哥湾扇贝与扇贝“渤海红”的遗传相似度为0.7146,遗传距离为0.3361。根据Thorpe(1982)提出的同种群体遗传相似性系数应介于0.80~0.97,而遗传距离应介于0.03~0.20的观点,本研究结果中,亲本群体这2个参数均不在同种群体范围内,说明2个群体间存在明显的遗传差异。理论上,二者杂交能够产生杂种优势明显的杂交子代。研究表明,墨西哥湾扇贝×扇贝“渤海红”的杂交子代生长速度与亲本墨西哥湾扇贝相比,提高了20%,具有明显的生长优势,其结果也验证了本研究的推论(谭杰, 2018)。但贝类遗传育种的实践及理论证明了杂种优势利用的关键不仅在于亲本间的遗传差异,还需要亲本间具有较高的配合力(张国范等, 2004)。这在海湾扇贝自交家系与交杂家系(郑怀平等, 2004)及太平洋牡蛎()近交系间杂交(Hedgecock, 1995)的杂种优势比较中均证明了这一点。因此,在扇贝“渤海红”与墨西哥湾扇贝2个亲本群体中选择更高配合力亲本个体可能会加强杂交子代杂种优势的利用。 在贝类杂交育种中,杂交子代在遗传结构偏向亲本一方的现象已经有较多报道,如合浦珠母贝()×珠母贝()的杂交子代在核型和同工酶上均与合浦珠母贝一致(姜卫国等, 1983; 李刚等, 1983);栉孔扇贝×虾夷扇贝的杂交F1代均偏向各自的母本(何斌等, 2007);栉孔扇贝×华贵栉孔扇贝的杂交F1代在遗传上也同样更加偏向母本栉孔扇贝(万俊芬等, 2004b)。本研究中,杂交子代与亲本扇贝“渤海红”的遗传距离为0.1188,与亲本墨西哥湾扇贝的遗传距离为0.279,杂交贝在遗传结构上更偏向于亲本扇贝“渤海红”。研究表明,杂交子代偏向某一方亲本,与杂交过程中染色体重组、突变和异染色质附近区域的SSR标记的偏分离聚集(张玉山等, 2008),以及在胚胎发育过程中染色体的选择性丢失等相关(Estoup, 2002; Fujiwara, 1997)。但也有学者认为,亲本遗传物质对后代贡献率不一致是因为两亲本基因纯合度不同,基因纯度高亲本基因型在后代中被检测几率增加(蔡磊等, 2012)。亲本基因纯合度及遗传物质的重组、变异等原因可能造成杂交子代在遗传结构上更加偏向于亲本扇贝“渤海红”。 杂交能够提高杂交子代的遗传多样性,如日本沼虾()太湖群体和鄱阳湖群体杂交后,其杂交子代的观测杂合度有明显的提高(李瀚声等, 2011);马氏珠母贝近交家系遗传多样性显著低于杂交家系(张嘉丽等, 2015)。但本研究中,杂交子代的遗传参数(N、H、H和PIC)均低于亲本墨西哥湾扇贝和扇贝“渤海红”。同样的现象也出现在紫扇贝×海湾扇贝的杂交实验中(孙妍等, 2013)。这可能与杂交实验中使用的亲本数量较少(5个)有关。研究表明,当亲本数量低于有效亲本数量时,会导致子代种群遗传变异力下降(Su, 1996);张海滨等(2005)等也发现,海湾扇贝过低的亲本数量能够显著影响子代的生长及存活。贝类配子具有多产性,贝类杂交育种常采用少数亲本杂交的方式,这种杂交方式可能导致子代中亲本部分等位基因丢失而影响子代的遗传变异程度,不利于后续杂交子代继代选育。此外,本研究中杂交子代群体选用了橙色群体,这也可能造成杂交子代遗传多样性低于双亲。 Abbott RT. American seashells (2nded). New York: Van Nostrand Reinhold Company, 1974 Cai L, Bai JJ, Li SJ,Genetic analysis of northern largemouth bass, Florida largemouth bass, and their reciprocal hybrids. Journal of Fishery Sciences of China, 2012, 19(1): 70–76[蔡磊, 白俊杰, 李胜杰, 等. 大口黑鲈北方亚种和佛罗里达亚种及其杂交子代的遗传分析. 中国水产科学, 2012, 19(1): 70–76] Cronquist A The evolution process: A critical study of evolutionary theory (2nded). New York: Columbia University Press, 1991 Estoup A, Jarne P, Cornuet JM. Homoplasy and mutation model at microsatellite loci and their consequences for population genetics analysis. Molecular Ecology, 2002, 11(9): 1591– 1604 Feng W, Li JL, Cai DQ,. Comparison of development and growth of the F1and F2hybrids between the Peruvian and Bay scallop. Chinese Agricultural Science Bulletin, 2012, 28(20): 118–124 [丰玮, 李建立, 蔡德泉, 等. 紫扇贝与海湾扇贝杂交后代生长发育的比较研究. 中国农学通报, 2012, 28(20): 118–124] Fujiwara A, Abe S, Yamaha E,. Uniparental chromosome elimination in the early embryogenesis of the inviable salmonid hybrids between masu salmon female and rainbow trout male. Chromosoma, 1997, 106(1): 44–52 Guo X, Zeng ZN, Zheng YY,. Genetic diversity in selected lines ofJournal of Fishery Sciences of China, 2018, 25(5): 1131–1136[郭香, 曾志南, 郑雅友, 等. 