利用CRISPR/Cas9 技术高效创制长粒香型水稻
2020-09-17徐善斌郑洪亮刘利锋卜庆云李秀峰邹德堂
徐善斌 郑洪亮 刘利锋 卜庆云 李秀峰, 邹德堂,
(1 东北农业大学 寒地粮食作物种质创新与生理生态教育部重点实验室, 哈尔滨 150030;2 中国科学院东北地理与农业生态研究所, 哈尔滨 150081;3 黑龙江省种业技术服务中心, 哈尔滨 150008;*通信联系人, E-mail: lixiufeng@iga.ac.cn; 13603609603@163.com)
水稻作为全球最重要的粮食作物之一,全世界半数以上的人口均食用稻米,水稻是保障中国粮食安全与实现农业可持续发展的主粮作物[1]。近年来,人们对高产优质稻米的需求量越来越大,特别是长粒香型稻米,如稻花香的价格是一般非香型大米的2~4 倍,长粒香米更易受到农民和消费者们的青睐。传统的水稻品种改良过程中,主要通过杂交、回交的方式实现优良基因的聚合,但传统育种具有育种周期长、效率低等缺点,而基因编辑技术可以为水稻品种的快速改良带来新的契机。
目前,CRISPR/Cas9 技术作为新一代的基因编辑技术,在作物遗传育种研究中应用较广泛[2-7],该技术可在水稻基因组水平上进行基因定向编辑改造[8-11]。水稻中已经鉴定出许多与籽粒大小相关的数量性状位点(QTL),如负调控因子GW2[12]、GS3[13]、GS9[14]、qW5/GW5[15]和TGW6[16],正调控因 子GL2[17]、GL3.1[18]、GS5[19]、GL7[20]、GLW7/OsSPL13[21]和GW8/OsSPL16[22]。Li 等[23]利用该技术编辑水稻中的GS3基因,T2代的粒长和粒重增加,产量得到提高。Zhao 等[14]使用CRISPR/Cas9 技术,在GS9第1 外显子处插入1 bp,造成移码突变,破坏GS9正常表达,使粒长增加。
香味是水稻重要的品质性状之一,由第8 染色体上的Badh2基因控制。Badh2编码一个由503 个氨基酸组成的蛋白产物[24]。Chen 等[25]研究发现,在香稻品种中,由于Badh2基因发生突变,使得香味的主要成分2-乙酰-1-吡咯啉(2-AP)含量增加,稻米产生香味。Shan 等[26]通过TALEN 技术,对Badh2进行基因组编辑,稻米的2-AP 含量从无增加至0.35~0.75 mg/kg,获得具有香味的稻米。孙慧宇等[27]利用CRISPR/Cas9 技术对Badh2进行编辑,使寒地粳稻东农425 香味获得了改良,但东农425适应积温区较小。因此,我们拟利用CRISPR/Cas9技术对黑龙江省第二、三积温带品种龙粳11 进行香味改良,从而扩大香稻的种植面积。龙粳11 属早熟品种,其抗稻瘟病较强,抗倒伏,前、后生育时期耐寒性强。同时,龙粳11 的稻米品质优良,各项指标均达到国家优质米标准。
本研究利用CRISPR/Cas9 技术,以龙粳11 为研究材料,对GS3、GS9和Badh2基因同时进行敲除,能够高效地将圆粒水稻变成长粒香型水稻,可为加快水稻的种质资源改良提供一定的理论指导。
1 材料与方法
1.1 试验材料
本研究以粳稻龙粳11 为试验材料。在黑龙江哈尔滨(北纬45°)和海南(北纬19°),均以25 cm×25 cm 的株行距种植,自然长日照条件下生长。
试验所用 pYL-U6a-gRNA 和 pYLCRISPR/Cas9Pubi-H 双元载体[28]由华南农业大学刘耀光院士惠赠。
1.2 gRNA 靶点接头设计及载体构建
通过NCBI(https://www.ncbi.nlm.nih.gov/)获得GS3(Os03g0407400)、GS9(Os09g0448500)、Badh2(Os08g0424500)的基因序列,利用CRISPRGE 在线网站(http://skl.scau.edu.cn/)[29],分别在GS3第1 外显子、GS9第1 外显子和Badh2第2 外显子处设计1 对靶点接头引物,B1-F/R、B2-F/R 及B3-F/R(表1)。利用软件CRISPR Primer Designer设计GS3、GS9和Badh2基因靶序列,比对水稻序列,检测设计靶点是否特异,排除潜在脱靶序列。
参照Ma 等[30]的实验方法进行载体构建,将连接好的载体通过热激法转入大肠杆菌Match1-T1中,菌落PCR 检测后,选取阳性菌落摇菌后提取质粒。利用检测引物NOSF/M13F-47(表1)对载体质粒进行PCR 扩增,对产物进行测序,将检测无误的质粒通过热激法转入EHA105 农杆菌感受态细胞中。

表1 本研究中所用的引物Table 1.Primers used in this research.
