江苏近年育成粳稻新品种/系的稻瘟病抗性基因及穗颈瘟抗性分析
2020-09-17王小秋杜海波陈夕军李明友王嘉楠许志文冯志明陈宗祥左示敏
王小秋 杜海波 陈夕军 李明友 王嘉楠 许志文 冯志明, 陈宗祥,左示敏,,
(1 植物功能基因组学教育部重点实验室/江苏省作物基因组学和分子育种重点实验室/扬州大学 农学院,江苏 扬州 225009;2 扬州大学 江苏省粮食作物现代产业技术协同创新中心/江苏省作物遗传生理重点实验室,江苏 扬州 225009;*通信联系人,E-mail: smzuo@yzu.edu.cn)
稻瘟病是由稻瘟病菌(Magnaporthe oryzae)引起的真菌性病害,是全世界范围内影响水稻生产的重要病害之一,发病时会造成粮食损失,严重时甚至颗粒无收[1],被称作水稻的“癌症”[2]。根据危害时期和部位,稻瘟病可分为苗瘟、叶瘟、节瘟和穗颈瘟等,其中又以穗颈瘟对水稻生产的威胁最大[3]。穗颈瘟发生在水稻穗颈(穗轴与稻穗连接部位),一般在水稻抽穗破口前感染,在始穗后稻瘟菌侵入其中进行繁殖和危害,导致穗颈节坏死,阻断输送到穗的养分通道,形成瘪粒或白穗,严重影响水稻的产量,甚至会造成绝收[4]。目前对于稻瘟病的防控主要依赖化学药剂和选育抗病品种。相比于化学防治的诸多缺点[5],培育和推广抗病品种无疑是防治稻瘟病流行最经济安全和绿色高效的方法[6]。
定位克隆抗稻瘟病基因是开展抗病育种的重要前期工作。迄今已报道的稻瘟病质量抗性基因(resistance gene,R基因)有100 余个,数量性状基因(quantitative trait loci, QTL)500 余个,已克隆的R 基因有近27 个[7-8],多数分布在水稻第1、6和11 染色体上的几个座位[9]。通过相关基因的功能标记分析,发现Pita、Pb1、Pia、Pik、Pi-kh、Pish等基因在江苏粳稻品种中各有不同程度的分布,相反,Pi9/2座位中的不同广谱抗性基因在江苏粳稻品种中基本不存在[10-11]。李刚等[12]对江苏省554 份水稻种质资源进行抗稻瘟病基因分子检测,发现携带Pita、Pib和Pik基因的材料都达到了30%及以上,其中Pik所占比例达到了82.2%。朱勇良等[13]认为以Pita为代表的一些抗性基因对江苏粳稻的穗颈瘟抗性正在逐渐丧失,但王军等[14]对2007-2013 年江苏省审定的粳稻品种的穗颈瘟抗性研究表明,携带Pita、Pib和Pikm的材料穗颈瘟抗性总体较好。总体而言,在以上这些研究中大多缺少了对基因互作效应的分析。有研究表明,通过聚合多基因来改良品种抗病性时,需要选择合适的基因组合,从而使聚合的抗病基因间能有效互补甚至产生增效作用,达到增强抗性、拓宽抗谱的目的[15]。陈羽等[16]在扬稻6 号背景下,构建了Pigm、Pi40、Pi9、Pi2和Piz中的不同基因分别与Pi1、Pi33、Pi54间的两两聚合系,发现不同聚合系之间的抗性存在明显差异,表明不同抗病基因间的互作机制可能广泛存在,并最终影响聚合效果。相比同一背景下特定地研究某些基因之间的互作效应,在复杂遗传背景下评估分析不同基因间的聚合效应更符合育种实际,但目前还少有相关的研究报道。
水稻是江苏优势农作物,单产位于全国前列,总产占全国的10%左右。江苏水稻以粳稻为主,种植面积占总面积的85%以上,因此,研究分析江苏粳稻品种在已知稻瘟病抗性基因位点上的基因型信息并分析其对穗颈瘟的抗性贡献,具有十分重要的意义。利用14 个抗病基因的功能标记,本研究对江苏近年育成的195 份粳稻新品种/系在相关位点上的基因型信息进行分析;同时,通过6 个稻瘟菌小种混合接种,对其中的158 个品种以及17 份携带Pigm基因的回交株系进行穗颈瘟抗性鉴定;最后,通过逻辑回归,评估了对穗颈瘟抗性有显著贡献的基因和基因组合。该结果有利于指导江苏粳稻抗穗颈瘟育种实践,也可为同类研究提供借鉴和参考。