福建牡蛎选育群体的遗传多样性. 中国水产科学, 2018, 25(5): 1131–1136] Hang FF, Zhang JB, Li L,. Genetic diversity in two- generation of cultured Japanese scallop (()) populations. Marine Sciences, 2012, 36(9): 1–8[韩斐斐, 张继彪, 李莉, 等. 虾夷扇贝养殖群体及其子代的遗传多样性分析. 海洋科学, 2012, 36(9): 1–8] He B, Yang AG, Wang YQ,. Segregation of ISSR markers in hybrid larvae of scallop×. Marine Fisheries Research, 2007, 28(6): 32–38[何斌, 杨爱国, 王清印, 等. 栉孔扇贝×虾夷扇贝单对杂交子一代幼虫ISSR标记的分离方式. 海洋水产研究, 2007, 28(6): 32–38] Hedgecock D, McGoldrick DJ, Bayne BL. Hybrid vigor in Pacific oysters: An experimental approach using crosses among inbred lines. Aquaculture, 1995, 137(1): 285–298 Jiang WG, Wei Y, Li G. Studies on the cultivated interspecific hybridizations between pairs ofand(Mollusca, Bivalvia)Ⅱ. Observation on the chromosomes of fertilization and hybrids. Tropic Oceanology, 1983, 2(4): 316–320[姜卫国, 魏贻尧, 李刚. 合浦珠母贝、长耳珠母贝和大珠母贝种间人工杂交的研究——Ⅱ. 受精过程和杂交后代的染色体观察. 热带海洋, 1983, 2(4): 316–320] Li DM, Yang AG, Wu B,Development and application of the EST-SSR markers inProgress in Fishery Sciences,2017, 38(2): 137–142[李东明, 杨爱国, 吴彪, 等. 栉江珧()EST-SSR标记的开发与应用. 渔业科学进展, 2017, 38(2): 137–142] Li G, Jiang WG, Wei YY. Studies on the cultivated interspecific hybridizations between pairs ofand(Mollusca, Bivalvia)Ⅲ. Comparative study on zymograms. Tropic Oceanology, 1983(4): 321-328[李刚, 姜卫国, 魏贻尧. 合浦珠母贝、长耳珠母贝和大珠母贝种间人工杂交的研究——Ⅲ. 同工酶谱的比较研究. 热带海洋, 1983(4): 321–328] Li HS, Feng JB, Xie N,. Genetic structure analysis of parents and F1generation groups offrom Taihu Lake and Poyang Lake. Journal of Shanghai Ocean University, 2011, 20(3): 321–327 [李瀚声, 冯建彬, 谢楠, 等. 日本沼虾太湖和鄱阳湖群体及其F1的遗传结构分析. 上海海洋大学学报, 2011, 20(3): 321–327] Li JL, Wang CD, Li ZX,. Growth and path analyses of hybrid families between the Peruvian and Bay scallop, Marine Sciences, 2012, 36(8): 15–20 [李建立, 王春德, 李朝霞, 等. 紫扇贝和海湾扇贝杂交家系的生长和通径分析. 海洋科学, 2012, 36(8): 15–20] Liu BZ. Study on the population genetics of Bay scallopand the molecular phylogeny of Pectinidae. Doctoral Dissertation of Institute of Oceanology, Chinese Academy of Sciences, 2003 [刘保忠. 海湾扇贝群体遗传学和扇贝科分子系统演化的研究. 中国科学院海洋研究所博士研究生学位论文, 2003] Liu ZG, Wang H, Fu SW. Morphological growth of culturedSay in Beibu Bay in Zhanjiang. Journal of Fisheries of China, 2007a, 31(5): 675–681[刘志刚, 王辉, 符世伟. 湛江北部湾养殖墨西哥湾扇贝的形态增长规律. 水产学报, 2007a, 31(5): 675–681] Liu ZG, Wang H, Li ZM,. Upper incipient lethal temperature ofSay. Journal of Fishery Sciences of China, 2007b, 14(5): 778–785[刘志刚, 王辉, 栗志民, 等. 