中国西南地区热带森林演替序列碳动态
2020-09-17步巧利谭正洪张一平
步巧利,谭正洪,张一平
1 广东省佛山市气象局, 佛山 528000 2 中国科学院热带森林生态学重点实验室, 昆明 650223
热带森林因其复杂的群落结构,多样的物种和巨大的生物量而著称。在热带森林研究享有盛誉的生态学家Kira Tatuo将热带森林生态学的研究喻为生物学研究的“临界状态”[1]。热带森林在全球碳平衡和碳循环中也扮演着十分重要的角色。虽然热带森林只占有约22%的潜在陆地植被分布面积[2],却拥有着全世界陆地生产力的43%[3]和碳储量的59%[4]。
在地球系统碳循环的研究中,热带地区和热带森林也是最不确定的[5]。无论是使用涡度相关技术,生物调查和生理生态尺度扩展,还是使用大气CO2浓度反演,进行热带雨林碳平衡的评测都是一个挑战[6]。因此,在回答全球碳循环“失汇”的问题上,热带森林也有着显著的重要地位。
随着人口的增长,社会的发展和经济的膨胀,原生森林在不断的减少,取而代之的是经济价值较高的人工林,如橡胶林。同时,一部分破坏后遗弃的林地,次生林随着时间生长起来。关于次生林演替过程中的碳储量和碳平衡的变化却鲜为人知,特别是在热带地区。因此针对中国西南地区热带森林演替碳动态的研究非常有必要,且可为全球碳循环的“失汇”问题寻找答案。
本文利用3个热带次生林的样地的数据,探讨:(1)处于不同演替状态的热带次生林的碳储量状况如何;(2)森林的净碳蓄积,死亡碳损失和更新碳增长等碳动态如何;(3)次生林在未来的固碳方面的潜力如何。
1 研究方法
1.1 研究地概况
研究样地位于云南省西双版纳傣族自治州勐腊县勐仑镇(101°16′E, 21°55′N)。受大气环流、地理位置及地方气候的影响,地处热带北缘的西双版纳1年中有雾凉季(11—翌年2月)、干热季(3—4月)和雨季(5—10月)之分。该地的年平均温度为21.5℃,年平均降雨量为1492 mm。地带性植被类型为热带季节雨林和季雨林,在海拔较高(800—1800 m)的山地分布有南亚热带季风常绿阔叶林, 在部分中山的局部地段分布有山地雨林。