持续干旱下金边黄杨的滞尘效应
2020-09-17林鑫涛
林鑫涛,张 晶,陈 健
浙江农林大学亚热带森林培育国家重点实验室, 杭州 311300
城市植物(例如道路绿化、屋顶花园、生命墙、城市森林公园、城市绿地等)能够有效吸附空气中的颗粒物[1-5],在减缓颗粒物污染上具有很大的潜力。树木覆盖率的增加有助于大气中悬浮颗粒物浓度的降低[6],进而改善空气质量。植物的种类、生活型、叶片形态、叶表的微结构等均会影响植物的滞尘能力[4, 7-13]。Song等[9]通过扫描电镜(Scanning electron microscope)观测到叶片的气孔是颗粒物沉积的最佳区域之一,气孔大小、密度、数量以及开度等均会影响植物表面的颗粒物滞留量[4, 10-11, 13]。Weerakkody等[3]发现蜡质含量丰富的植物,其表面吸附颗粒物的密度也较高。蜡质的化学成分、结构对颗粒物的滞留也具有重要作用[14-16]。
水分被认为是影响植物生长发育的重要限制性因子[17],城市的供水紧张使绿地植物长期处于干旱状态。水分的匮乏会迫使植物气孔活动减弱或关闭、蒸腾降低[18-19],也会使植物加大烷烃类物质、醛类物质的合成,进而增加蜡质含量以适应不利环境[20-21],这些形态结构的变化会减弱或增加植物叶片对颗粒物的吸附能力。此外,干旱会改变叶片表面的微环境而影响颗粒物的滞留效果,如植物蒸腾作用减弱后,局部湿度的降低会减弱叶表粘性而减小大粒子的沉降[22]。
目前干旱对植物滞尘的影响研究多集中在针叶植物[10, 15, 23-24]。针叶植物的枝条结构复杂,气孔排列紧密,气孔密度和表面粗糙度均高于阔叶树种,且其丰富的蜡质含量与油脂的分泌,使得针叶植物的滞尘能力通常被认为是强于阔叶植物的[11-12, 25-27]。然而针叶植物对高强度交通污染的耐受性较差[14],在南方城市的绿化中,其比例远低于阔叶植物,在机动车尾气污染严峻的长三角地区尤为明显。长期暴露在颗粒物污染中会导致植物形态特征的改变,最明显的现象就是叶片失绿[28-30],有限的供水亦会使叶片的脱色衰败[17, 19],然而干旱对颗粒物暴露下植物形态的研究鲜有报道,干旱下植物的滞尘机理尚不明晰。
本研究以金边黄杨(EuonymusjaponicusThunb.var.aurea-marginatusHort.)为对象,将机动车尾气注入开顶式气室内以模拟颗粒物污染,根据供水条件差异分成正常供水下的尾气暴露组(P组)与持续干旱下的尾气暴露组(PD组)。本研究旨在:(1)探究持续干旱对尾气暴露下的金边黄杨的形态影响;(2)对比金边黄杨在正常供水下与持续干旱下对不同粒径颗粒物的单位叶面积滞留重量的变化;(3)分析干旱对金边黄杨滞尘效应的影响。
1 材料和方法
1.1 实验材料
