利用鸡胚表达谱数据筛选皮肤发育相关基因的研究
2020-09-18樊庆灿
樊庆灿
(宜春学院生命科学与资源环境学院,江西宜春 336000)
动物皮肤在体温调节、保护、感觉、伪装和交配中发挥重要作用,这些功能是通过皮肤及其衍生物特有的生理结构完成的。动物发育过程中外胚层表皮与中胚层真皮相互作用,分化成多种多样的皮肤和衍生物[1]。在皮肤发育的研究中,鸡是一种理想模型,这是由于鸡躯体被覆羽毛,胫部含有鳞片状皮肤表面,代表了比较典型的皮肤结构,对其发育调节的研究有助于促进对人类皮肤发育机理的研究。在禽类皮肤研究中,发现多个基因参与皮肤发育过程,如成纤维细胞生长因子10(Fibroblast Growth Factor 10,FGF10)的表达促进羽芽发育[2],骨形态发生蛋白2(Bone Morphogenetic Protein 2,BMP2)和骨形态发生蛋白4(Bone Morphogenetic Protein 4,BMP4)可以通过抑制羽芽发育调节羽径大小[3],德尔塔1 基因(Delta1,DELTA1)诱导鳞片皮肤发育[4],BMP 受体蛋白等则将鳞片转化为羽毛[5],Wnt 信号通路参与了整个皮肤形态发生过程[6]等。鸡发育初期的皮肤上皮(7 d 羽毛皮肤和9 d 鳞片皮肤)具有高度可塑性,蕴含分化为羽毛或鳞片的潜能[7],决定早期胚胎羽毛和鳞片皮肤的命运。本研究利用鸡胚表达谱数据[8],结合生物信息技术,筛选影响鸡胚胎羽毛和鳞片皮肤发育的调控通路和相关基因。本研究成果将有利于人类皮肤发育机理的研究。
1 材料方法
1.1 数据采集 本研究表达谱数据来自基因表达数据库,编号为GSE62882(https:// www.ncbi.nlm.nih.gov/geo/query/acc.cgi),样品信息见表1,包括白来航鸡7日龄胚胎背侧皮肤上皮样品(FE7)2 个、背侧皮肤间充质(FM7)样品2 个,9 日龄胚胎背部皮肤上皮(FE9)样品3 个、背部皮肤间充质(FE9)3 个,9 日龄胚胎跖皮肤上皮样品(SE9)2 个、跖皮肤间充质(SM9)2个,11 日龄胚胎跖皮肤上皮(SE11)3 个、跖皮肤间充质(SM11)3 个,共20 样品,采用Affymetrix Chicken Genome Array(GPL3213)进行表达谱测序,获得20个样品33 457 个转录本的数据[8]。
1.2 统计软件 表达谱数据的转换、表达差异分析使用在线软件GEO2R(https://www.ncbi.nlm.nih.gov/geo/geo2r/)。显著基因的互作分析使用在线软件STRING(https://string-db.org/cgi/input.pl? sessionId=NPdugdy y2MoP & InInput_page_show_ search=on),基因互作分析图、核心基因计算作图和GO 分析作图均使用软件Cytoscape 3.6.1。基因通路分析(KEGG)使用在线软 件DAVID(https://david.ncifcrf.gov/tools.jsp),韦恩图解分析使用在线软件Veen Diagram(http://bioinformatics.psb.ugent.be/webtools/Venn/)。