该地区是我国热带雨林集中分布的重要区域,同时也是东南亚热带雨林分布的最北缘。由于地处古热带植物区系向泛北极植物区系的过渡区及东亚植物区系向喜马拉雅植物区系的过渡区,该区的生物区系成分十分复杂、物种多样性高度富集。由于特殊的地形地貌,气候的立体分异相当大,其东、北、西三面高,在一定程度上阻挡了西北方来的冷气流,使得最冷月平均温度并不低,弥补了积温的不足。在冬季低山沟谷及低丘上的浓雾又弥补了降水的不足。因而在该地区的低山沟谷及低丘上,形成了热带湿润气候,具有热带雨林发育的条件。
1.2 调查样地和样地调查
本文选择了西双版纳热带森林生态站调查样地中关于次生林的三个样地用于本研究。样地的基本情况见表1。
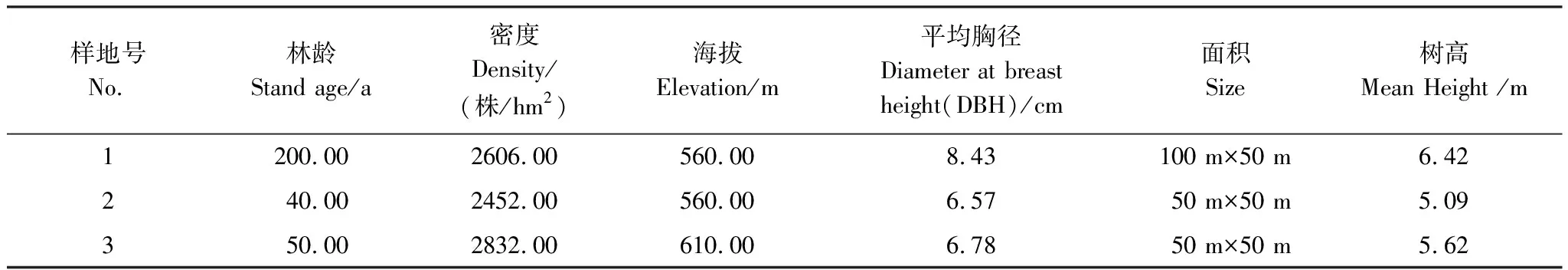
表1 三块研究样地的概况
样地的每木调查按照CERN(中国生态系统研究网络,Chinese Ecosystem Research Network)规定完成,利用每木调查法观测胸径、高度,1次/5 a(人工林或幼龄次生林1次/2 a或1次3 a,轮换观测),按Ⅱ级样方统计密度,同时在调查样地内安置收集框来确定凋落物量,同时在调查样地内安置收集框来确定凋落物量,具体调查细节请登陆CERN网站,参考西双版纳生态站链接(http://bnf.cern.ac.cn/)。样地的具体情况如下:
样地(1)是原生热带雨林样地,位于西双版纳热带植物园的迁地保护区内,该样地的植被受到一定程度的干扰(择伐),以绒毛番龙眼(Pometiatomentosa)、箭毒木(Antiaristoxicaria)、葱臭木(Dysoxylumexcelsum)、梭果玉蕊(Barringtoniafusicarpa)、窄序崖豆树(Millettialeptobotrya)等占优势的热带季节雨林。群落盖度为90%,群落的垂直结构可分为乔木层、灌木层和草本层,其中乔木层又可分为2个亚层(乔木I、II亚层)。