金边黄杨是大叶黄杨(EuonymusJaponicusL.)的一个变种,为卫矛科卫矛属的常绿阔叶灌木,是长三角地区常见的绿化植物,因其优美的彩色叶片而在园林配置中广受关注,对光强具有较低的耐受性,能适应较为荫蔽的环境[31]。本研究选用树高(均高0.8 m)和基径(平均3 cm)较一致的3年生苗木,于2017年3月栽植于直径30 cm,高28 cm的塑料盆中,基质配比为园土∶泥炭土∶蛭石=7∶7∶6,最大田间持水量约为50%。
1.2 实验地点
栽植好的苗木养于浙江农林大学平山实验基地(30°15′50.61″N; 119°42′54.51″E)的田间环境下约1年,基地周边以农村居住地为主,3 km范围内不存在高污染源。根据浙江省大气复合污染立体检测网络中的第四中学(与基地的直线距离约4.1 km)和市府大楼(与基地的直线距离约2.9 km)两站点在2013年—2016年间的大气污染物监测数据,实验区的PM2.5年均浓度约54.38 μg/m3,空气质量良好。
1.3 实验设计
金边黄杨的受控实验在修改后的开顶式气室(Open top chamber,OTC,以下简称为“气室”)内进行。气室由塑钢、无色钢化玻璃构成,主体高3.5 m,横截面为边长1.5 m的正八边形,顶部为45°锥形收口,高为1 m,并与大气相通。锥形收口顶上0.5 m处架空放置一块钢化玻璃以避免降水进入气室。将机动车尾气直接排入气室外的尾气收集装置中,后经鼓风系统将收集装置中的尾气通过管道注入气室内,注入时长为每日5 h,采取“早晚双高峰”模式以模拟城市污染特征,即早高峰:7:00—10:00;晚高峰:18:00—20:00。根据《环境空气质量指数(AQI)技术规定(试行)》(HJ 633—2012)中的空气污染指数分级标准,将空气质量控制为重度污染,即气室内PM2.5的24 h浓度控制在150—250 μg/m3,则排入尾气的PM2.5浓度控制在720—990 μg/m3。气室内的PM2.5浓度通过CPR-KA空气质量检测仪(北京康尔兴科技发展有限公司,中国)进行动态监测,温度通过DS1923系列iButton纽扣式温度记录仪(上海沃第森电子科技有限公司,中国)进行监测。
实验处理于2018年4月10日18:00开始,至2018年5月15日18:00结束。将尾气暴露气室内的金边黄杨随机分成2组,一组正常供水(P组),另一组持续干旱(PD组),每组设置4个重复,金边黄杨的处理说明见表1。实验前通过充分浇水使所有盆栽的土壤含水量达至饱和,后通过停止供水让PD组的盆栽自然消耗土壤水分,而P组的盆栽则在每日的土壤含水量测定后进行补水。土壤含水量使用便携式土壤水分仪TDR100(Spectrum Technologies, Inc.美国)在每日17:00左右测定并记录,测定频率为1次/d,以体积含水量(Soil volumetric water content,θv)表示。类似的持续干旱实验表明,当植株开始进入枯萎状态但不至于大面积死亡时的土壤体积含水量在2.0%—5.0%之间[15, 19, 23]。因而当PD组每日测得的θv低于5.0%时,对植株做20—100 mL的适当补水,使得土壤体积含水量不低于2.0%,以保证植株在试验周期内不出现大面积的死亡现象。P组植株的土壤体积含水量控制在37.5%—40.0%。

