喀斯特区不同岩性下广玉兰幼苗的养分利用研究
2020-09-18符裕红
符裕红,彭 琴
(1.贵州师范学院生物科学学院,贵阳 550018;2.黔西南民族职业技术学院,贵州 兴义 562400)
广玉兰(MagnoliagrandifloraL.)为木兰科木兰属常绿乔木[1],耐干旱瘠薄、抗污能力强[2],耐烟抗风,抗寒性强[3],材质优良[4],且有较高的药用价值[5,6],适宜在庭院、行道、污染、退化区域种植[2],是进行植物恢复的优良树种。喀斯特区岩石类型多样,以贵州最为典型,土被浅薄,蓄水肥能力低[7-8];植被退化[9],生态破坏[10],植被亟待恢复,植被选择、配置、特性研究尤为重要。基于不同岩性特征的研究,大多表现在对土壤养分、土壤酶活性、土壤肥力等方面[11-14],结合岩性对植物生长的影响也主要体现在对马尾松[15]、构树[16]、日本柳杉[17]等类型,且主要针对生长方面的影响,未涉及到养分利用特征;结合岩性与广玉兰的研究甚少,主要从其存活,苗高、地径的生长量方面进行考量[12],但养分利用吸收的方面未见报道。在喀斯特地区选择不同的岩性研究广玉兰的养分利用,有利于全面表现喀斯特生境条件的基本特征,详细了解广玉兰对于喀斯特地区的生长特性及生境适应机制,从而为喀斯特地区的生态恢复服务,为该区的植被选择、配置及植被栽培提供科学的参考和指导。
1 材料与方法
1.1 试验材料
选择性质相当的一年生广玉兰幼苗,定植到分别装有玄武岩(清镇)、石英砂岩(龙里)、长石石英砂岩(黔陶)、变余砂岩(清镇)、第四纪红色黏土(孟关)、煤系砂页岩(彭官)土壤的石框中[12]。
1.2 试验方法
每种岩性土壤分别定植30株广玉兰幼苗,栽植密度为15株/m2,分别设置3个重复,实验期间按常规管理。土壤采集于定植2年后进行,每个石框设6个样点,以30 cm深度进行“S”形取样,共108个;部分土壤直接处理后进行土壤酶活性测定,部分土样处理风干后保存待用。
植物样品随机采摘每个石框中正常且大小一致的植株叶片,处理和粉碎后密封保存待用。
1.3 评价指标
土壤指标:土壤全氮、全磷、全钾、碱解氮、速效磷、速效钾;土壤蔗糖酶、淀粉酶、脲酶、磷酸酶、蛋白酶、过氧化氢酶、多酚氧化酶。土壤全氮采用蒸馏法,土壤全磷采用钼锑抗比色法,土壤全钾采用火焰光度法;土壤碱解氮采用扩散吸收法;速效磷采用钼锑抗比色法;速效钾采用火焰光度法[18-20];土壤蔗糖酶、淀粉酶的测定采用3,5-二硝基水杨酸比色法,脲酶采用苯酚钠-次氯酸钠比色法,磷酸酶磷酸苯二钠比色法,蛋白酶采用茚三酮比色法,过氧化氢酶采用容量法,多酚氧化酶采用碘量滴定法[21]。
植物指标:植物全氮、全磷、全钾;植物吸收系数Ax。植物全氮采用蒸馏法,植物全磷采用钼锑抗比色法,植物全钾采用火焰光度法;植物养分的吸收代表了植物适应生长环境的对策[22-23],采用生物吸收系数表示植被对各元素的吸收能力;具体计算公式如下:
Ax=Lx/nx