表1 20 个样本的信息统计表
1.3 分析方法 GEO2R 中输入编号GSE62882,获取20个样本信息,进行log 转化后绘制箱式图观察表达谱数据分布,并按照表1 将样本定义为8 个组,按照FE7-FE9、FM7-FM9、SE9-SE11 和SM9-SM11 两两进行表达差异分析,并分别命名为FE7-9、FM7-9、SE9-11 和SM9-11,选择False discovery rate(FDR)法进行P值调整(Padj),基因注释选择NCBI。
4 组差异基因(P<0.01)分别输入到Venn diagram中进行韦恩图解分析,获得差异分析的共同基因。使用String 对4 组显著基因(P<0.01)进行基因互作分析,根据参与互作网络的基因数目的差异,确定最低综合得分值,导出4 组分析差异基因的互作网络数据,并导入Cytoskype 中作图,删除位于互作图边缘地带的基因。核心基因的计算使用cytoHubb 插件的Betweenness 算法,并对核心基因进行合并分析。差异基因(P<0.01)的GO 分析采用Cytoskype 软件中的BINGO 插件,KEGG 分析使用在线软件DAVID。
2 结果
2.1 数据标准化和箱式图 如图1 所示,样本的下、上边缘值分别为(2.43±0.03)和(13.39±0.33),下、上四分之一值分别为(4.15±0.07)和(7.85±0.09),中位数为(5.65±0.02),说明20 个样本表达量数据比较一致,可以进行转录本的差异分析。

图1 20 个样本表达量数据标准化后的统计信息
2.2 基因表达差异分析 由表2 可知,FE7-9 筛选出28个Padj<0.05 和590 个P<0.01 的基因,9 日龄与7 日龄相比,191 个基因(P<0.01)上调,399 个基因(P<0.01)下调,上调基因细胞基质蛋白(Periostin,POSTN)是Padj最小且差异倍数最大的基因。FM7-9 筛选出1 个Padj<0.05和473 个P<0.01 的基因,其中185 个基因(P<0.01)上调,288 个基因(P<0.01)下调,上调基因海绵糖基因(Spondin 2,SPON2)的差异倍数最大。SE9-11 筛选出116 个Padj<0.05 和795 个P<0.01 的基因,11 日龄与9 日龄相比,上调基因(P<0.01)292 个,下调基因(P<0.01)503 个,其中下调基因丝氨酸/苏氨酸激酶基因(Serine/Threonine Kinase 32B,STK32B)的Padj最小且差异倍数最大。SM9-11 筛选出0 个Padj<0.05 和73个P<0.01 的基因,45 个基因(P<0.01)上调(P<0.01),28 个基因(P<0.01)下调,其中上调基因角蛋白5 基因(Keratin 5,KRT5)的差异倍数最大。