表 1 金边黄杨的处理说明
实验处理前,使用去离子水冲洗所有植株的叶片以去除叶表原先滞留的颗粒物。有研究表明润湿的叶片比干燥时具有更大的粘性,会加速颗粒物在叶表的沉降,并减弱颗粒物的再悬浮作用[22, 26]。为避免叶片清洗对实验期间植株滞尘速率的影响,将叶片的清洗工作安排在尾气处理前一天,以待植株叶片充分干燥。多数学者认为降水或简单的清洗不能完全去除叶片表面原先滞留的颗粒物[7, 26, 32],因而在尾气处理前(4月10日17:00)采集植株叶片做初始滞尘测定,之后每隔7天采集一次叶片用以植株滞尘量的测定。
1.4 叶片采集
在每株植株的“东”、“南”、“西”、“北”四方位随机采取4—5片叶片,混合后装在干净的封口袋内,保存于冰箱保鲜室直至样本滞尘量的测定。实验期间共采集6批样本(含本底采集),每批均为4包(重复)。为确保得到足够的细颗粒物量,同时避免过多采样导致大颗粒物堵塞滤纸滤孔,且不影响植株的正常生长[33],每包叶片的采集面积控制在200 cm2左右。
1.5 滞尘测定
叶片表面滞留的颗粒物采用洗脱称量法测定[8, 33-34]:将样本置于500 mL烧杯中,加入250 mL蒸馏水后使用超声波清洗器清洗4 min,使叶表滞留的颗粒物充分脱离叶片,后取出叶片并使用蒸馏水进一步冲洗。对上述两部分得到的浸洗液与冲洗液用标准分样筛(160目,孔径约100 μm)过滤后依次使用已烘干称重的孔径为10 μm(Whatman Type 91,英国)、2.5 μm(Whatman Type 42,英国)和0.2 μm(Whatman PTFE滤膜,英国)的滤纸(膜)在直径47 mm的抽滤装置中进行共计3次的抽滤,从而得到三个粒径范围的颗粒物:大颗粒物(10—100 μm),粗颗粒物(2.5—10 μm),细颗粒物(0.2—2.5 μm)。抽滤前后均将滤纸置于称量瓶中并在60℃的烘箱中烘干1 h后置于干燥皿中冷却24 h,从而使滤纸、称量瓶与干燥皿内的湿度保持一致,之后对含有滤纸的称量瓶用万分之一的分析天平称重,抽滤前后两者差值即为样本吸附的各粒径颗粒物的重量Wi(g)。
将上述浸洗后的叶片晾干/擦干,之后平整置于扫描仪上(Epson Perfection V370 Photo,爱普生(中国)有限公司,中国)进行扫描,得到的数码图像导入Image J1.46r(National Institute of Health,美国)中进行叶面积S(cm2)的计算,各样本不同粒径颗粒物的单位叶面积的滞留重量(以下简称“单位叶面积滞尘量”)即为Wi与S的比值(μg/cm2)。