乔木I亚层(乔木上层)高约20—25 m,主要由绒毛番龙眼(Pometiatomentosa)、箭毒木(Antiaristoxicaria)、勐仑翅子树(Pterospermummenglunense)等组成;乔木II亚层(乔木下层)高约5—20 m,主要由葱臭木(Dysoxylumexcelsum)、窄序崖豆树(Millettialeptobotrya)、小叶红光树(Knemaglobularia)、黑皮柿(Diospyrosnigrocortex)等组成;灌木层高约1—3.5 m,主要由大花哥纳香(Goniothalamusgriffithii)、假海桐(PittosporopsisCraib)、山木患(Harpulliacupanioides)等组成;草本层主要由长叶实蕨(Bolbitesheteroclita)、小叶楼梯草(Elatostemaparvum)、假斜叶榕(Ficussubulata)等组成,层间附(寄)生植物丰富。
样地(2)是热带季节雨林破坏后形成的以窄序崖豆树为优势树种的次生林。群落盖度较高,达到90%,群落高度为10—15 m,群落的垂直结构可分为乔木层、灌木层和草本层;乔木层主要由窄序崖豆树(Millettialeptobotrya)、披针叶楠(Phoebelanceolata)、短药蒲桃(Syzygiumbrachyantherum)、椴叶山麻杆(Alchorneatiliaefolia)等组成;灌木层主要由滇南九节(Psychotriahenryi)、弯管花(Chasaliscurviflora)等组成;草本层主要由南山花(Prismatomerisconnata)、马唐(Digitariasanguinalis)等组成。