1.3 T0 代阳性植株的获得
通过农杆菌介导法,将CRISPR/Cas9 表达载体转入龙粳11 的愈伤组织中,经过潮霉素抗性筛选愈伤组织,分化成T0代植株。在T0代植株成熟期时,采用CTAB 的方法提取叶片的全基因组DNA,通过引物Hyg-F/R 进行检测,PCR 产物大小为289 bp 的植株为阳性转基因植株,筛选出阳性苗。设计引物Seq1-F/R、Seq2-F/R 和Seq3-F/R,分别扩增GS3、GS9和Badh2基因靶点及其附近的序列,使用兼并序列解码(DSDecode)法[31]分析测序结果。
1.4 T1 代无T-DNA 元件的纯合突变植株的筛选
T1代植株成熟时期,提取叶片基因组DNA,利用引物Hyg-F/R(表1)筛选出无T-DNA 元件插入的植株。再利用测序引物Seq1-F/R、Seq2-F/R 和Seq3-F/R,对无T-DNA 元件插入的植株进行PCR扩增,测序结果通过DSDecode 方法,筛选出无T-DNA 元件的三基因纯合突变的植株,将其自交繁殖至T2代。
1.5 T2 代植株农艺性状考查
在大田环境下种植龙粳11野生型和T2代植株,成熟时期分别测量粒长、粒宽、结实率、单株产量及千粒重,并采用气相色谱-质谱联用技术,分析水稻籽粒中的2-乙酰-1-吡咯啉(2-AP)含量。利用SPSS 18.0 软件对农艺性状数据进行t检验。
2 结果与分析
2.1 水稻GS3、GS9 和Badh2 敲除靶点设计
GS3基因位于第3 染色体,有5 个外显子,编码232 个氨基酸。GS9基因位于第9 染色体,有4个外显子,编码345 个氨基酸。Badh2基因位于第8 染色体上,有15 个外显子,编码503 个氨基酸。GS3、GS9和Badh2基因CDS 序列均与日本晴相同,本研究分别在GS3第1 外显子、GS9第1 外显子、Badh2第2 外显子处设计1 对gRNA 靶点接引物,分别对GS3、GS9、Badh2基因进行编辑(图1)。
2.2 GS3、GS9 和Badh2 基因表达载体的构建
根据“金门”克隆法将带有3 个靶点的gRNA 表达盒连接到pYLCRISPR/Cas9Pubi-H 载体骨架上,构建好的载体即为 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 载体(图2)。
2.3 T0 代转基因阳性苗检测及突变情况
16 株T0代再生苗中有8 株为阳性苗,阳性率为50%(图3)。利用测序引物Seq1-F/R、Seq2-F/R和Seq3-F/R 扩增8 株阳性植株的靶点区域序列,并进行PCR 扩增,测序比对后,8 株阳性植株中有4株发生了突变,突变率为50%。

图1 GS3、GS9 和Badh2 基因结构和靶点位置Fig.1.Gene structure and target site ofGS3,GS9 and Badh2.