1.6 数据分析
单位叶面积滞尘量在各采样时间节点上的差异使用单因素方差分析(ANOVA)进行检验,之后采用LSD进行两两比较。方差分析前需检验数据的正态性分布(Shapiro-Wilk检验,P>0.05)和方差齐性(Levene′s检验,P>0.05),若数据不满足正态性分布或方差齐性时,则在分析前采取自然对数转换。干旱与正常供水下滞尘量的比较采用独立样本t检验进行分析。所有检验分析的显著水平为0.05,数据以“均值±标准误”的形式表示。数据的统计分析均在SPSS 25.0(IBM,美国)中完成,图件在Origin 2018(OriginLab,美国)中绘图。
2 结果
2.1 气室环境分析
临安自动气象观测站(区站号:58448:30°14′N; 119°41′E)记录的历史数据(国家气象科学数据共享服务平台,http://data.cma.cn/site/index.html)表明,实验基地在近5年内同期的日均温度约18.6℃。iButton温度记录仪显示,实验期间,气室内的日均温度约25.1℃,日最高温度可达32.7℃,多出现于10—14时,表明气室内的温度较适宜植物生长[35]而又不对其产生过度的高温胁迫。
空气质量检测仪显示(图1),气室内注入的尾气中的PM2.5时均浓度为(789.94±132.37)μg/m3,日均注入时间为5 h,气室内PM2.5的24 h平均浓度为214.64 μg/m3,对应的空气质量指数级别为五级(IAQI=265,HJ 633—2012),属于重度污染,尾气处理符合实验设计需求。

图1 实验期间气室中PM2.5浓度时间序列图
盆栽的土壤体积含水量于每日17时测量并记录(图2),后计算P组植株所需的补水量并做补水措施,从而使P组植株的θv保持在37.5%以上。PD组的起始θv约为36.6%(日常补水前),在持续干旱的前10日,θv呈明显的下降趋势,平均每日降低约2.76%。第10日后(4月20日)θv降低至10%以下,4月25日θv降低至5%以下,之后开始对PD组植株做适当补水措施,使PD组的θv维持在2%—5%之间,直至实验结束。
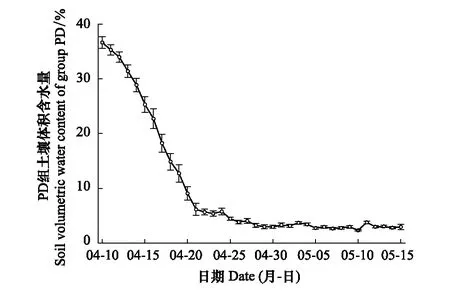
图2 实验期间PD组土壤体积含水量时间序列图
2.2 金边黄杨的形态变化
实验期间,金边黄杨的形态特征变化见表2。P组金边黄杨的部分老叶在第19天开始黄化并脱落,至5月15日,大部分植株完成老叶的脱落,小部分个体的幼叶有所枯黄,除此外,未发现其他明显变化。PD组的金边黄杨在第8天便出现叶片干枯现象,此时θv为14.86%±1.53%,2天后,老叶开始脱落,而到第19天,PD组的成熟叶甚至幼叶均有明显的黄化、脱落现象,到5月8日,大部分植株的叶片枯黄(θv<5%),受外力刺激时易脱落,在实验结束后,大部分植株进入休眠状态,仅尖端残留少量干枯叶片(枯绿)。