样地(3)是热带季节雨林破坏后发育起来的次生林。群落盖度达到80%,群落高度为10—12 m,群落的垂直结构可分为乔木层、灌木层和草本层,其中乔木层主要由清香木姜子(Litseaeuosma)、伞花木姜子(Litsea umbellata)、黄丹木姜子(Litseaelongata)、印度栲(Castanopsisindica)、鹅掌柴(Scheffleraoctophylla)、披针叶楠(PhoebeNees)、短药蒲桃(Syzygiumbrachyantherum)、滇银柴(Aporusayunnanensis)等组成;灌木层主要由大花哥纳香(Goniothalamusgriffithii)、粗毛榕(Ficushirta)等组成;草本层主要由滇南素馨(Jasminumwangii)、铜锤玉带草(Pratia nummularia)、南山花(Prismatomerisconnata)等组成。
1.3 数据处理
按照已经建立的异速生长方程[7-8]计算群落胸径大于2 cm林木的生物量。已有研究表明,群落的生物量主要集中在胸径大于2 cm的林木。因此本文以大于2 cm的林木的总生物量来代替整个群落的生物量。使用普遍的0.5为转换系数,将生物量转化为碳储量[6]。
使用Sigmoidal函数拟合演替年份与群落的碳储量。部分文献针对群落碳动态中的几个主要参量做了说明[9-10]。森林演替年龄的确定如下:其中2,3,4,5年4个林龄的数据引自冯至立等[8];5,10,14,22年4个林龄的数据引自唐建维等[7];本研究的样地1,2和3的林龄根据查阅文献和资料记载[6,11-13],并结合群落的物种组成和胸径频度分布,将样地1、2和3的林龄分别定为200年,40年和50年。
2 结果与讨论
在森林的演替过程中,森林的胸径分布频度从近正态分布逐渐向小径级的偏态分布发展(图1)。也就是说,随着演替的进展,小径级林木所占的比例越来越高。
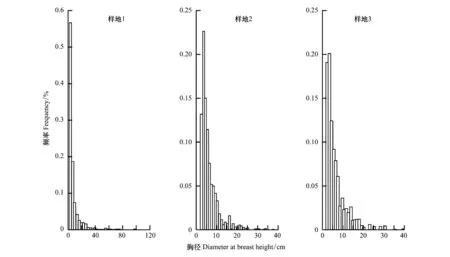
图1 调查样地内不同胸径的林木的频度分布(胸径大于2 cm)
2.1 处于不同演替状态的热带森林的碳储量
不同演替阶段西双版纳热带森林的碳储量如图2所示。整个演替过程可以被划分为5个阶段:干扰发生(I);干扰后的迅速生长期(II);较慢的生长期(III);储量近稳定期(IV);下一次干扰发生(V)。