墨西哥湾扇贝高起始致死温度的研究. 中国水产科学, 2007b, 14(5): 778–785] Lü LL, Du XD, Wang Y,. Genetic diversity of three populations and the first generations of hybridization between different populations of pearl oyster,(Dunker). Acta Hydrobiologica Sinica, 2008, 32(1): 26–32[吕林兰, 杜晓东, 王嫣, 等. 马氏珠母贝3个野生种群及种群间杂交后代遗传多样性的ISSR分析. 水生生物学报, 2008, 32(1): 26–32] Nan YH, Zhang JS, FengW,. Inter-specific hybridization betweenand. Chinese Agricultural Science Bulletin, 2012, 28(20): 131–135 [南乐红, 张金盛, 丰玮, 等. 紫扇贝和墨西哥湾扇贝种间杂交的初步研究. 中国农学通报, 2012, 28(20): 131–135] Nie HT, Li Q, Kong LF. Development of four multiplex PCR panels of microsatellites and application to kinship analysis in the Pacific abalone (). Journal of Fisheries of China, 2013, 37(2): 207–215 [聂鸿涛, 李琪, 孔令锋. 皱纹盘鲍微卫星多重PCR体系构建及其在家系鉴定中的应用. 水产学报, 2013, 37(2): 207–215] Pan YD, Feng J, Liu H,. Reciprocal crosses betweenand. Journal of Fishery Sciences of China, 2017, 24(4): 698–709 [潘奕达, 封杰, 刘行, 等. 墨西哥湾扇贝与华贵栉孔扇贝的远缘杂交. 中国水产科学, 2017, 24(4): 698–709] Pan YD. A preliminary study on the inter-genus hybridization betweenSay and. Master´s Thesis of Guangxi University, 2016[潘奕达. 墨西哥湾扇贝与华贵栉孔扇贝属间远缘杂交的初步研究. 广西大学硕士研究生学位论文, 2016] Peng ZM, LiuFu SM, Liu ZG. Research progress of artificial culture and genetic breeding ofSay. Journal of Guangdong Ocean University, 2014, 34(1): 91–97 [彭张明, 刘付少梅, 刘志刚. 墨西哥湾扇贝人工养殖及遗传育种研究进展. 广东海洋大学学报, 2014, 34(1): 91–97] Qian YQ, Ma KP. Principles and methods of biodiversity research. Beijing: China Science and Technology Press, 1994[钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994] Su GS, Liljedahl LE, Gall G. Effects of inbreeding on growth and reproductive traits in rainbow trout (). Aquaculture, 1996, 142(3–4): 139–148 Sun Y, Huang XT, Hu LP,. AFLP analysis of genetic structure of,and their positive and negative hybrid offspring. Marine Sciences, 2013, 37(8): 1–5[孙妍, 黄晓婷, 胡丽萍, 等. 紫扇贝、海湾扇贝及其正反杂交子代群体遗传结构的AFLP分析. 海洋科学, 2013, 37(8): 1–5] Tan J, Peng XJ, Liu ZG,. SSR analysis of genetic diversity ofand Zhongkehong. Genomics and Applied Biology, 2018, 37(4): 1397–1405[谭杰, 彭晓君, 刘志刚, 等. 墨西哥湾扇贝与“中科红”海湾扇贝群体遗传多样性SSR分析. 基因组学与应用生物学, 2018, 37(4): 1397–1405] Tan J. Transcriptome analysis ofand study on the molecular markers of its progeny crossed with‘’. Master´s Thesis of Guangdong Ocean University, 2018 [谭杰.墨西哥湾扇贝转录组分析及其与扇贝“渤海红”杂交F1代分子标记研究. 