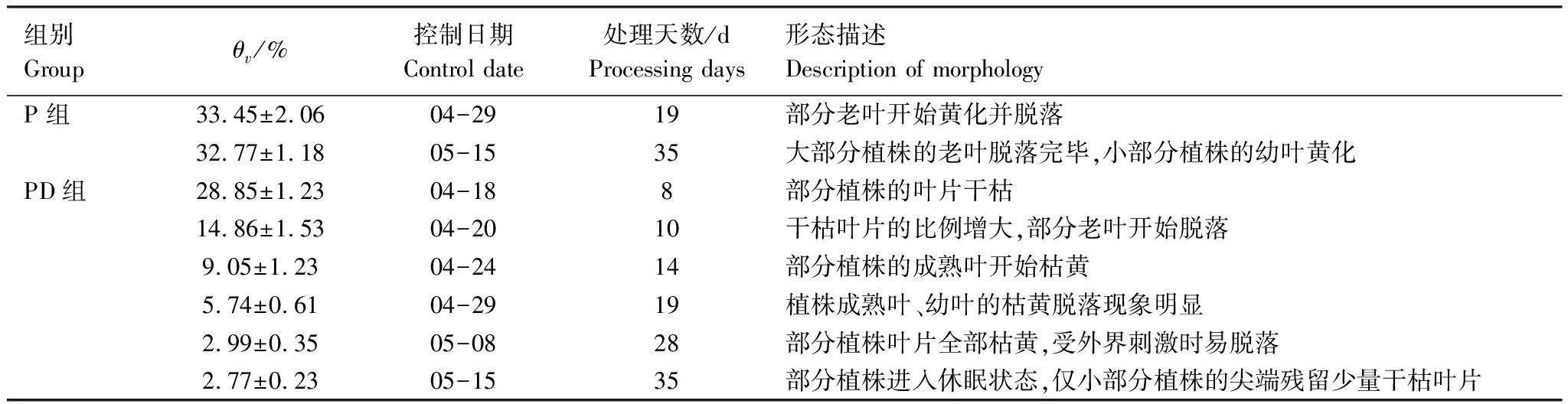
表2 实验期内金边黄杨的形态特征
2.3 金边黄杨对不同粒径颗粒物的滞留动态变化
金边黄杨的初始滞尘量(本底滞尘)为(30.27±2.44)μg/cm2,P组与PD组间的初始滞尘差异不显著(t=-0.377,Pns=0.719),其中大颗粒物的初始滞留量为(16.21±1.80)μg/cm2,粗颗粒物为(9.16±1.07)μg/cm2,而细颗粒物为(4.75±0.66)μg/cm2(图3)。正常供水时,P组植株对不同粒径颗粒物的滞留量在前14天增速较大,但相邻两采样节点上的滞尘量差异不显著,大颗粒物和细颗粒物均在第35天达到极大值52.41 μg/cm2和9.98 μg/cm2,较本底净增长约38.31 μg/cm2和5.49 μg/cm2,粗颗粒物在第28天较本底净增长了98.10%,之后略有降低;持续干旱时,PD组植株滞留的三种颗粒物在前28天均随时间的推移而增大或波动,之后,大颗粒物与粗颗粒物有显著降低,大颗粒物由极值的45.89 μg/cm2降低到28.02 μg/cm2,而粗颗粒物则由极值的18.39 μg/cm2降低了53.1%,两种颗粒物在第35天的滞留量与初始值的差异不显著。细颗粒物在实验期间的变化弱于另两种颗粒物,其滞留量在各采样节点间的差异不显著,极大值出现在第14天,之后开始小幅下降,至实验结束,仅减少了0.89 μg/cm2,但仍比初始滞尘量高约1.13 μg/cm2。
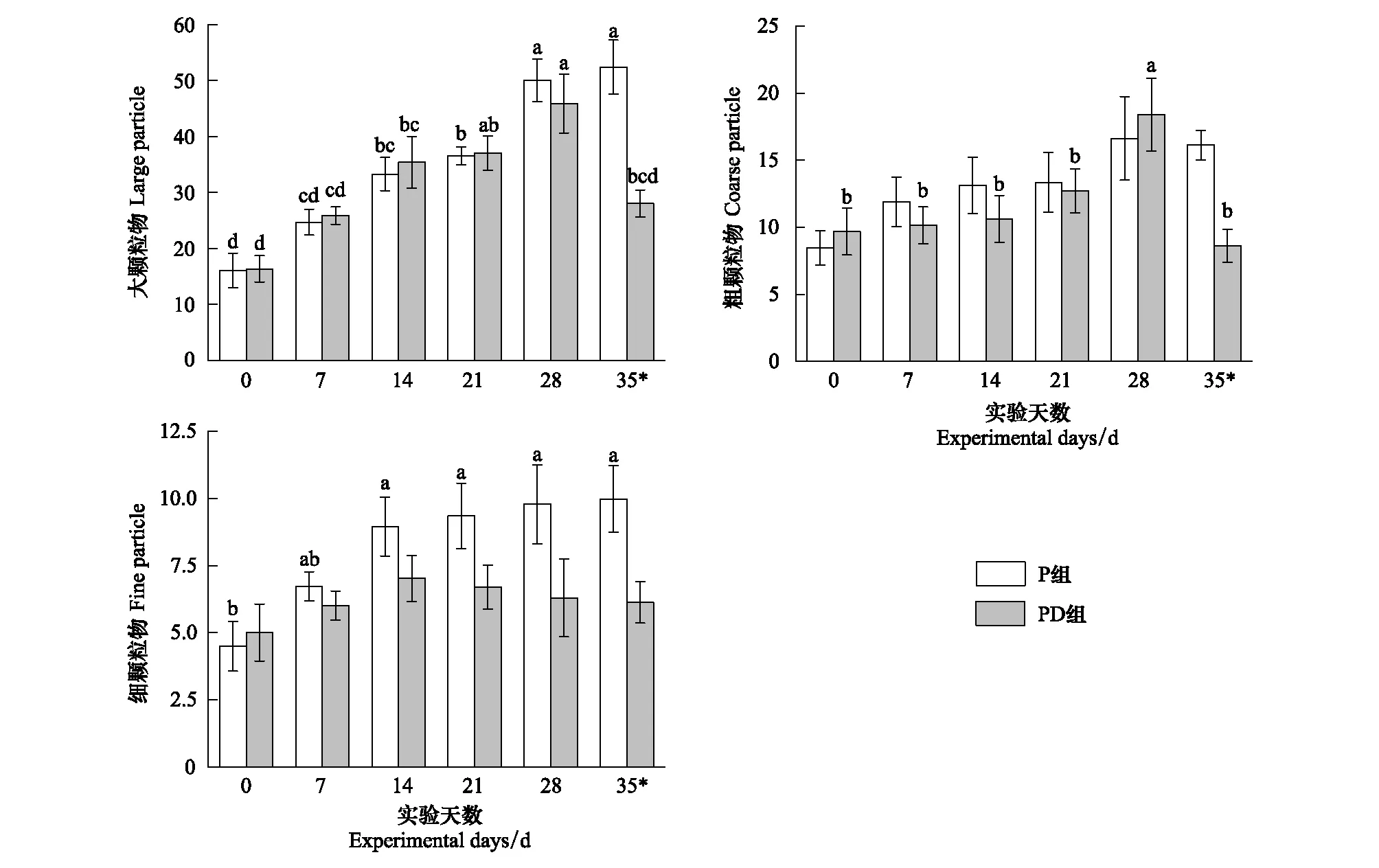
图3 不同粒径颗粒物的单位叶面积滞留量/(μg/cm2)
2.4 金边黄杨滞留颗粒物的粒径构成变化
实验期间,金边黄杨叶片表面滞留颗粒物的粒径构成(贡献率)见图4,其中以大颗粒占主要优势(57.0%—66.8%),粗颗粒其次(20.1%—27.5%),细颗粒做最小贡献(8.9%—16.2%)。正常供水时,P组植株叶表上大颗粒的贡献率平均每7日增加2.3%,细颗粒的贡献率在前21天相对稳定,之后下降了约3.0%,粗颗粒以每周平均1.7%的降幅持续下降;持续干旱时,PD组植株叶片上的颗粒物粒径构成在前14天变化明显,主要表现为大颗粒大幅上升而粗颗粒急剧下降。之后,大颗粒每周的变幅在1.1%内,而细颗粒在第5周的贡献率急剧升高了约5.4%。

图4 叶表滞留颗粒物的粒径构成
2.5 持续干旱对金边黄杨滞尘能力的分析