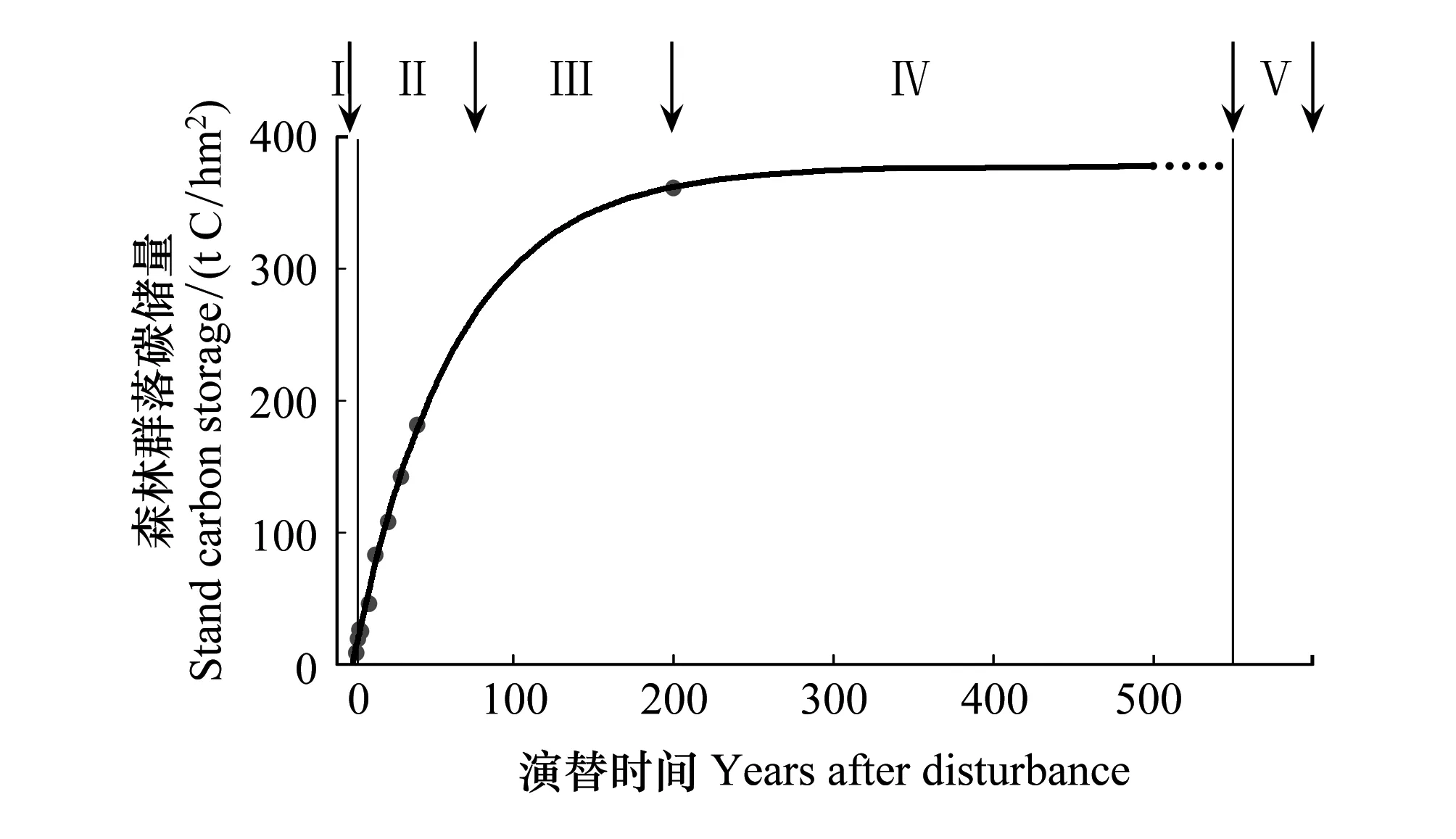
图2 图解演替过程中森林的碳动态
年轻的森林(Young forest)固碳能力最强是一个较为普遍的观点。如图2,西双版纳热带森林阶段II的固碳能力明显强于其他各阶段的固碳能力。我们希望在此强调的是不论在森林演替的哪个阶段,大树(不同演替阶段关于大树没有绝对的定义,对于原生热带雨林而言,大树通常被定义为胸径大于70 cm的林木)在森林固碳方面都占有非常重要的地位。
对于同一片森林,根据植物的生长规律,随着胸径的增长,胸径的相对增量越小(图3)。然而,从胸径绝对增量来看,胸径大的林木却发挥着主导性的地位。生物量与胸径之间存在较好的幂函数关系,方程表达式分别为y=0.6x1.14,R2=0.58;y=0.5x1.09,R2=0.525(图3)。因此胸径的绝对增量对生物量的贡献按照幂次放大。因此,大树在影响森林碳蓄积方面有着非常重要的地位。
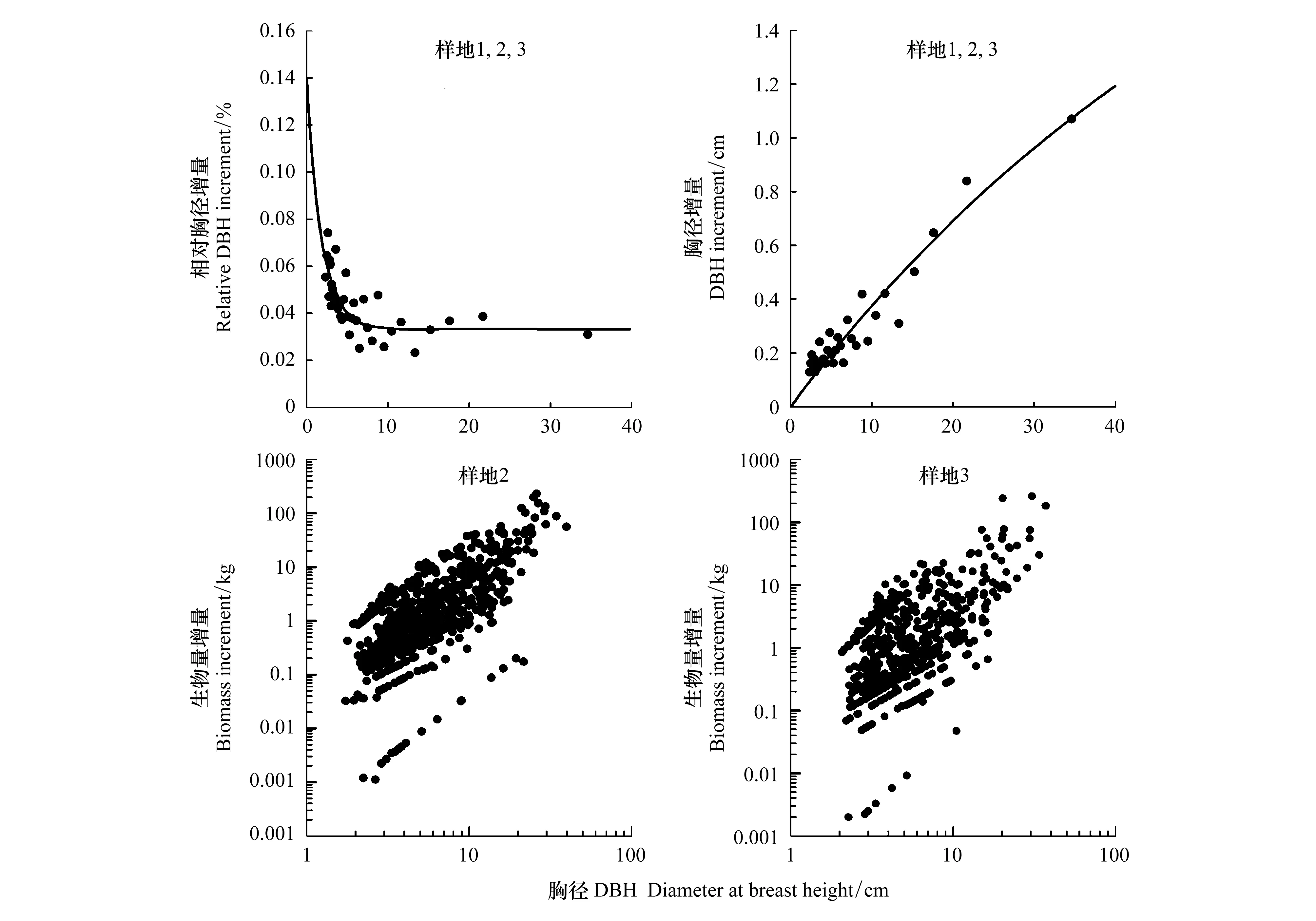
图3 不同胸径林木的相对胸径增长,胸径绝对增长,生物量增量
关于森林演替末期的碳平衡状态是至今争论较大的一个研究热点。根据Gorden的预测[14],在森林自然演替的末期(称老年林或过熟林(old-growth forest)),森林处于光合碳吸收和呼吸碳释放的均等状态,生态系统既不吸收碳亦不释放碳。然而,实测数据却并不支持该预测[15-16]。以西双版纳热带雨林为例,按照Odum的预测,当生态系统位于IV演替阶段时,生态系统的碳储量达到最高值,植被的碳蓄积过程几近停止;然而,在对一块未经明显人为干扰的永久研究样地的数据进行分析时,我们发现因为森林内林木的生长,森林每年以3.50 tC/m2的速度蓄积着碳[6]。根据全世界范围内广泛收集的数据建立的数据库,Luysseart等[16]也得出世界上多数森林并没有达到生物量的极限最大值。