广东海洋大学硕士研究生学位论文, 2018] Thorpe JP. The molecular clock hypothesis: Biochemical evolution, genetic differentiation and systematics. Annual Review of Ecology and Systematics, 1982, 13: 139–168 Wan JF, Bao ZM, Liu GX,. Segregation of ISSR markers in hybrid larvae of two scallop species. Chinese High Technology Letters, 2004b, 14(5): 82–87[万俊芬, 包振民, 刘广旭, 等. 扇贝种间单对杂交一代幼虫ISSR标记的分离方式. 高技术通讯, 2004b, 14(5): 82–87] Wan JF, Bao ZM, Wang XL,. The influence of parental stock size on the genetic structure of. Journal of Fisheries of China, 2004a, 28(2): 127–132[万俊芬, 包振民, 汪小龙, 等. 亲本数目对鲍养殖群体AFLP标记位点及其遗传结构的影响. 水产学报, 2004a, 28(2): 127–132] Wang C, Liu B, Li J,. Introduction of the Peruvian scallop and its hybridization with the Bay scallop in China. Aquaculture, 2011, 310(3–4): 380–387 Wang C, Liu B, Liu X,. Selection of a new scallop strain, the Bohai Red, from the hybrid between the Bay scallop and the Peruvian scallop. Aquaculture, 2017, 479: 250–255 Wang Y, Wang X, Wang A,. A 16-microsatellite multiplex assay for parentage assignment in the eastern oyster (Gmelin). Aquaculture, 2010, 308(1): S28–S33 Wei JH, Zhao JL, Wu JW,. Genetic characterization of Oreochromis niloticus (♀) × Sarotherodon melanotheron (♂) hybrid F2and F3by microsatellite analysis, South China Fisheries Science, 2016, 12(1): 30–35[魏继海, 赵金良, 吴俊伟, 等. 尼罗罗非鱼(♀)×萨罗罗非鱼(♂)杂交F2与F3群体遗传特征的微卫星分析. 南方水产科学, 2016, 12(1): 30–35 Wright S. Genetical structure of populations. Annals of Eugenics, 1951, 15(1): 323–354 Xie XD, Ni ZF, Meng FR,. Relationship between differences of gene expression in early developing seeds of hybrid versus parents and heterosis in wheat. Acta Genetica Sinica, 2003, 30(3): 260–266 [谢晓东, 倪中福, 孟凡荣, 等. 小麦杂交种与亲本发育早期种子的基因表达差异及其与杂种优势关系的初步研究. 遗传学报, 2003, 30(3): 260–266] Zhang FS, He YZ, Qi LX,. Introduction and spat-rearing of the F1generation ofSay. Oceanologia et Limnologia Sinica, 1994, 25(4): 372–377[张福绥, 何义朝, 亓铃欣, 等. 墨西哥湾扇贝的引种和子一代苗种培育. 海洋与湖沼, 1994, 25(4): 372–377] Zhang GF, Liu X, Que YH,. The theory and application of hybridization and heterosis in marine mollusks. Marine Sciences, 2004, 28(7): 54–60 [张国范, 刘晓, 阙华勇, 等. 贝类杂交及杂种优势理论和技术研究进展. 海洋科学, 2004, 28(7): 54–60] Zhang GM, Sun XJ, Wu BTransferability of EST-SSR frominto. Progress in Fishery Sciences, 2018, 39(4): 139–146[张广明, 孙秀俊,吴彪, 等. 虾夷扇贝EST-SSR标记在栉孔扇贝中的通用性研究. 渔业科学进展, 2018, 39(4): 139–146] Zhang H, Liu X, Zhang G,. Growth and survival of reciprocal crosses between two Bay scallops,Say andLamarck. Aquaculture, 2007, 272(S1): S88–S93 Zhang HB, Liu X, Zhang GF,Effects of effective population size on the F1growth and survival of Bay scallop.Acta Oceanologica Sinica, 2005, 27(2): 177–180[张海滨, 刘晓, 张国范, 等. 