式中:Ax为x元素生物吸收系数;Lx为x元素在植物中的含量;nx为相应土壤中x元素的含量。
1.4 数据分析
测定结果采用Excel 2010软件和SPSS 22.0软件对数据进行处理分析。
2 结果与分析
2.1 不同岩性的土壤养分
经过方差分析,不同岩性的土壤养分存在显著差异(p<0.05);从图1可看出,有机质在玄武岩(岩性1)上表现最高,全氮、碱解氮、全磷均在此类型上体现出最大值;速效磷在煤系砂页岩(岩性6)上表现最大;全钾、速效钾在第四纪红色黏土(岩性5)上数值最高。经过不同岩性间的多重比较(LSD),岩性间两两差异显著。说明岩性对土壤养分含量的影响显著,导致提供给植物生长的条件差异。
2.2 不同岩性的土壤酶活性
对不同岩性土壤酶活性指标数值进行方差分析,结果表明,不同岩性的土壤酶活性除淀粉酶外,其它酶活性均表现为差异极显著(p<0.01)。从图2可看出,不同的土壤酶活性在不同岩性下均存在明显波动,其中,蔗糖酶在变余砂岩(岩性4)上最高,石英砂岩(岩性2)最低;脲酶在玄武岩(岩性1)上最高,且明显高于其它类型,长石石英砂岩(岩性3)最低;磷酸酶在煤系砂页岩(岩性6)上最高,第四纪红色黏土(岩性5)最低;蛋白酶在第四纪红色黏土(岩性5)上最高,变余砂岩(岩性4)最低;过氧化氢酶在玄武岩(岩性1)上最高,长石石英砂岩(岩性3)最低;多酚氧化酶在玄武岩(岩性1)上最高,石英砂岩(岩性2)最低。说明各种岩性的土壤酶活性变化差异较大,岩石类型也是导致发育土壤酶活性差异的主要原因。
2.3 不同岩性植物养分含量
不同岩性植物养分含量存在差异,通过对其进行方差分析可知,不同岩性条件对植物养分含量的影响均存在显著差异(p<0.05)。植物氮素、磷素在第四纪红色黏土(岩性5)含量最高,在玄武岩(岩性1)最低;植物钾素在石英砂岩(岩性2)上最高;钾素在第四纪红色黏土(岩性5)含量最高,在玄武岩(岩性1)上最低。经过多重比较(LSD)发现,除植物全氮在各岩性间无显著差异外,其它各种指标均随着岩性的变化均存在显著差异。说明不同岩性类型造成了植物体内养分元素含量差异,植物在各种岩性发育的土壤上对养分的吸收利用不尽相同,从而影响植物的生长。
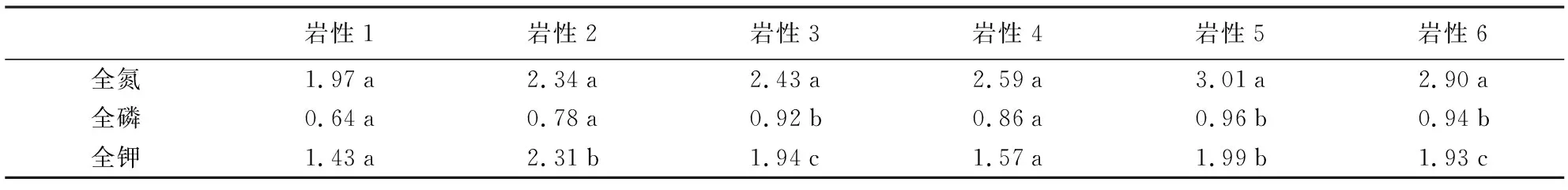
表1 不同岩性土壤及植物养分含量的变化
2.4 不同岩性上广玉兰的养分吸收
根据生物吸收系数公式计算广玉兰幼苗在不同岩石类型条件下的生物吸收系数,经过方差分析得出各类型条件下广玉兰幼苗的养分吸收系数差异显著(p<0.05)。从图3可看出,氮的吸收系数AN在玄武岩(岩性1)上的表现最低,在长石石英砂岩(岩性3)上最高;磷的吸收系数AP在玄武岩(岩性1)上表现最低,在石英砂岩(岩性2)上表现最高;钾的吸收系数AK在变余砂岩(岩性4)最低,在玄武岩(岩性1)上表现最高。说明不同岩性发育的土壤导致植物吸收系数差异显著,并进一步证实了关于岩性导致植物对养分吸收差异的论断。