1 材料与方法
1.1 水稻材料
用于稻瘟病抗病基因分析的水稻品种/系共209份,包括江苏省2013-2019 年审定的粳稻品种66份,2013 年前审定但在2019 年江苏水稻生产上仍有一定推广面积的粳稻品种22 份,2019 年参加江苏省区域试验的粳稻新品种/系107 份,以及14 份携带已知稻瘟病基因抗、感病等位基因的阳、阴性对照品种。各材料具体见附表1 和表1,其中丽江新团黑谷(LTH)背景下的相关抗病基因单基因系由浙江省农业科学院王艳丽老师实验室惠赠。
用于穗颈瘟接种鉴定的材料有175 份,包括上述材料中的158 份和本实验室创建的携带抗稻瘟病基因Pigm的回交高世代株系17 份。回交高世代株系具体为粳稻品种淮稻119 背景下的BC6F3株系13份和武运粳32 背景下的BC4F3 株系4 份。
1.2 菌株培养及接种鉴定
穗颈瘟抗性鉴定菌株由江苏省农业科学院植物保护研究所刘永峰研究员实验室提供,为2018年江苏不同地区病样材料中分离到的6 个代表性菌株2019-16-3、2019-522、2019-578、2019-746、2019-863 和2019-942。将供试菌株移植到稻秆玉米粉培养基(玉米粉40 g、稻秆粉50 g、琼脂20 g)上,在28℃培养箱中黑暗培养7 d,之后用灭菌载玻片刮去培养基表面菌丝,在26℃黑光灯下照射72 h,待稻瘟病菌产生孢子后,用无菌水洗下,将各稻瘟病菌株孢子等比例混合,配成10×10 倍显微镜下每视野30~40 个孢子的悬浮液[17]。接种材料于2019 年5 月15 日播于扬州大学农学院试验田,于6 月14 日移栽,单本栽插,每区2 行,每行12株,常规水肥管理。
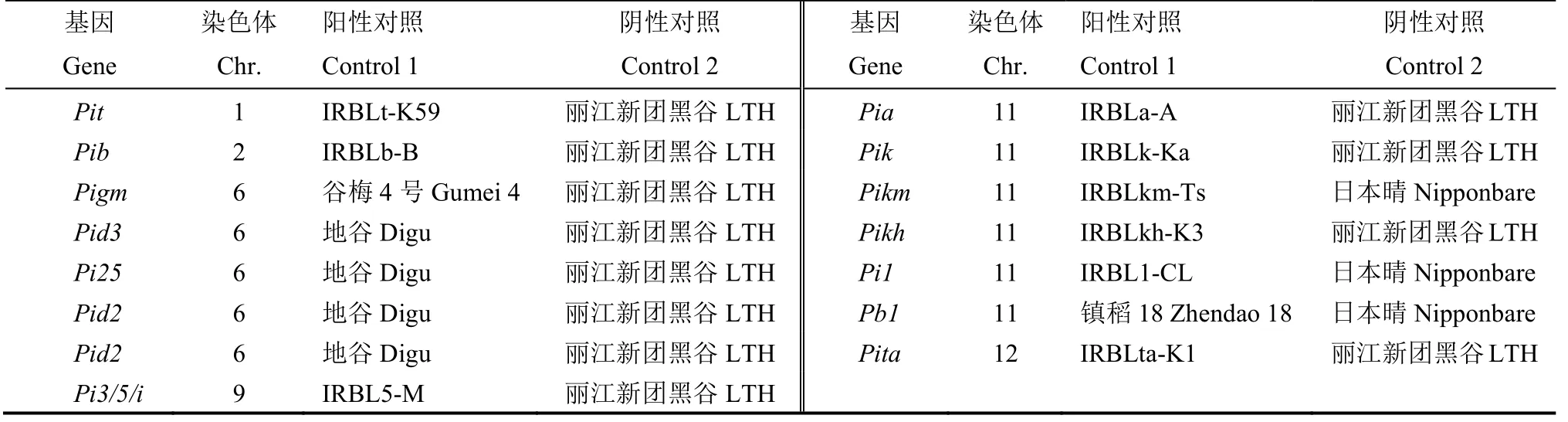
表1 本研究用于各抗病基因检测的阴阳性对照Table 1.Information of controls used in detection of resistance genes to blast disease in present study.
在水稻孕穗初期采用混合菌株进行接种,用注射器将接种液缓缓注入稻苞内,每小区接种5 株,每株接种1 个幼穗,每穗注射1 mL 孢子悬浮液。根据供试材料的生育期差异,分别于8 月14 日、20 日和27 日进行接种。接种后于水稻蜡熟期,按农业部发布的水稻品种试验稻瘟病抗性鉴定与评价技术规程中“水稻穗颈瘟单穗损失率0~9 级分级标准”进行病情调查[18]。在品种最终病级判定上,若5 个接种穗的病级级别间小于2 个分级差异时,则取最高病级;病级种类不小于3 个分级差异时,若高病级占多数,取最高病级;若低病级占多数,取最高病级的下一级为该品种的最终病级。
1.3 抗病或感病等位基因的功能标记
根据前人的研究报道,本研究对Pit、Pib、Pigm、Pid3、Pi25、Pid2、Pi3/5/i、Pia、Pik、Pikh、Pikm、Pi1、Pb1和Pita这14 个抗稻瘟病基因座位上的抗病和感病等位基因进行检测分析,采用的相关功能标记引物见表2。其中Pik功能标记为原菲律宾国际水稻研究所周波研究员团队惠赠。引物由北京擎科生物有限公司(南京)合成。
1.4 DNA 提取及PCR 检测