4 组差异基因(P<0.01)韦恩图解分析结果如图2 所示,脂肪酸结合蛋白(Fatty Acid Binding Protein7,FABP7)在4 组差异分析中均为差异基因(P<0.01),FE7-9、FM7-9 和SE9-11 有27 个共同基因。FE7-9 和SE9-11含有184 个共同基因(FE-SE)。对FE7-9、FM7-9、SE9-11、SM9-11 和FE-SE 的基因进行核心基因分析并合并作图(图3),可见核心基因组成一个较大的互作网络,并通过关键基因纤维连接蛋白1(Fibronectin1,FN1)和血小板源性生长因子(Platelet Derived Growth Factor Subunit B,PDFGB)连接在一起。

表2 差异表达分析筛选的显著基因信息
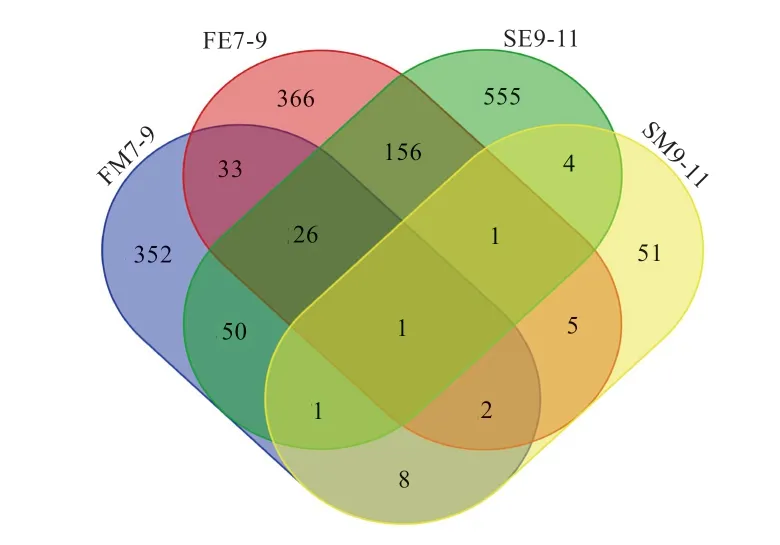
图2 4 组差异表达基因的韦恩图解分析

图3 4 组差异分析的核心基因合并后的互作图
2.3 差异基因的GO 分析 如图4 所示,FE7-9、FM7-9、SE9-11 和SM9-11 分别富集到21、8、20 个和0 个显著的条目(P<0.01),FE7-9、FM7-9 和SE9-11 筛选到3 个显著基因最多的共同GO 条目,从上到下分别为多细胞组织过程(0032501)、发育过程(0032502)和单细胞组织发育(0007275),这3 个条目在FM7-9 和SE9-11 的GO 分析中P值最小。关键核心基因PDFGB和基因POSTN存在于上述所有GO 条目中。FE7-9 中P值最小的GO 条目为生物黏附(0022610)和细胞黏附(0007155),关键的核心基因FN1和基因POSTN属于该条目。
2.4 差异基因的信号通路分析 如图5 所示,4 组差异基因分别筛选出5、7、10、2 条调节通路。FE7-9、FM7-9、SE9-11 中筛选的P值最小的信号通路分别为氮代谢(00910)、MAPK 信号通路(04010)和Wnt 信号通路(04310)。黏附斑激酶信号通路(04510)是SM9-11 通路分析中发现的P值最小的通路,也是4 组差异分析中筛选到的共同通路,FE7-9、FM7-9、SE9-11 和SM9-11 的差异基因中分别有12、10、17、4 个基因参与了此通路,FN1和PDGFB等5 个核心基因参与了此通路。
3 讨 论
3.1 差异表达基因分析 本研究筛选了鸡胚胎皮肤上皮和间充质组织的差异表达基因,羽毛和鳞片上皮组织中共筛选到144 个差异基因(Padj<0.05),间充质组织中只筛选到1 个差异表达基因(Padj<0.05),说明胚胎发育初期(7~11 d)的皮肤上皮比间充质更为活跃。9 胚龄与7 胚龄相比,背部羽毛上皮下调的基因数远远高于上调的基因数,11 胚龄与9 胚龄相比,鳞片上皮下调的基因数远远高于上调的基因数,可能是羽毛皮肤(9 d)、鳞片皮肤(11 d)已经历了细胞的发育高峰,胚胎发育初期的羽毛皮肤(7 d)和鳞片皮肤(9 d)是皮肤发育的关键时期[9]。韦恩图解分析中,FE7-9 与SE7-9 有较多的共同基因,羽毛皮肤和鳞片皮肤的发育过程具有很多的相似性。