2.5 不同岩性土壤养分与土壤酶的相关性
经过对不同岩性条件下的土壤养分、土壤酶活性指标及广玉兰幼苗吸收系数进行相关性分析可知,土壤养分、吸收系数与土壤酶活性间均在相关关系。
由表2可知,碱解氮与全氮、全磷、脲酶、过氧化氢酶间均达到了极显著的正相关关系,与全钾呈显著的负相关关系;速效磷与磷酸酶的正相关关系极显著;速效钾与各指标存在相关关系,但关系不显著。N的吸收系数与过氧化氢酶呈极显著的负相关关系,与脲酶、多酚氧化酶呈显著的负相关关系;P的吸收系数与脲酶呈显著的负相关关系;K与脲酶和过氧化氢酶呈显著的正相关关系。说明土壤养分对其对应的土壤有效养分影响显著,土壤有效养分与其相应土壤酶活性存在显著影响,养分元素相应的酶活性越高,养分元素含量越高,其对应的有效养分也越高,但相应养分元素的植物吸收系数反而越低;因此,土壤酶活性影响了土壤养分及其有效养分的含量及其广玉兰幼苗养分吸收的变化。广玉兰幼苗的对氮素的吸收在过氧化氢酶、脲酶和多酚氧化酶活性最高的玄武岩(岩性1)的土壤条件下吸收利用效率较低,在石英砂岩(岩性2)及长石石英砂岩(岩性3)上较高;对磷素的吸收也是在脲酶活性最高的玄武岩(岩性1)的土壤条件下吸收利用效率较低,在长石石英砂岩(岩性3)较高;对钾素的吸收则在脲酶、过氧化氢酶最低的长石石英砂岩(岩性3)的土壤条件下吸收利用效率较低,在玄武岩(岩性1)上较高。
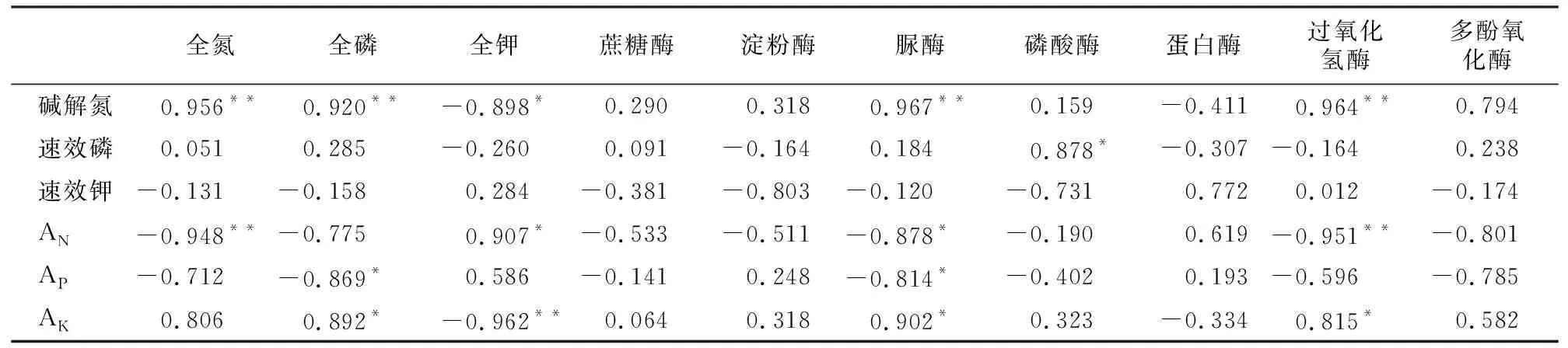
表2 不同岩性土壤养分、土壤酶及吸收系数的相关性
3 结论与讨论
喀斯特地区不同岩石类型的条件差异,使得其发育的土壤存在养分条件差异和提供有效养分元素的能力各有不同,且同时受到岩性控制,具体收到土壤酶活性的影响,使得养分的贮存转化和吸收利用各有不同,导致对其上的树木生长发育产生不同影响[24]。不同岩性土壤酶活性越高,则其养分含量越高,其上生长的广玉兰养分吸收越低,反之越高。
广玉兰幼苗在喀斯特石英砂岩(岩性2)、长石石英砂岩(岩性3)发育的土壤上对氮素和磷素的吸收利用效率较高,在玄武岩(岩性1)发育的土壤上对钾素的吸收利用最高,说明植物在高资源条件下水养的利用效率更高[25],这与韩愈等[26]研究土壤水分对喀斯特地枫皮幼苗生长影响的结论类似,即植物在干旱胁迫下渗透调节能力增强,更进一步揭示了广玉兰树种能通过提高养分利用效率的方式来达到耐瘠薄的基本适应机制。玄武岩(岩性1)发育的土壤属于粘壤土,石英砂岩(岩性2)、长石石英砂岩(岩性3)发育的土壤属砂质黏土[14],保水保肥的能力及土壤质量在玄武岩(岩性1)发育的土壤上表现最高[11],而广玉兰幼苗初期的生长效应在石英砂岩(岩性2)发育的土壤生长较快[14],说明喀斯特生境土壤质量的好坏影响了植物的生长条件,但不是决定性因素,植物对养分的吸收利用才是决定植物生长的重要指标。研究结果可为喀斯特地区广玉兰幼苗的培育提供参考,并为喀斯特地区的植被恢复和不同岩石类型区域的植被选择、配置和栽培提供科学参考和指导。