采用CTAB 法提取DNA。PCR 扩增反应总体积为20 μL,包括模板DNA 2 μL,上下游引物(10 μmol/L)各0.4 μL,2×Taq混合液(含染料)10 μL,灭菌双蒸馏水7.2 μL。PCR 程序如下:95℃下预变性5 min,94℃下变性30 s,55℃~60℃下(视不同引物而定)退火30 s,72℃下延伸30~72 s(视不同片段大小而定),共35 个循环;72℃下再延伸10 min。PCR 产物直接通过1%或4%(根据产物片段大小而定)琼脂糖凝胶(EB 染色)电泳分离,然后在紫外凝胶成像仪上观察并拍照或聚丙烯酰胺凝胶电泳成像。
1.5 统计分析
采用常用办公软件Excel 2016 对研究中获得的各类数据进行整理和作图。对品种携带的抗病基因数和穗颈瘟抗性间的相关分析,采用 SPSS 16.0.0.247 统计分析软件中的回归分析模型进行。对于中抗以上和中感以下两类品种间携带的抗病基因数比较,以及对携带不同基因或基因组合的品种间病级差异分析,均采用SPSS 16.0.0.247 统计分析软件中的常用方差分析(ANOVA)和t测验进行差异显著性检测。
采用SPSS 16.0.0.247 统计分析软件中的逻辑(logistic)回归模型[19],分析不同基因对穗颈瘟抗性的贡献。首先根据江苏水稻新品种审定过程中对稻瘟病抗性级别要求不低于5 级的标准将供试品种表型重新划分为两个等级,即病级大于5 级的定义为1,小于等于5 级的定义为2 级;其次,分别采用单基因逐一逻辑回归和多基因逻辑回归,获得各基因与抗性间的回归系数(B)、显著性P值、95%CI(confidence interval)值、优势比(odds ratio,OR,即Exp B);最后,根据P值确定基因的效应是否达到统计显著水平,进而根据OR 值判断基因的效应,相关结果采用森林图进行展示。
2 结果与分析
2.1 抗病基因功能标记的扩增验证
为了验证各抗稻瘟病基因功能标记的PCR 扩增结果是否与预期一致,利用14 个稻瘟病抗性基因的功能标记,分别对已知或已报道的携带相应抗病基因的供体(含抗病单基因系)和未携带相应抗病基因的阴性对照进行PCR 检测,同时随机检测10 个江苏粳稻品种。结果显示(图1),采用一对引物或单标记即可区分抗、感等位基因的9 个抗病基因,在具有相应抗病基因阳性对照中均能扩增出与预期大小一致的片段,而在阴性对照中均未扩增出或扩增出与携带感病等位基因预期大小的片段;对于Pit基因,采用3 条引物(tRn1、tRn2 和tN11)同时进行PCR 扩增时,在阳性对照中分别出现733 bp 和530 bp 两个片段,而在阴性对照中没有扩增条带,与理论预期一致;对于Pigm基因,在阳性对照中扩增出两条带,一条为内参标记Actin1-1的扩增片段,另一条为Pigm基因特异标记GMR-3 的扩增片段,而在阴性对照上只有Actin1-1标记的扩增片段,与预期一致;对于Pib和Pita,在抗病对照中均只有采用抗病基因功能标记才能扩增出预期片段,而在阴性对照中也只能采用感病基因特异标记才能扩增出预期片段;对于Pikm基因,因其座位上存在两个相互作用才能发挥抗病功能的基因Pikm1-TS和Pikm2-TS,因此需要两对标记进行检测,只有同时检测到两个基因预期片段大小的材料才能被认为携有Pikm,结果显示两个标记在阳性和阴性对照上的扩增结果也与预期结果一致。随机选择的10 个粳稻品种用不同基因标记扩增呈现不同结果,表明品种间携带的抗、感等位基因的不同。以上结果说明,本研究采用的各抗病基因功能标记可以用于评价后续江苏粳稻品种中携带的相关抗病基因信息。
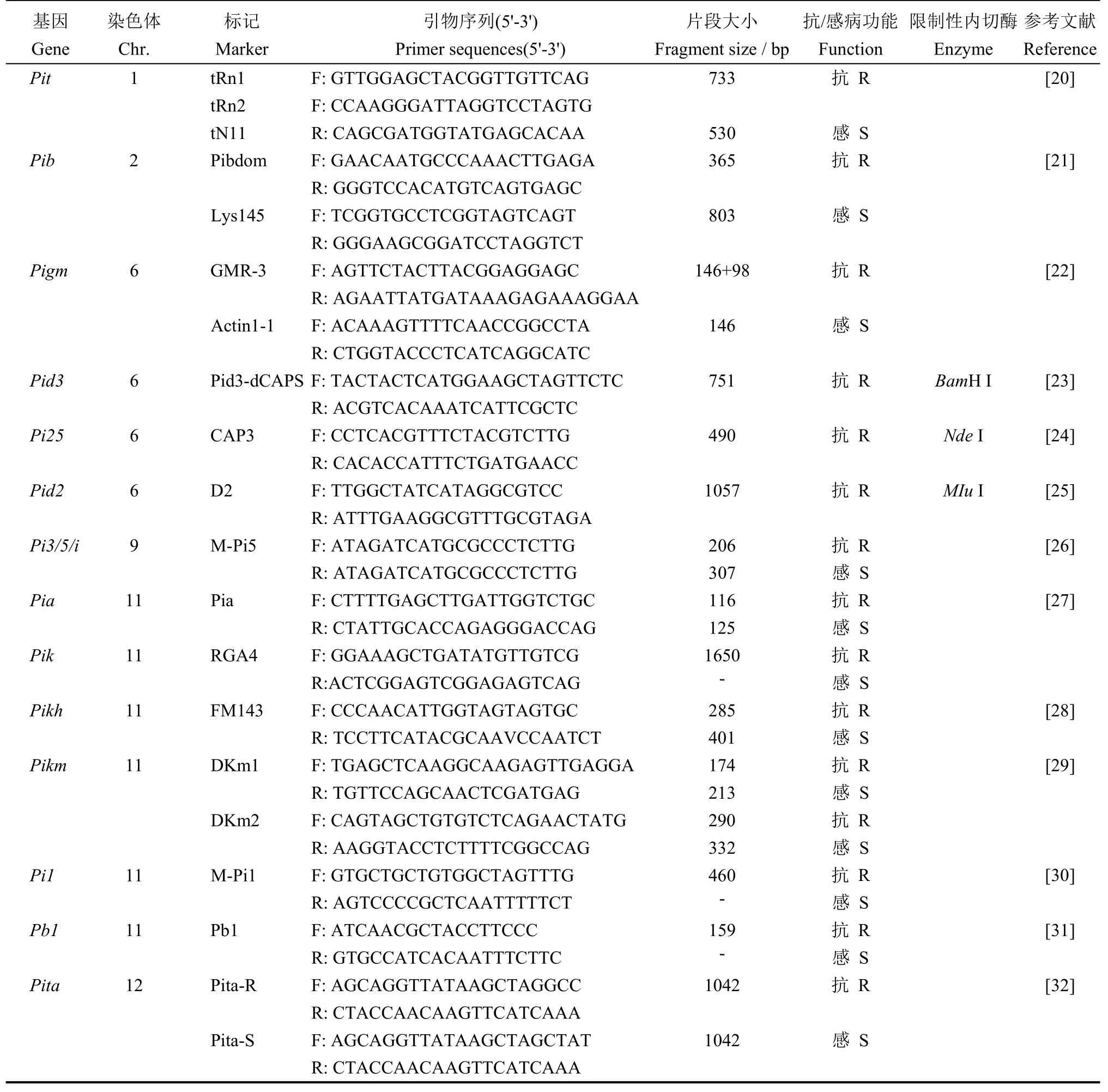
表2 本研究用于各抗病基因检测的功能标记引物信息Table 2.Information of molecular markers used in detection of resistant genes to blast disease in present study.