图2 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 载体Fig.2.pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA vector.
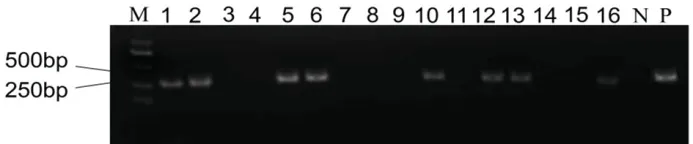
图3 T0 代植株转基因检测Fig.3.Transgenic detection of T0 generation plants.
2.4 T1代无T-DNA 元件三基因纯合突变植株的筛选
将三基因突变植株自交繁殖获得T1代植株,于成熟期提取叶片基因组DNA,利用引物Hyg-F/R筛选出无T-DNA 元件插入的植株。再利用引物Seq1-F/R、Seq2-F/R 和Seq3-F/R 进行扩增,通过DSDecode 方法分析测序结果,筛选出 2 株无T-DNA 元件的三基因纯合突变植株(55-2、58-2),分析GS3、GS9、Badh2基因突变的具体情况(图4)。
2.5 T2 代植株农艺性状分析及香味鉴定
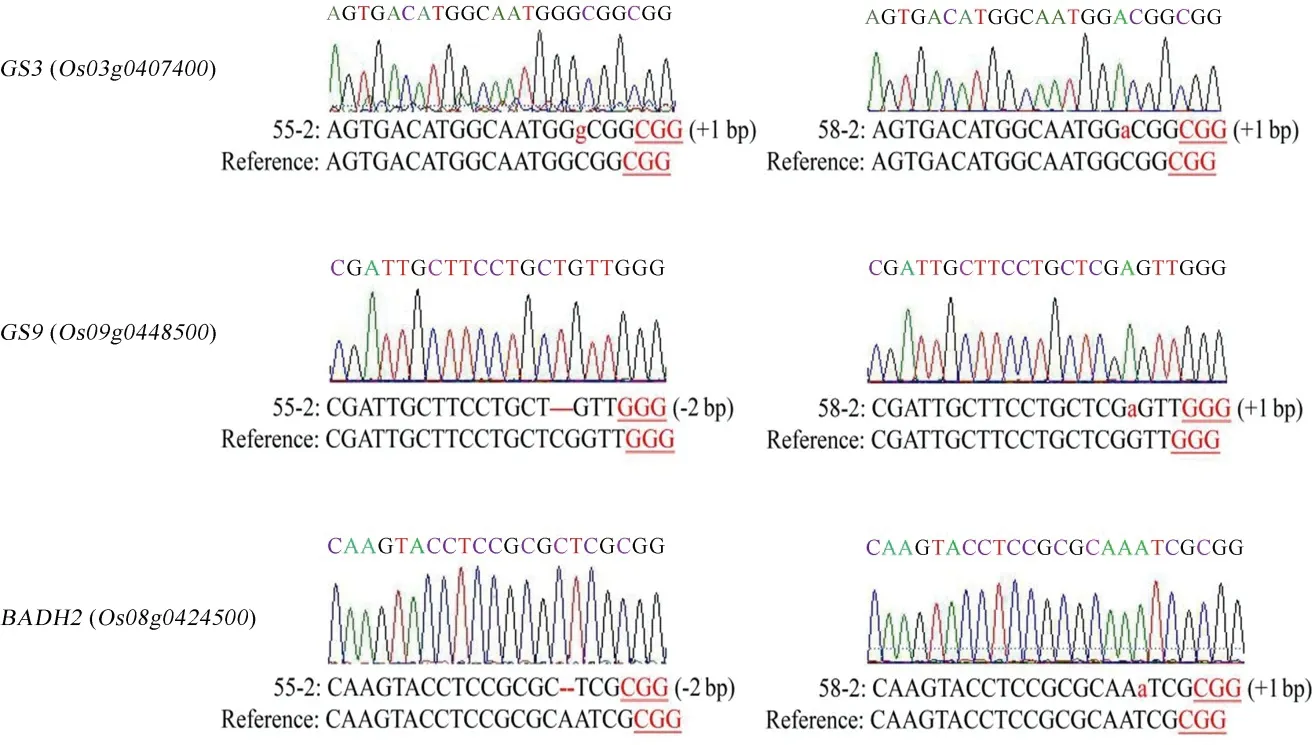
图4 三基因纯合突变植株测序结果Fig.4.Sequencing result of homozygous plants with triple gene mutation.