不同有效繁殖群体数对海湾扇贝Fl生长和存活的影响. 海洋学报, 2005, 27(2): 177–180] Zhang JJ, Li J, Zhang GS,. Microsatellite-based analysis of genetic diversity of parent and hybrid of yellow catfish(♀)×(♂). Fisheries Science, 2018, 37(5): 612–621[张佳佳, 李杰, 张国松, 等. 杂交黄颡鱼(黄颡鱼♀×瓦氏黄颡鱼♂)及其双亲遗传多样性的微卫星分析. 水产科学, 2018, 37(5): 612–621 Zhang JL, Luo YQ, Huang Y,. Genetic structure of inbred and hybrid families of pearl oyster. Genomics and Applied Biology, 2015, 34(4): 723–730 [张嘉丽, 罗燕秋, 黄怡, 等. 马氏珠母贝近交与杂交家系的遗传结构比较. 基因组学与应用生物学, 2015, 34(4): 723–730 Zhang XY, Zhang XJ, Zhao C,. The development of BAC-end sequence-based microsatellite markers and analysis on population genetic diversity in Zhikong scallop (). Journal of Fisheries of China, 2012, 36(6): 815–824 [张秀英, 张晓军, 赵翠, 等. 栉孔扇贝BES-SSR的开发及遗传多样性分析. 水产学报, 2012, 36(6): 815–824] Zhang YS, Chen QQ, Wu W,. Integration of centromeric regions on the rice molecular linkage map of SSR markers. Journal of Huazhong Agricultural University, 2008, 27(2): 167–171 [张玉山, 陈庆全, 吴薇, 等. 水稻SSR标记遗传连锁图谱着丝粒的整合及其偏分离分析. 华中农业大学学报, 2008, 27(2): 167–171] Zhao XX, Deng YW, Du XD,. Genetic structure of four progeny stocks ofby microsatellite markers analysis. Genomics and Applied Biology, 2010, 29(5): 879–884 [赵晓霞, 邓岳文, 杜晓东, 等. 利用微卫星标记分析马氏珠母贝4个养殖群体遗传结构. 基因组学与应用生物学, 2010, 29(5): 879–884] Zheng H, Xu F, Zhang G. Crosses between two subspecies of Bay scallopand heterosis for yield traits at harvest. Aquaculture Research, 2011, 42(4): 602– 612 Zheng HP, Zhang GF, Liu X,. Comparison of growth and survival between the self-fertilized and hybridized families inJournal of Fisheries of China, 2004, 28(3): 267–272 [郑怀平, 张国范, 刘晓, 等. 海湾扇贝杂交家系与自交家系生长和存活的比较. 水产学报, 2004, 28(3): 267–272] Genetic Analysis of, “Bohai Red” and Their Hybrids YAO Gaoyou1,3, TAN Jie1,3, WU Yuyuan1, SU Xiaoying1, LIUFU Shaomei2, ZHANG Yuan1,3, FANG Jiaxi4, CHEN Nansheng5, WANG Chunde6, LIU Zhigang1,3①� (1. Fisheries College of Guangdong Ocean University, Zhanjiang 524088; 2. Zhanjiang Silverwave Marine Biotechnology Co.Ltd., Zhanjiang 524022; 3. Guangdong Province Nanhai Economic Invertebrates Healthy Breeding Engineering Research Center, Zhanjiang 524088; 4. Department of Applied Biology and Chemical Technology of Hong Kong Polytechnic University, Hong Kong 999077;5. Department of Molecular Biology and Biochemistry of Simon Fraser University, British Columbia V5A 1S6; 6. College of Marine Science and Engineering, Qingdao Agricultural