2.2 各抗病基因在江苏粳稻品种/系中的应用情况
利用各功能标记对供试的江苏粳稻品种/系进行检测,结果显示(图2-A),所有材料均不带有抗病基因Pigm,其余13 个抗病基因则有不同程度的分布;Pi25与Pit出现频率较低,分别为3%和6%;大部分的基因出现频率为10%~30%,在50%左右的有2 个,为Pikh和Pita;Pib基因出现频率最高,达到72%,即有141 份材料检测到含有Pib。就品种而言,多数品种携带的基因数为2~5 个,有20 个品种各只携带1 个基因,有6 个品种携带7 个以上基因且其中1 个品种W046 携带抗病基因数达到9 个,为供试品种中携带抗稻瘟病基因数目最多的品种(图2-B)。以上结果表明,江苏粳稻品种在携带的已知抗稻瘟病基因上存在明显差异,Pikh、Pita和Pib总体应用较广,且又以Pib的应用最广。
检测的供试粳稻品种包括中熟晚粳、早熟晚粳、迟熟中粳和中熟中粳四种生态类型。为了分析不同生态类型品种间携带的抗病基因是否存在差异,对各基因在供试材料中的分布频率进行比较。由于中熟晚粳稻生态区较小且供试的品种数只有5 份,因此将其与早熟晚粳合并分析。结果显示(图3),大多数基因在三种生态类型间的出现频率相近,少数基因存在明显差异,如Pit和Pi3/5/i基因。Pit基因的检出频率总体较低,但更多分布在中熟中粳中,且在迟熟中粳中未检测到该基因。Pi3/5/i基因在中熟中粳中的出现频率是其在另两类品种中的近4 倍。此外,有少数基因在不同品种类型间存在较小的分布差异,如Pid2、Pik较多存在于早/中熟晚粳中,Pid3较多存在于迟熟中粳中,Pia则较多在早/中熟晚粳和中熟中粳中。结果表明,多数抗病基因在不同生态型品种育种中得到了较为相近的应用,仅有少数基因在不同生态类型品种间存在分布差异。
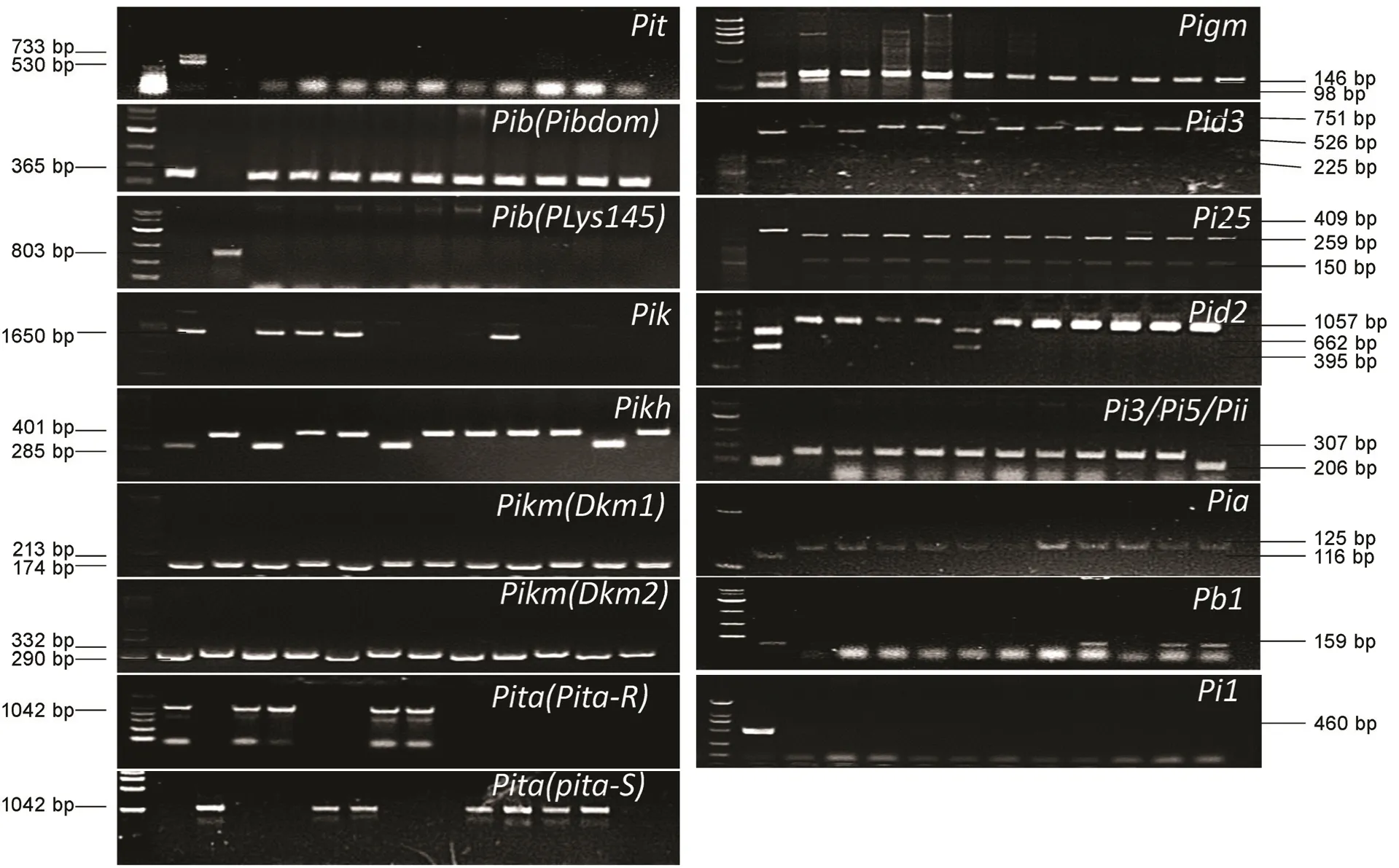
图1 各抗病基因功能标记在阳性、阴性等品种/系上的扩增验证Fig.1.Validation of the molecular markers in determining the corresponding resistance gene using known positive and negative varieties or lines as well as 10japonica varieties randomly selected from Jiangsu Province.