Luysseart等[16]认为,关于Odum的预测和实测数据之间的不吻合,最重要的原因在于老年林(old-growth)的定义。自然界的不同尺度的干扰是普遍存在的,森林并不能隔绝于干扰之外存在。小的干扰,如森林林窗的形成,将大大的改变森林内部的光环境,进而影响辐射,能量,水分平衡等生物物理过程,最终波及森林的碳动态。大的干扰,如火灾和砍伐,将导致森林的次生演替,对森林的碳动态产生不可逆转的改变。因而,我们认为的老年林并非Odum理想状态下的生态系统。
2.2 森林净碳蓄积,死亡碳损失和更新碳增长等碳动态
使用生物调查的数据来探讨森林生态系统的净生物量增量已经比较成熟:如单木跟踪方法(track individual method)[17-18]。然而,多数的研究只提供一个样地净生物量增量的结果,很少有编制生物量动态表。不论使用生物量来计算生态系统的净初级生产力还是净生物量增量,编制森林碳动态表格是必要的。从西双版纳热带次生林的生物量动态表格(表2,3),我们可以看出研究样地3中导致生物量损失的重要因素就是大径级(>60 cm)的林木发生了死亡,样地3中导致生物量损失的重要因素经查阅大量资料和咨询相关研究人员后发现造成该死亡的主要原因则是干扰。
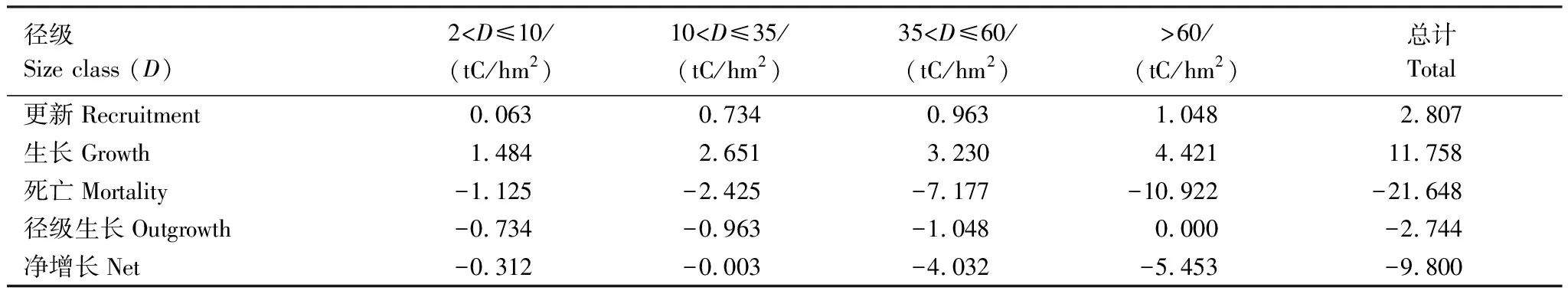
表2 样点2乔木生物量在2003—2007年之间的变化
森林凋落物量是按照生物调查法计算生态系统净初级生产力和探讨森林碳动态的另一个重要方面。与蓄积在林木中周转时间长的碳相比,凋落物可以在更小的一个时间尺度内影响碳动态。从2003—2007年样地2和3的凋落物动态如图4所示。与原生林相类似,在月尺度上,有两个凋落峰,主峰出现在干热季(3—4月),次峰在雨季末期(10月)。但是,次生林的凋落峰相对于原生林而言不太明显。次生林的树高较原生林低被推测为导致该现象的原因。特别需要指出的是,2003—2004年度西双版纳勐仑地区经历了过去50年降雨最少的一年。由于2001和2002年为两个丰水年,土壤含水量最低值并不是出现在2003年,而是在2004年。