University, Qingdao 266109) Hybridization between the ‘Bohai Red’ andwas conducted. The hybrids exhibited heterosis in terms of survival rate, growth rate, and heat tolerance, demonstrating substantial potential to replace. To ensure the efficient use of these hybrids, we need to understand the genetic structure of the hybrids and their parents. Microsatellites are widely used to assess the genetic diversity of aquaculture species and to construct a quantitative trait locus due to their characteristics such as abundant polymorphism, rich information, and high codominance and conservation. In this research, the genetic diversityof 90 individuals in 3 populations of, ‘Bohai Red’, and their hybrids was analyzed using 8 pairs of microsatellite primers. A total of 67 alleles were amplified, and 4~14 alleles at each locus were generated, with a mean allele number of 8.5. Effective allele number, expected heterozygosity, and polymorphic content were the highest in ‘Bohai Red’ (2.3947, 0.504, and 0.462, respectively). The genetic differentiation index, gene flow, and fixed coefficient were 0.1398, 1.5387, and 0.3698, respectively. Between the hybrids and ‘Bohai Red’, the genetic distance was the smallest (0.1188) and the genetic similarity was the highest (0.888), suggesting their similar genetic structures. These findings provide a reference for the genetic analyses of ‘Bohai Red’ andas well as for the breeding of new varieties. ; “Bohai Red”; Hybridization; Genetic diversity LIU Zhigang, E-mail: liuzg919@126.com S966.9 A 2095-9869(2020)05-0014-09 10.19663/j.issn2095-9869.20190502001 http://www.yykxjz.cn/ 姚高友, 谭杰, 吴羽媛, 苏晓盈, 刘付少梅, 张元, 方家熙, 陈楠生, 王春德, 刘志刚. 墨西哥湾扇贝和扇贝“渤海红”及其杂交子代的遗传分析. 渔业科学进展, 2020, 41(5): 118–126 Yao GY, Tan J, Wu YY, Su XY, Liufu SM, Zhang Y, Fang JX, Chen NS, Wang CD, Liu ZG. Genetic analysis of, “Bohai Red” and their hybrids. Progress in Fishery Sciences, 2020, 41(5): 118–126 * 广东海洋大学2017年“高水平大学”省财政资金支持项目(002026003003)、2019广东省重点领域研发计划(2019B020238002)和2019广东省现代农业产业技术体系创新团队(2019KJ146)共同资助[This work was supported by Guangdong Ocean University Project of “High-level University”(002026003003) in 2017, Key Research and Development Program of Guangdong (2019B020238002), and 2019 Innovation Team of Modern Agricultural in Guangdong(2019KJ146)]. 姚高友,E-mail: gaoyouyao@126.com 刘志刚,教授,E-mail:liuzg919@126.com 2019-05-02, 2019-07-10 (编辑 马璀艳)

2.2 等位基因频率
2.3 遗传距离
2.4 3个群体遗传多样性分析
2.5 群体间遗传分化

3 讨论
3.1 亲本群体遗传多样性
3.2 杂种优势与亲本遗传距离
3.3 杂交子代遗传偏向性与遗传变异