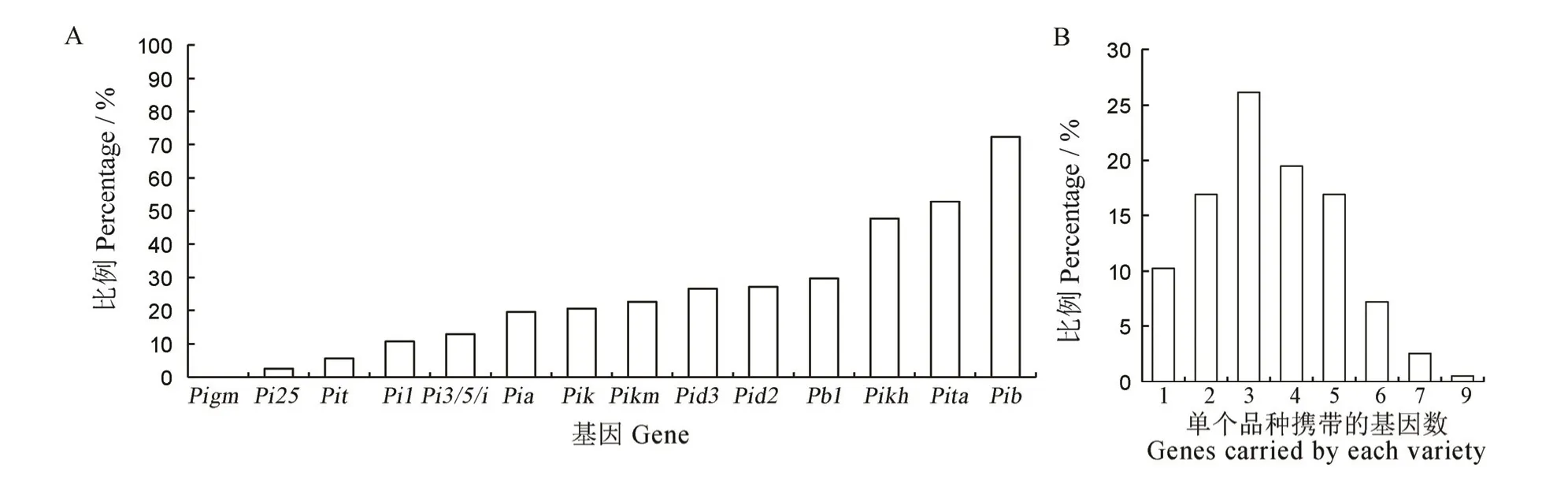
图2 各抗病基因在江苏粳稻品种中的分布频率(A)及品种携带不同抗病基因数分布(B)Fig.2.Distribution frequency of resistance genes injaponica rice from Jiangsu Province(A) and the distribution of gene number carried by varieties(B).
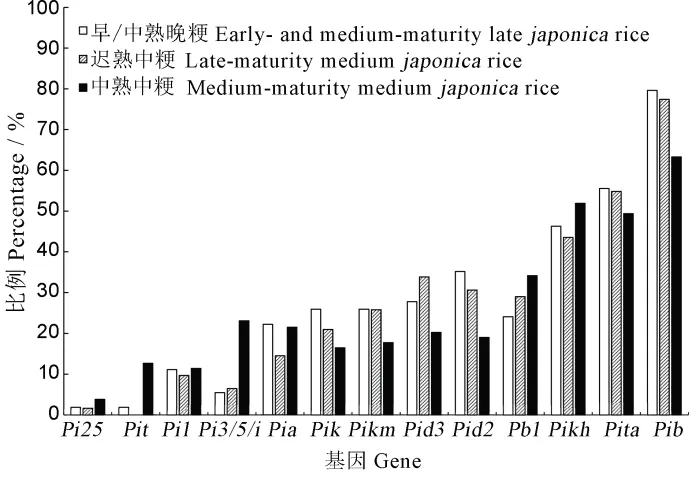
图3 抗病基因在江苏不同生态型粳稻品种中的分布频率Fig.3.Distribution of resistance genes in different types ofjaponica rice.
由于供试品种/系涉及已经审定(以下简称审定)和正在参加江苏省新品种区域试验(以下简称区试)的两大类型材料,因此,进一步比较各基因在审定和区试品种间出现的频率差异。结果显示(图4),Pid3、Pid2、Pia、Pb1这4 个基因在区试品种/系中的占比明显高于其在审定品种中的占比,频率差异在14%至23%之间;Pik、Pikh、Pi1和Pita在区试中的频率同样高于审定品种中,但差值均不足12%;其他基因的分布频率在两类品种间基本接近。总体而言,这些抗性基因在区试品种中的应用频率要高于审定品种。
2.3 供试品种穗颈瘟抗性分析
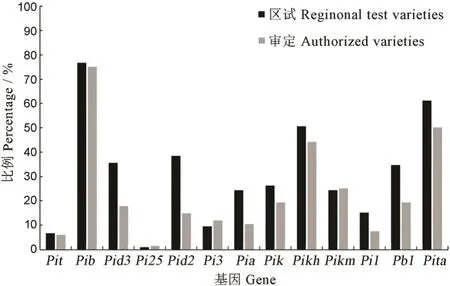
图4 各抗病基因在2019 年前审定及2019 年区试品种/系间的频率分布比较Fig.4.Comparison of resistance gene distribution frequency between varieties authorized before 2019 and regional test varieties in 2019.