水分亏缺导致了2004—2005年度旱季凋落物的迅速增加,凋落物的增加,促进了森林的呼吸异化作用,增加了碳排放,这个结果也表现在涡度碳通量的观测结果上[6,19-20]。同时,该结果也表明,造成西双版纳凋落物季节和年间动态上,干旱也起着非常重要的作用。
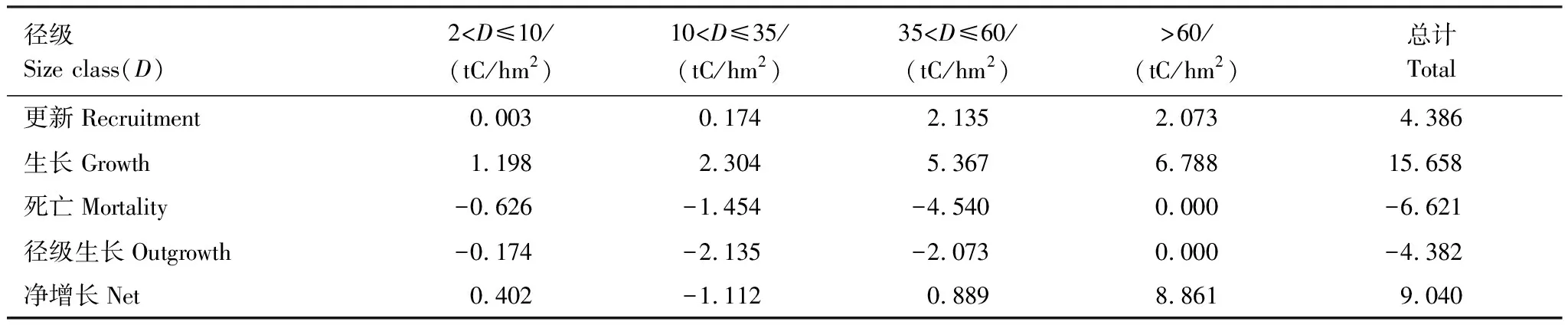
表3 样点3乔木生物量在2003—2007年之间的变化
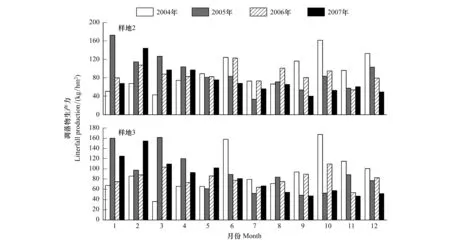
图4 西双版纳热带次生林的凋落物月动态
2.3 对热带森林碳蓄积的启示
虽然我们强调热带原始生林在固碳方面的作用,但是次生林在固碳方面的作用也不可小觑,特别是在快速生长期(图2)。从FLUXNET(碳通量观测网络)的数据来看,干扰是导致生态系统成为碳源的主要原因[21],未遭受明显干扰的森林基本上都表现出碳汇作用[22]。干扰可以导致原始林成为碳源[8],同时也使次生林的碳蓄积表现为负值(死亡的碳损失大于生物量增长的碳蓄积)(表2)。因此,不论是原生林还是次生林,加强森林的管理,竭力杜绝人为干扰,减弱自然干扰的影响,是促进森林碳蓄积的基础。
3 结论
(1)热带森林在演替过程中,森林的碳储量可以被划分为五个阶段:干扰发生(I);干扰后的迅速生长期(II);较慢的生长期(III);储量近稳定期(IV);下一次干扰发生(V)。在热带森林的演替过程中,存在一个迅速生长期,该生长时段森林的固碳速度达到最高。因此热带次生林在森林固碳方面也发挥着不可忽略的作用。
(2)不论原生林还是次生林,干扰是导致森林成为碳源的主要原因,因此加强森林的管理,竭力杜绝人为干扰,减弱自然干扰的影响,是促进森林碳蓄积的基础;推荐使用编制森林生物量动态表,以便详细获知造成森林碳蓄积或者损失的可能原因。
(3)热带次生林的凋落物动态与原生林的相类似,但是其季节趋势较弱。干旱事件是影响凋落物的季节和年间动态的原因,也是短时间尺度上影响碳平衡的一个重要因子。