实验期间,P组金边黄杨的平均滞尘量(总颗粒物)为56.98 μg/cm2(图5),高于PD组7.93 μg/cm2,独立样本t检验表明P组与PD组在实验期间的平均滞尘量无显著差异(t=1.610,P=0.114)。各时间节点的t检验表明,持续干旱仅在第35天极显著的降低了植株的总滞尘量约45.53%(t=7.456,P<<0.01),而在前28天,虽然植株的滞尘量总体上表现为P组>PD组,但该时期内未发现两组间的滞尘量有显著性差异。当以净单位叶面积总滞尘量(去除本底后的滞尘量)评估持续干旱对植株滞尘量的影响时(图5),P组在实验期间的平均滞尘量比PD组高56.05%且达到显著水平(t=2.400,P=0.021),但两组在第7—28天的平均净滞尘量上未发现显著性差异(t=0.087,P=0.382)。在各时间节点上,持续干旱仍仅在第35天显著降低了金边黄杨的净滞尘量(t=6.759,P≤0.01)。
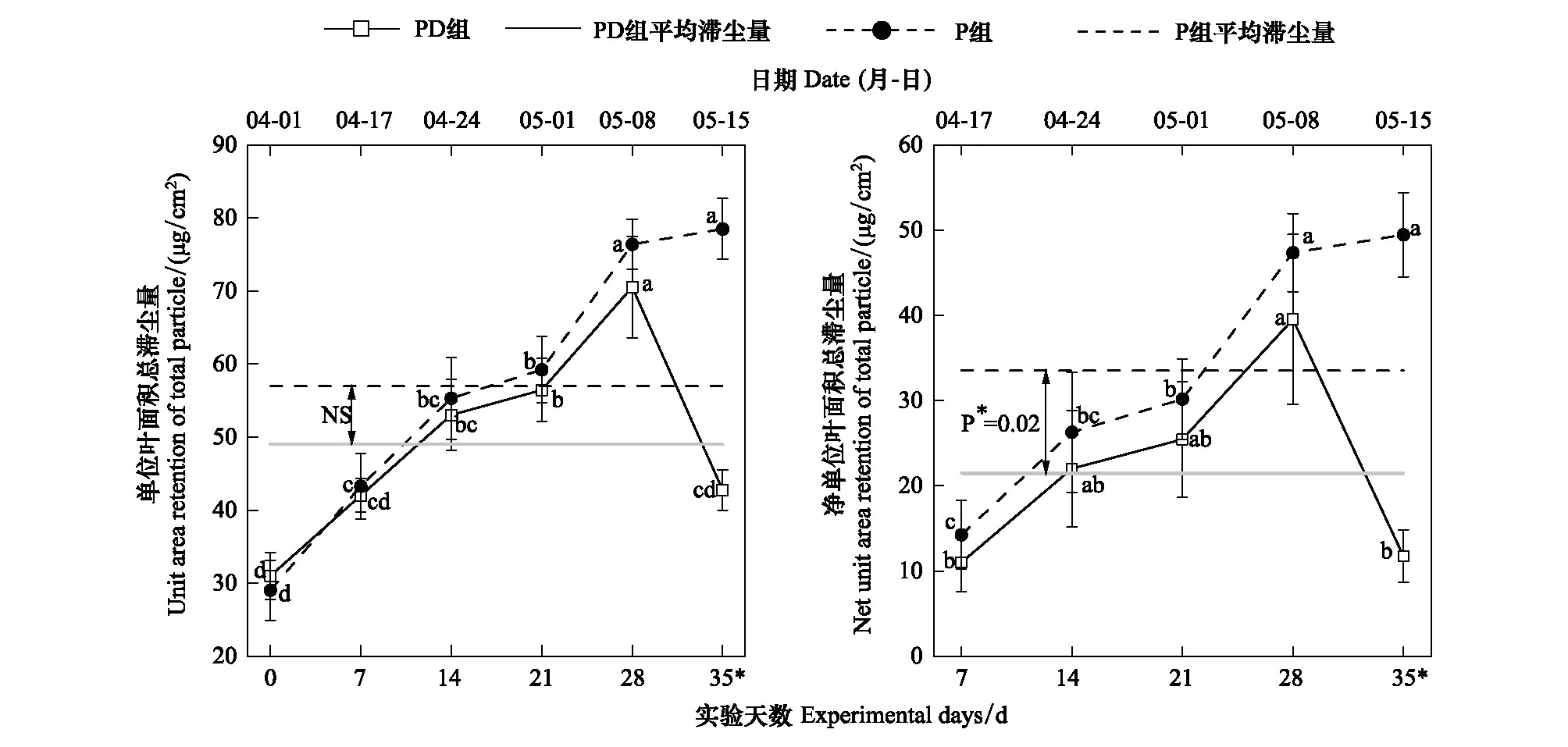
图5 干旱对单位叶面积总滞尘量的影响
3 讨论
3.1 金边黄杨的形态变化
P组植株在实验期间的形态变化表明,正常供水时,尾气暴露35天对金边黄杨成熟叶、幼叶的形态影响较小,而老叶则在第19天开始黄化、脱落,暗示此时尾气对植物细胞产生了毒害作用。暴露初期,叶片滞留的颗粒物或因堵塞气孔[9],干扰气体交换,使叶表温度增高[30],或因“遮荫效应”[36],而导致叶绿素合成受阻;尾气中的某些成分也会促使叶绿素降解或抑制叶绿素合成酶的活性[2, 36],或因活性氧(Reactive Oxygen Species,ROS)的过量累积[28]而导致叶绿素的含量降低,进而叶片失绿黄化。当尾气对细胞的毒害作用超过其耐受性极限时,便发生叶片的脱落现象,但该现象仅在老叶上发生则可能是因为老叶对尾气的抗性较弱,或因老叶表面更高的颗粒物滞留量[15-16, 37]而加大了其损伤。此外,植物也可能通过将尾气中吸收的有害物质富集到老叶中来减小尾气对成熟叶与幼叶的毒害。
PD组植株的老叶黄化脱落比P组提前了9d,部分植株在处理35d后进入休眠状态(去除胁迫后有复苏现象),成熟叶与幼叶几乎脱落完毕,这暗示着干旱与尾气可能存在正交互作用而加重了植物体的损伤。Hanslin等[23]与Subhash等[38]均表示水分对植物的负面影响强于尾气,干旱则因为对气体交换的强烈干扰[18-19]或加快了ROS的积累[39]而使尾气对植物的伤害加快,而尾气则可能因破坏叶片蜡质结构、形状,甚至使其降解或退化[2, 15, 30, 40]而降低了植株对干旱的耐受性[29, 39]。因此,持续干旱与尾气共同作用对金边黄杨的破坏明显高于单独的尾气暴露。
3.2 持续干旱下金边黄杨的滞尘效应
大气颗粒物沉降到植物表面的过程与其粒径大小密切相关[22, 37]。P组的金边黄杨对各粒径颗粒物的滞留量随时间推移而增大或在波动中达到动态平衡,而PD组的金边黄杨在第35天对大、粗颗粒物的滞留量显著降低了近38.9%和53.1%(图3、图5),主要原因有以下两方面:(1)PD组植株的叶片在第28天已基本枯黄,之后叶片纷纷脱落(表2),第35天时仅在植株尖端残留干枯幼叶,迫使最后一次的采样对象由原先的成熟叶改至幼叶。成熟叶更加粗糙的叶片表面可能使其对颗粒物的捕获能力高于幼叶[15-16, 37]。