为了尽可能排除生育期差异较大影响人工接种鉴定结果,本研究选择了接种时生育进程尽可能一致的分蘖穗进行接种,最终进行穗颈瘟人工接种鉴定的材料有158 份,其中早熟晚粳47 份、中熟晚粳5 份、中熟中粳47 份、迟熟中粳59 份。结果显示(图5-A),158 份供试品种中,病级在7 级或以上的品种达到了72%,达到中抗(病级为3 级)、抗病(病级为1 级)及高抗(病级为0)的材料也只有35 份。各种生态类型中的抗感品种分布总体与总供试品种中的抗病分布趋势相同,但中熟中粳类型中达到中抗以上品种/系的比例明显高于另两种生态类型,中熟中粳与早/中熟晚粳类型品种中的抗和高抗病品种比例相当。进一步比较了2019 年前审定品种和2019 年区试品种中病级小于等于5级的品种比例(图5-B),结果显示,区试品种中小于等于5 级的品种比例(42.9%)明显高于审定品种中的比例(14.8%)。以上结果表明江苏粳稻品种的穗颈瘟抗性总体较弱,区试中的新品种抗性水平总体好于已审定的推广品种。
2.4 品种携带的抗病基因数与穗颈瘟抗性相关分析
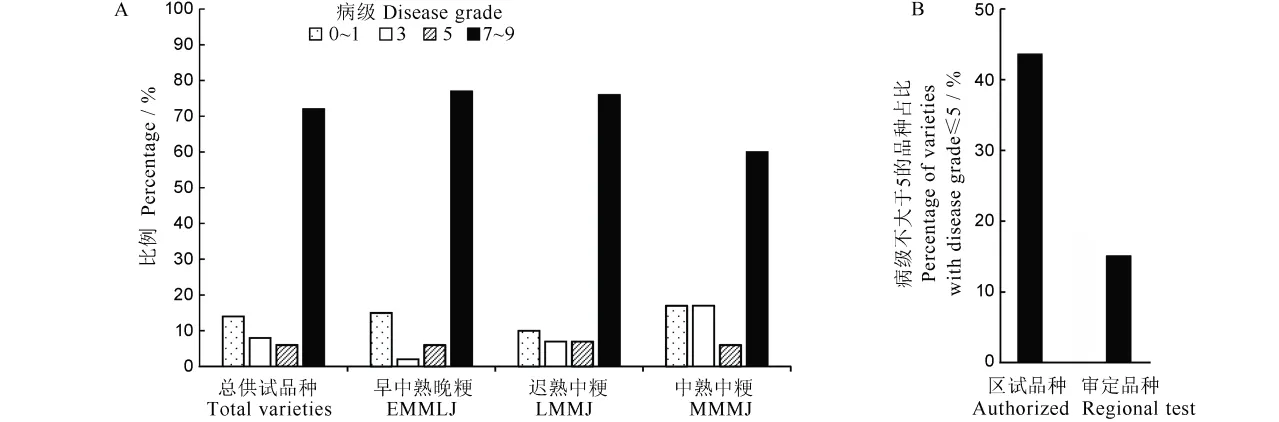
图5 供试品种稻瘟病病级分布Fig.5.Distribution of disease grade in varieties tested.
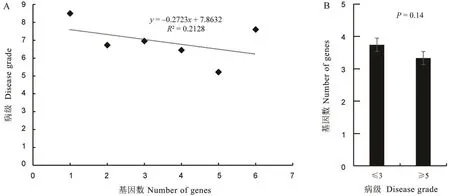
图6 品种携带抗病基因数与病级间的关系分析Fig.6.Analysis of the resistance gene number carried by each variety and disease grade.
对供试品种分别携带的抗病基因数与穗颈瘟抗性病级进行回归分析,发现携带的抗病基因数多少与穗颈瘟抗性强弱间的决定系数R2为0.2128,未达统计显著水平(P值为0.357),表明两者相关不显著(图6-A)。进一步统计比较病级≤3 的中抗以上品种和病级≥5 的中感以上品种间携带的抗性基因数差异(图6-B),发现两类品种携带的平均抗性基因数分别为3.7 和3.3,差异不显著。在供试品种中,发现有品种仅携带少数几个基因但却表现较强的穗颈瘟抗性,如宁5916,只携带Pia和Pita两个基因,接种鉴定为高抗;同时也发现携带抗性基因数较多却表现出高感的品种。以上结果表明,在抗稻瘟病育种中,不能简单地通过增加基因数来实现品种的抗性改良。
2.5 优异抗病基因和基因组合分析
为分析哪个/些基因对本次混合小种有较好的抗性,首先采用单基因逻辑回归方法评估了各基因对抗性的贡献,结果显示(图7-A),只有Pia和Pi3/5/i两个基因与抗性间的回归系数分别达到极显著相关,回归系数分别为2.20(P=0.000)和1.19(P=0.017),说明在单基因水平上其他基因对抗性均无显著贡献;进一步计算得出Pia的Odds ratio(OR)值最高,为9.01,表明携带该基因对抗性的贡献是不带该基因的9 倍;其次为Pi3/5/i,其OR值为3.30。考虑到基因之间可能广泛存在互作效应,进一步采用多基因逻辑回归方法评价了各基因对抗性的贡献,结果显示(图7-B),除了Pia和Pi3/5/i外,Pita基因对抗性的贡献同样达到了极显著水平,而其他10 个基因均未达到显著相关,表明他们存在与否对品种的抗性无影响;结合OR 值,可知三个显著相关的基因贡献率从高至低依次为Pia(OR=28.42)、Pi3/Pi5/Pii(OR=16.02)和Pita(OR=7.49)。
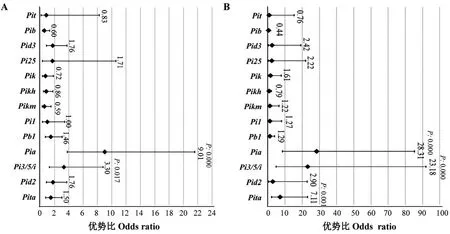
图7 各抗病基因对穗颈瘟抗性的贡献逻辑回归统计参数Fig.7.Logistic regression statistical analysis about the contribution of each resistance genes to neck blast resistance.