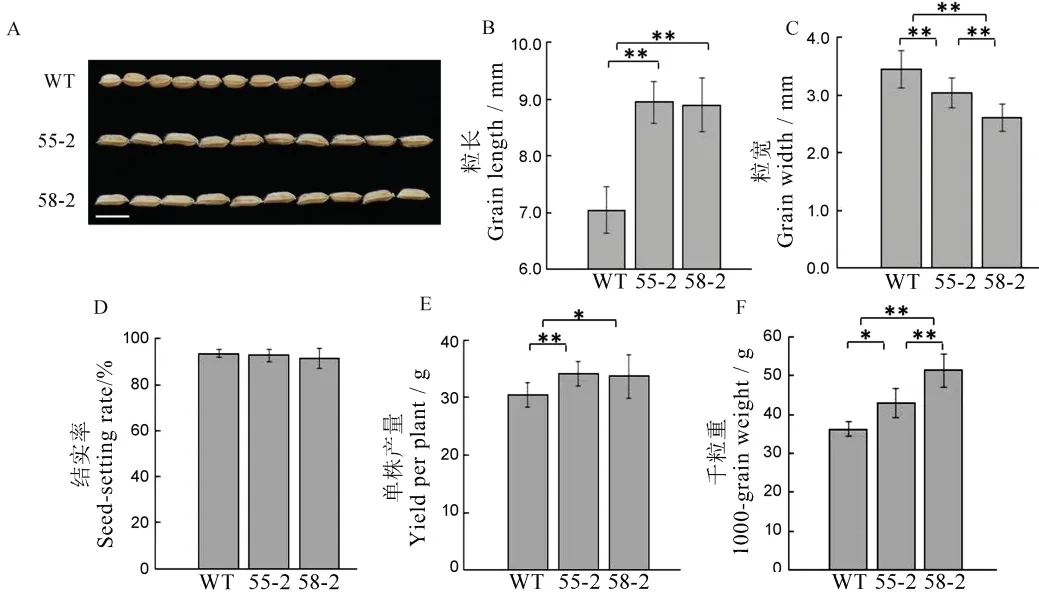
图5 T2 代植株农艺性状考查Fig.5.Agronomic traits of T2 generation plants.
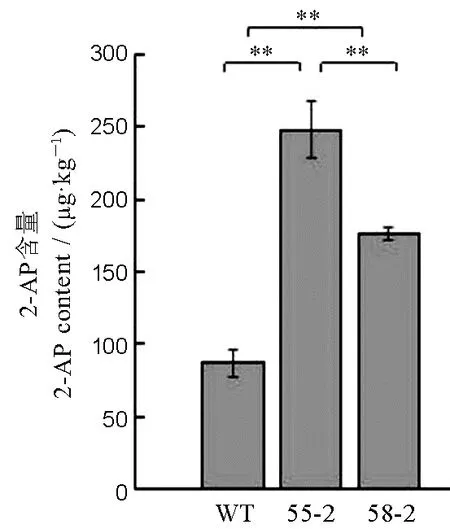
图6 龙粳11 及突变体中香味物质2-AP 的含量Fig.6.2-acetyl-1-pyrroline(2-AP) content of Longjing 11 and mutant plants.