(2)极度干旱下,PD组植株的叶片含水量低下,实验28天后的叶片在外界刺激时易脱落(图2、表2),这暗示着在叶片采集、土壤含水量测量等实验过程对植株叶片的抖动强度会相对增大,以及在气室内走动时带来的软风也会使部分滞留在表面的颗粒物重新悬浮至大气中[22],虽然该风速对颗粒物的再悬浮作用有限[26, 32]且实验中已尽量避免,但PD组植株在胁迫后,叶片表面的润湿性和粘性的降低可能减弱其对颗粒物的滞留[13],振动或软风对颗粒物的再悬浮作用会有一定的加强。此外,叶片大量脱落后会减弱植株内的气体湍流,以此减少了粗颗粒物对植株表面的撞击机会[22, 26],从而造成了粗颗粒物更高的降低率。
细颗粒物主要滞留在叶片的气孔周边或密毛覆盖区[37, 41],雨水对其冲刷作用或风力对其再悬浮作用也弱于大、粗颗粒物[7, 42],其滞留程度也是三种粒径中最高的。细颗粒物体积小、质量轻,且在水汽的作用下易聚集成大粒子[22, 26],因而对滞尘重量的反应较弱,无论是正常供水或持续干旱时均是如此(图3)。Räsänen等[10]表示阔叶植物较强的蒸腾作用会降低叶片的表面温度,而热泳现象会增加细颗粒物向叶表迁移,较高的气孔活动也会使颗粒物(尤其是盐粒子)更容易潮解[40]而增加其沉降速率。在持续干旱14d后,PD组的土壤体积含水量基本低于10%,此时植株的蒸腾速率与气孔导度可能已大幅下降[19],伴随的是蒸腾作用与气孔活动的减弱以及细粒子沉降的减缓,因而细颗粒物的滞留量不再明显升高而在波动中变化。对于PD组在第35天的细颗粒物滞留量变化不明显,我们猜测可能是因为振动或风力对细粒子的再悬浮作用较弱[26],且尾气中高浓度的细颗粒物使其在叶表的沉降多于再悬浮,而此时的幼叶表面的气孔与密毛覆盖区的发育程度可能较高,对细颗粒物具有良好的滞留效果。
植物表面滞留颗粒物的粒径构成主要受到植物叶片特征和污染源的影响[12, 34]。在金边黄杨的粒径构成上,大颗粒物占据50%以上,细颗粒物在20%以下(图4),这与道路旁的植物表面的粒径构成是相似的[8, 36]。各粒径颗粒物的贡献率变化可能是由沉降速率的差异造成的,沉降速率则与颗粒物的性质与沉降形式相关。细颗粒物主要以布朗扩散或随气流运动时被植物截获而沉降,粗颗粒物的沉降以惯性碰撞为主,而大颗粒物主要在重力作用下沉降[1, 22]。通常大颗粒物的沉降速率最高,细颗粒物最低,这解释了P组中大颗粒的贡献率逐渐增长的趋势。细颗粒物在前21d保持较高的贡献率可能是因为尾气中高浓度的细粒子加大了植株对其捕获效率[36],之后植株对细粒子的滞留基本达到饱和(21d后植株对细颗粒物的滞留量变化差异不显著,图3),但大颗粒物仍保持较高的滞留速率,因而产生了P组植株各粒径颗粒物的贡献率变化。在持续干旱下,PD组植株的气孔活动减弱及叶片微形态的变化均会影响叶片对颗粒物的滞留效率,其中以细颗粒物受到的影响最为明显(图3)。因此,在相同的污染源下,PD组植株的粒径构成变化异于P组。两组在各时间节点的粒径构成上,以第35天的差异最小,这可能是因为此时植株对各粒径颗粒物的滞留达到饱和,污染源对粒径构成的影响强于叶表微形态的差异。