相比单基因逻辑回归结果,多基因分析中增加了一个对抗性显著贡献的基因Pita,暗示Pita基因可能通过与另两个基因间的互作提供抗性。为此,进一步统计比较Pia、Pi3/5/i和Pita这3 个基因不同组合方式间的品种病级差异。由于供试品种中未发现3 基因聚合的品种,同时Pia+Pi3/5/i双基因聚合品种的样本数也只有2 个,因此,这两种组合未进入比较分析。结果显示(图8),与3 个基因均不带的对照品种平均病级(7.77)相比,携带Pi3/5/i(5.46)和Pia(5.94)中任一个基因的品种平均病级均得到了显著降低,但两者之间差异不显著,而仅携带Pita单基因的品种病级与对照差异不显著;就基因组合而言,发现携带Pi3/5/i+Pita双基因的品种病级(3.00)显著低于Pi3/5/i和Pia单基因品种病级,但又显著高于携带Pia+Pita双基因的品种(0.38),说明尽管Pita基因单独存在时的抗性效应不明显,但在与另两个基因聚合时,则可显著增强另两个基因的抗病效应,且Pia与Pita间的聚合效应显著高于Pi3/5/i+Pita。随后,发现在供试的158 份品种中,所有携带Pia+Pita双基因的13 个品种其病级均为0 级或1 级。
以上结果表明,在单基因状态下Pi3/5/i和Pia可显著提升供试品种的穗颈瘟抗性,但抗性效应不如其分别与Pita聚合的效应,聚合状态下又以Pia+Pita间的聚合效果最好,利用该基因组合有望显著提升江苏粳稻品种的穗颈瘟抗性。
2.6 携带Pigm 基因回交株系的穗颈瘟抗性分析
由于在供试材料中未检测到携带Pigm基因的品种,因此本研究分别利用 HD119 和武运粳32(WYJ32)背景中导入Pigm的回交株系为材料,对Pigm基因的抗病效果进行了分析。结果显示(图9-A、B):对照HD119 的抗性级别为7 级,为感病水平,而其背景下的13 个携带Pigm的高世代回交株系的抗性水平均达到中抗及以上,其中10 份病级均为0,达到了高抗水平;对照WYJ32 表现为高感穗颈瘟,在其背景下导入Pigm的4 份高世代回交株系中,有2 份分别达到抗和高抗水平。这表明利用Pigm基因可显著提高江苏粳稻品种的穗颈瘟抗性。
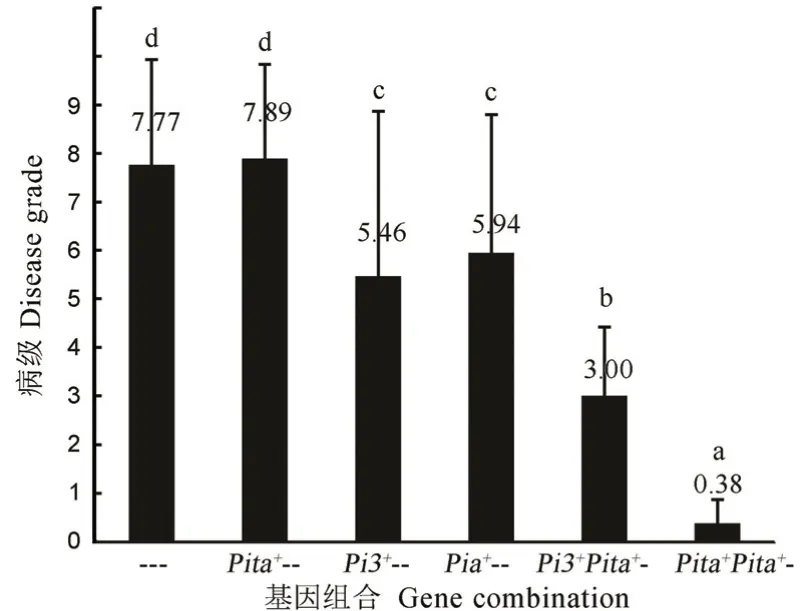
图8 三个对穗颈瘟抗性有显著贡献的基因及组合间的品种病级差异比较Fig.8.Contribute significantly of three genes to the resistance of neck blast resistance and the comparison of disease grade differences among them and their combinations.

图9 携带Pigm 高世代回交株系的穗颈瘟抗性表现Fig.9.Resistance performance of advanced backcrossing lines withPigm and the corresponding recurrent parents.