将野生型龙粳11 与55-2 株系、58-2 株系的农艺性状进行比较(图5),发现55-2 株系和58-2 株系的粒长比野生型植株分别增加27.01%、26.43%,都达到了极显著的水平,粒宽较野生型植株降低13.4%、24.6%,结实率没有显著变化,单株产量分别增加10.82%、12.11%,千粒重分别增加18.34%、41.36%,58-2 株系千粒重增加比例显著(图5)。研究表明,通过CRISPR/Cas9 技术同时对GS3、GS9基因进行编辑,高效地将圆粒水稻变长,在育种方面加速了新品系的创制。
采用气相色谱-质谱联用技术,对野生型龙粳11、突变体55-2、58-2 株系中籽粒的2-AP 含量进行分析(图6)。野生型龙粳11 中2-AP 含量约87.13 μg/kg,与龙粳11 对照相比,55-2 株系中香味物质2-AP 含量增加至248.22 μg/kg,58-2 株系中2-AP增加至176.43 μg/kg。55-2 株系和58-2 株系分别在Badh2基因第2 外显子处缺失2 bp、插入1 bp,均产生移码突变。结果表明,通过对Badh2基因进行编辑,能有效创制香型水稻种质新资源。
3 讨论
CRISPR/Cas9 技术与传统育种相比,因其具有操作简单、编辑效率高和成本低等优点,正广泛应用于水稻的种质改良中[32-35],极大地缩短了培育高产优质水稻品种的周期。本研究以粒型负调控基因GS3、GS9和香味负调控基因Badh2为目标基因,构建 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 载体,对GS3、GS9、Badh2基因进行敲除,筛选出两个无T-DNA 元件的三基因纯合突变株系。
GS3和GS9是粒长的负调控因子。Nan 等[36]利用具有gs3等位基因的供体GKBR 与黑龙江省优良栽培品种空育131 杂交后,以空育131 为轮回母本,经过回交3 代改善了空育131 的GS3位点,籽长增加12.05%,千粒重和单株总粒数提高,进而产量增加,但改良周期过长,效率低。通过CRISPR/Cas9 技术可以极大地缩短育种年限,还大幅度提高粒长。Li 等[23]利用CRISPR/Cas9 技术对GS3基因进行编辑,破坏GS3正常表达,粒长增加20%。Zhao 等[14]使用CRISPR/Cas9 技术,在GS9第1 外显子处插入1 bp,使粒长增加5.5%。本研究通过将两个基因连入同一载体,同时对GS3和GS9进行编辑。通过对GS3和GS9基因进行编辑,粒长与野生型相比差异达到极显著水平,增幅为26.43%~27.01%,增幅大于通过CRISPR/Cas9 技术分别敲除GS3和GS9的效果,单株产量增加10.82%~12.11%,结实率没有显著变化,千粒重增加 18.34%~41.36%。研究结果表明,利用CRISPR/Cas9 技术可以培育出长粒型且高产的品种,加快长粒种质资源的创制。
传统的香稻育种主要通过杂交和回交选育,但因其后代分离不稳定、试验周期漫长、抗逆性较弱、产量较低等因素,使得培育出稳定遗传的优良香稻品种较为困难。本研究对Badh2基因进行编辑后,通过对籽粒的2-AP 含量进行分析,Badh2基因功能缺失后香味物质2-AP 含量由87.13 μg/kg 增加至176.43~248.22 μg/kg,香味物质2-AP 显著提高。孙慧宇等[27]通过CRISPR/Cas9 技术对Badh2基因进行编辑,在香味检测时采用咀嚼法和氢氧化钾浸泡法,主观干扰性大。本研究采用气相色谱-质谱联用技术,定量分析水稻籽粒中的2-AP 含量,结果更客观准确。
综上所述,本研究以龙粳11 为材料,通过CRISPR/Cas9 技术对GS3、GS9和Badh2基因进行定点敲除并获得了无T-DNA 元件的三基因纯合突变体材料,使龙粳11 粒型变长,单株产量增加,千粒重增加,同时香味物质2-AP 含量增加,创制出长粒香型水稻品种,为培育产量与品质均提高的水稻品种提供了理论和材料基础,加速了高产优质香稻品种的选育进程。
谢辞:中国科学院东北地理与农业生态研究所对本研究给予了极大帮助,在此表示感谢。