在单位叶面积的滞尘重量上,持续干旱使金边黄杨对各粒径颗粒物的平均滞留量减少2—3 μg/cm2,但两组植株在前28d的统计学差异不显著。Hanslin等[23]发现中度干旱能增加欧洲赤松(PinussylvestrisL.)幼苗的滞尘重量,Räsänen等[10, 15, 24]则发现重度干旱会减少毛桦(BetulapubescensEhrh.)的颗粒物捕捉效率,而中轻度干旱对欧洲云杉(Piceaabies(L.)Karst.)与欧洲赤松的作用则是相反的。与本研究相似的是:上述研究均未发现干旱对植物滞尘的显著影响。
在第35天,PD组金边黄杨的滞尘量极显著低于P组,其原因与PD组滞尘量在第35天的急剧下降类似,将采样对象改为幼叶后,虽然使两组在该节点的滞尘量比较失去了意义,但这也是持续干旱与尾气暴露共同作用下,植株形态变化造成的必然结果——成熟叶基本脱落完毕,此时植株的滞尘能力明显减弱。受限于实验材料,本研究只在单位叶面积的尺度上分析持续干旱对植物滞尘重量的影响,而实际应用中则更为关注在单木尺度或群落尺度上的滞尘效应,其中单木尺度的滞尘量可简单表征为总叶面积与单位叶面积滞尘量的乘积[9, 33]。持续干旱下金边黄杨总叶面积的大幅降低明显会减弱植株在单木尺度上的滞尘量,以此将降低其滞尘效应,而受限的实验数据无法定量分析金边黄杨的单木滞尘量与干旱胁迫间的关系。
4 结论
本研究在开顶式气室(OTC)中以机动车尾气模拟颗粒物污染,对不同供水条件下的金边黄杨的形态与颗粒物滞留量的动态的观测分析表明:(1)正常供水时的尾气暴露(P组)使金边黄杨在第19天出现老叶黄化、脱落现象,而持续干旱下的尾气暴露(PD组)则比P组提前9d显现该现象,之后在第19天促使植株的成熟叶与幼叶脱落。(2)P组植株在实验期间对大颗粒物(10—100 μm)、粗颗粒物(2.5—10 μm)和细颗粒物(0.2—2.5 μm)的平均滞留量分别为(34.76±2.92)μg/cm2、(13.33±0.91)μg/cm2、(8.21±0.58)μg/cm2,而持续干旱使PD组的植株对各粒径颗粒物的平均滞留量减少2—3 μg/cm2。然而,在前28d未发现干旱对滞尘量的显著性影响,而在第35天,干旱则显著降低了各粒径颗粒物的滞留量约38.60%—46.54%。可见,短期的中轻度干旱对金边黄杨滞尘能力的影响较小,而极度干旱则促使尾气暴露下植株成熟叶的大面积脱落,并造成其滞尘能力大幅度降低,从而减弱了金边黄杨的滞尘效应。
致谢:李铭、黄垌茗、舒达、叶璞对实验处理给予帮助,林权虹、陆婷、钱瀚睿、黄菡菡等对室内指标分析给予帮助。