3 讨论
3.1 江苏近年育成的粳稻新品种/系对穗颈瘟的抗性总体较弱
由于每年流行的稻瘟病菌优势生理小种可能不同,并直接影响品种的抗性表型[33]。因此,就生产实践而言,定期采集新近流行的小种对推广或审定品种的稻瘟病抗性尤其是穗颈瘟抗性进行鉴定和评价,具有重要的现实意义。本研究利用江苏省植物保护研究所刘永峰团队2018 年采集分离的6个代表性小种,对江苏近年育成的粳稻新品种/系的进行穗颈瘟接种鉴定。发现2019 年前审定的品种中病级小于等于5 级的品种比例(14.8%)明显低于2019 年区试新品种(42.9%)(图5-B),这暗示当年审定时抗性过关的多数品种可能不适应新流行的优势小种了。另外,在供试的158 份粳稻品种中,病级小于等于5 级的品种占比不到30%,大多数品种表现感和高感(图5-A)。供试品种总体可分为4 种生态型,覆盖江苏从南至北的所有粳稻生态区,比较显示,尽管中熟中粳中抗性好的品种比例略高于另两种生态型,但总体差异不太明显(图5-A)。这暗示江苏水稻生产上的稻瘟病暴发流行风险巨大。
3.2 江苏粳稻品种的穗颈瘟抗性改良方法
通过分子标记辅助目标基因选择是开展抗稻瘟病育种的最高效方法,但是需要了解目标基因的分布状态和潜在的育种价值。通过对14 个抗稻瘟病基因的检测分析(图2),发现除Pigm外其余的13 个抗病基因在供试材料中都有分布,其中Pib分布频率最高,达到了72%,这与陈涛等[33]、张善磊等[34]和李刚等[12]研究结果一致,表明Pib在江苏粳稻育种中得到了广泛应用。除此之外,Pita和Pikh在江苏粳稻品种中也有相对较多的分布,其余基因的出现频率均在30%以内,而Pi25、Pit、Pi1和Pi3/5/i甚至不足15%。有意思的是,相对于已审定品种,Pid3、Pid2、Pia、Pb1等4 个基因在区试品种/系中的出现频率明显高于已审定品种(图4),暗示这些基因在江苏近年粳稻育种中的应用频率得到了明显提高。就品种而言,发现绝大多数品种携带2~5 个基因,也有个别品种/系携带了7 个和9个基因(图2-B),但我们发现品种携带的基因数与抗性没有显著相关性(图6),这暗示在以基因对基因抗性为主的抗稻瘟病育种中,需要考虑的是关键基因或组合的应用,而不是简单的进行抗病基因累加。
通过逻辑回归分析(图7),发现除了Pigm基因以外,其余13 个基因中只有Pia、Pi3/5/i和Pita对穗颈瘟抗性有显著贡献。通过病级数据差异比较,发现Pita单独存在时的抗性效应不显著,而Pia和Pi3/5/i单独存在时均可显著提高抗性,且两者间的抗性效应相当(图8)。这一结果与朱勇良等[13]认为以Pita为代表的一些抗性基因对江苏粳稻的穗颈瘟抗性正在逐渐丧失的结论相近,但与王军等[14]对2007-2013 年江苏省审定的粳稻品种的研究结论不尽一致,其认为携带Pita、Pib和Pikm的材料穗颈瘟抗性总体较好。此外,曾晓珊等[35]利用江苏的72 个菌株对Pia基因的供体爱知旭进行了抗谱分析,发现抗性频率达到了83.3%,这与本研究的结果一致。但是,也有研究发现Pia的抗性较弱,如Abdullah 等[36]通过离体接种的方法,评价了江苏67个水稻品种对已经明确抗瘟基因型反映的22 个稻瘟病菌株的抗性水平,发现携带有Pia基因的品种抗性并不理想。总体而言,产生以上不尽一致的结果可能与彼此选用的材料、接种方法和接种菌株不同有关。
在针对上述3个基因的抗性效应评价中(图8),尽管Pita单独存在时没有抗性效应,但当其与Pia或Pi3/5/i间聚合时,可显著增强后者的抗病效应,表明Pita与这两个基因间均存在显著的正向互作效应,且Pita+Pia的互作效应显著强于Pita+Pi3/5/i,携带Pia+Pita双基因聚合的品种均达到了抗病和高抗水平。有关稻瘟病基因间可能存在的不同互作关系,前人已有不少研究[15,29,37]。这表明在育种实践中,选择合适的基因组合可起到事半功倍的效果,否则将适得其反甚至徒劳无功。本研究显示,在当前和今后的江苏粳稻抗稻瘟病育种中,应该重视Pia与Pi3/5/i或Pita间的聚合育种,尤其是与Pita间的聚合育种。可喜的是,2019 年区试新品种中Pia的检出频率明显高于已审定品种,加上有近一半的江苏粳稻品种携带了Pita,因此,未来应该可以快速实现这两个基因在江苏粳稻品种中的聚合育种,提升江苏粳稻品种的穗颈瘟抗性水平。
除了通过基因聚合实现品种广谱性和持久性抗性外,挖掘和利用广谱持久性抗性单基因无疑具有更重要的价值。从谷梅4 号中克隆到的Pigm被认为具有广谱持久的稻瘟病抗性[7,38]。目前Pigm已被广泛用于国内多个水稻生态区的稻瘟病抗病育种[39-40]。曾生元等[41]通过标记辅助选择,将Pigm导入江苏三个不同生态型粳稻品种中,发现导入系对南方毒性小种均表现出较强的穗颈瘟抗性。李育红等[42]将Pigm基因导入到生产上广泛使用的光温敏感型核不育系广占63-2S 中,育成了苗瘟和穗瘟抗性显著提升的新光温敏感型核不育系扬籼3S。本研究发现,江苏近年育成新品种/系中均不含有Pigm基因,通过标记辅助选择,分别在感病品种HD119和WYJ32 背景中构建了14 份回交高世代株系,接种鉴定结果显示,所有携带Pigm的株系均表现为抗病或高抗(图9-A, B)。这表明,Pigm在江苏粳稻穗颈瘟抗性改良中具有较高的育种利用价值。
以上研究表明,抗病基因组合“Pia+Pita”及抗病基因Pigm在江苏粳稻抗穗颈瘟育种中具重要应用价值,当然今后仍然需要不断增加接种小种的数量,以评估其抗性广谱性和持久性。另外,这三个基因位于不同的染色体位置,下一步有必要详细评估三个基因间的不同组合效应,包括三者间的聚合是否有利于进一步增强江苏粳稻品种对穗颈瘟的广谱性和持久性抗性。