3.2 差异基因的GO 分析 4 组差异表达基因的GO 分析中,筛选出3 条相同的GO 条目,下级条目单细胞组织发育指多细胞有机体从最初的状态(如合子)发展到后来状态的生物过程,包括胚胎分化和生长的整个过程。本研究差异基因的GO 分析富集在单细胞发育及相关条目基因数最多,因为皮肤发育是胚胎发育的一部分,出现这种结果也是必然的。FE7-9 富集在细胞黏附基因数均28 个。7~9 日龄胚胎是皮肤分化发育关键时期,本研究筛选到28 个黏附相关基因。在胚胎发育过程中,细胞与细胞、细胞和基质之间的黏附和定向细胞移动,是细胞分化和发育成特定组织器官的基础,在整个过程中,黏附分子发挥着重要作用[9],其广泛参与了鸡胚胎皮肤细胞的分化发育。这与本研究的结果一致。
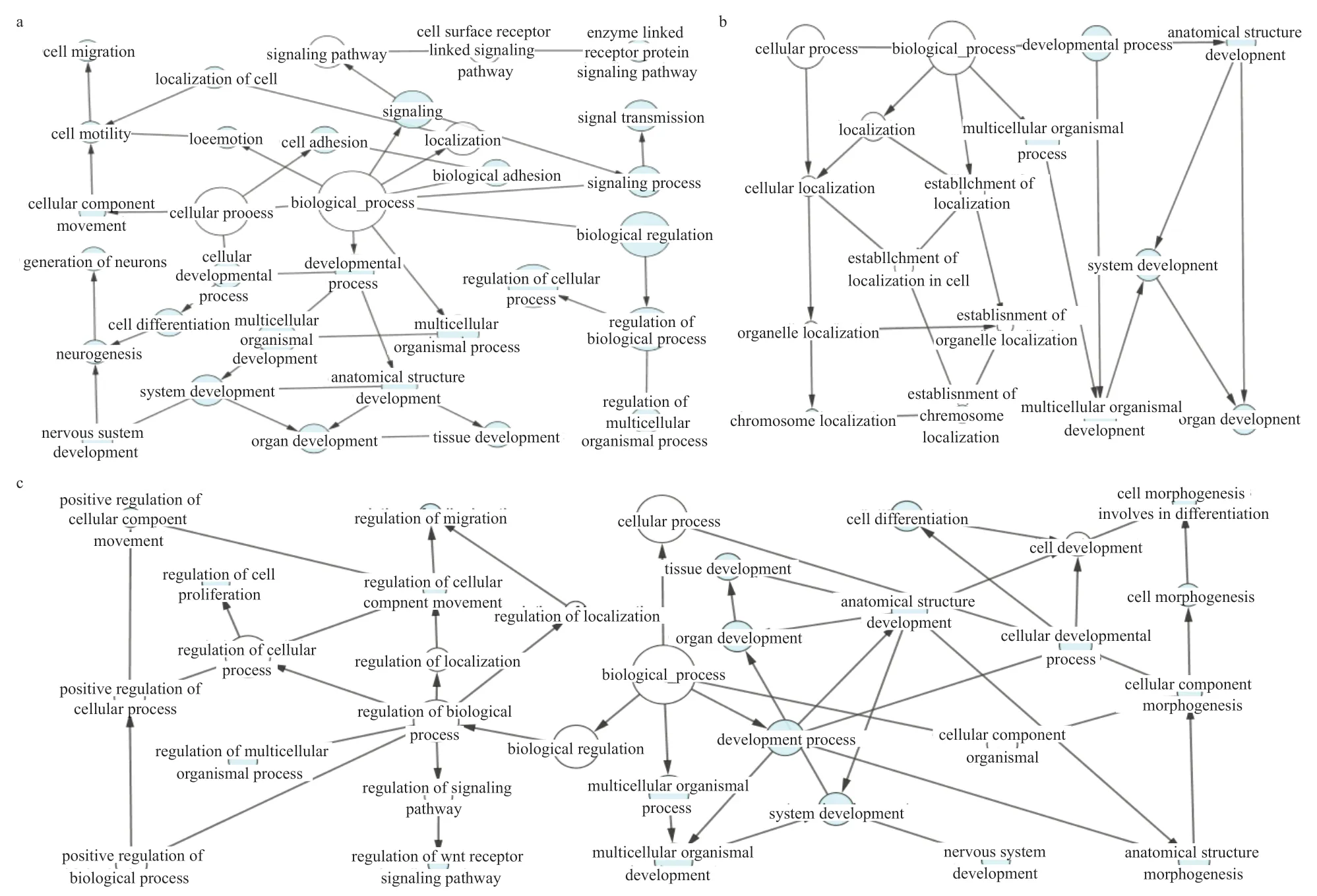
图4 差异表达基因GO 树状分析图

图5 差异表达基因KEGG 分析的显著通路
3.3 FAK 信号通路和关键基因 KEGG 分析中筛选出的共同信号通路是FAK 信号通路。FAK 通路参与调节细胞生长、凋亡、黏附和迁移等各细胞过程[10],并通过调节金属蛋白酶-9(Matrix Metallo Proteinase 9,MMP-9)活性导致细胞外基质的降解,调控细胞粘附性[11-13]。研究发现,细胞黏附在禽类羽毛图案的形成中起着中心作用[14],细胞黏附活动不仅促进细胞聚集形成真皮[15],稳定的细胞聚集体还能启动表皮板的形成[16]。本研究筛选出的基因中,PDFGB、FN1和POSTN都是FAK 信号通路相关的基因。PDFGB能刺激细胞合成大量特定的细胞外基质,引起细胞增殖、迁移和分化[17-18],其异常表达导致皮肤纤维瘤的发生[19-20]。FN1通过结构域与细胞表面受体、纤维蛋白等特异性结合,调控细胞的迁移、分化和黏附[21],参与胚胎细胞分化和器官形成[22]。POSTN参与细胞的募集和黏附,调节胚胎发育[23],并通过结合整合素类蛋白来激活FAK 信号途径发挥调节作用[24]。
目前禽类皮肤发育的研究较少,相关基因及调节机理并不完全清楚。本研究发现的黏附斑信号通路具有广泛的生物功能,但其对禽类皮肤发育的调节作用未见报道。3 个关键基因PDFGB、FN1和POSTN与FAK 信号通路有关,并在细胞黏附和胚胎发育中具有重要的调节作用。正如前人所言,细胞黏附活动是细胞分化和发育成特定组织器官的基础[9],FAK 信号通路介导的细胞黏附活动是禽类胚胎皮肤发育研究的重要方向。
4 结 论
本研究利用数据库数据和生物信息技术,筛选影响鸡胚胎皮肤发育的差异表达基因,发现胚胎发育早期(7 d羽毛皮肤和9 d 鳞片上皮)是皮肤发育的关键时期。筛选的基因中与单细胞组织发育相关的基因最多,羽毛上皮(7 d、9 d)部分差异基因富集在细胞黏附。FAK 信号通路介导的细胞黏附活动可能参与鸡胚胎皮肤的整个发育过程,关键基因PDGFB、FN1和POSTN是值得关注的候选基因。
